
The Impact of Carbohydrate Management on Coleoptile Elongation in Anaerobically Germinating Seeds of Rice (Oryza sativa L.) under Light and Dark Cycles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
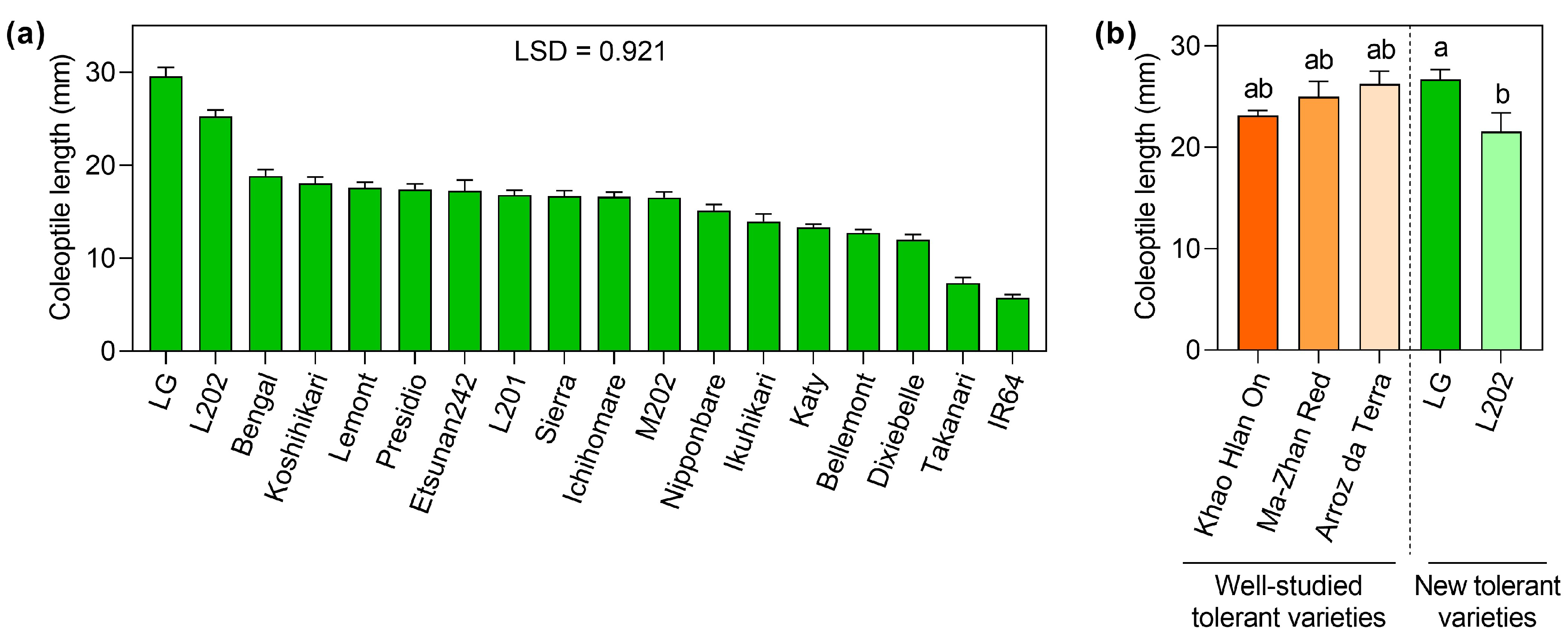
2.1. Screening of Rice Varieties Regarding Coleoptile Elongation under Oxygen Deprivation
2.2. Effect of Light on Coleoptile Elongation under Oxygen Deprivation
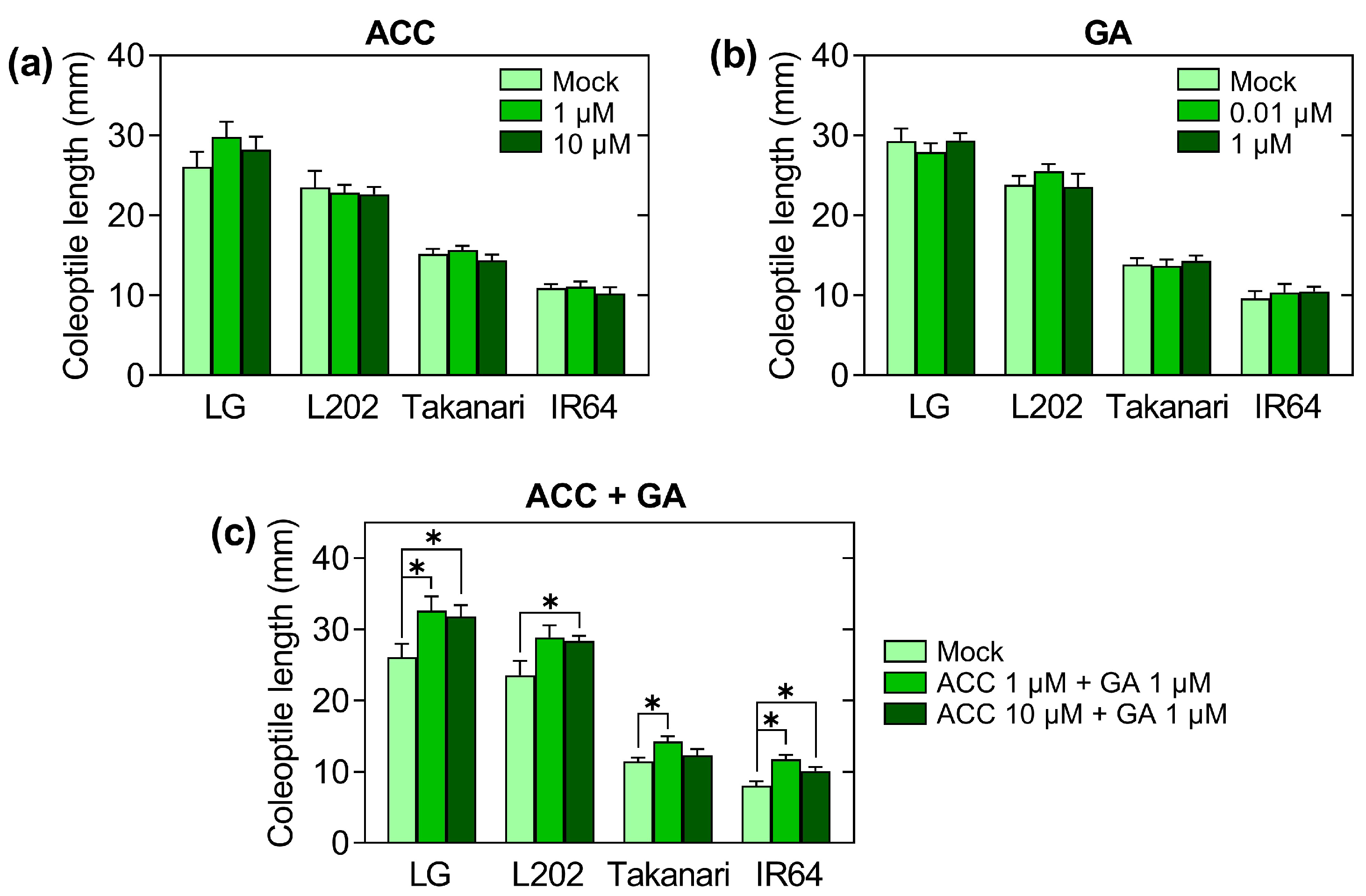
2.3. Effect of Growth-Promoting Hormones on Coleoptile Elongation under Oxygen Deprivation
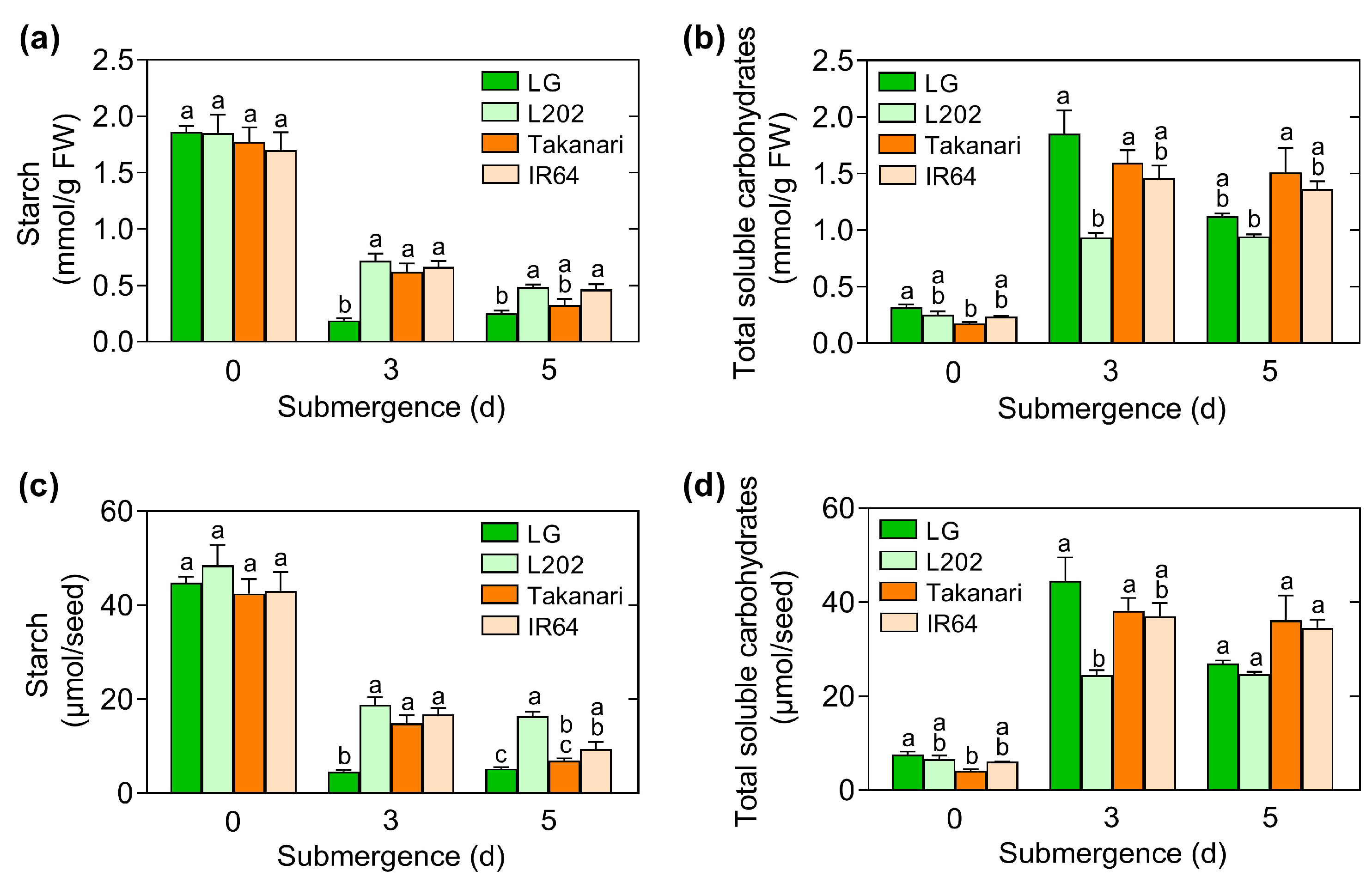
2.4. Role of Starch Breakdown and Soluble Carbohydrate Accumulation in Coleoptile Elongation under Oxygen Deprivation
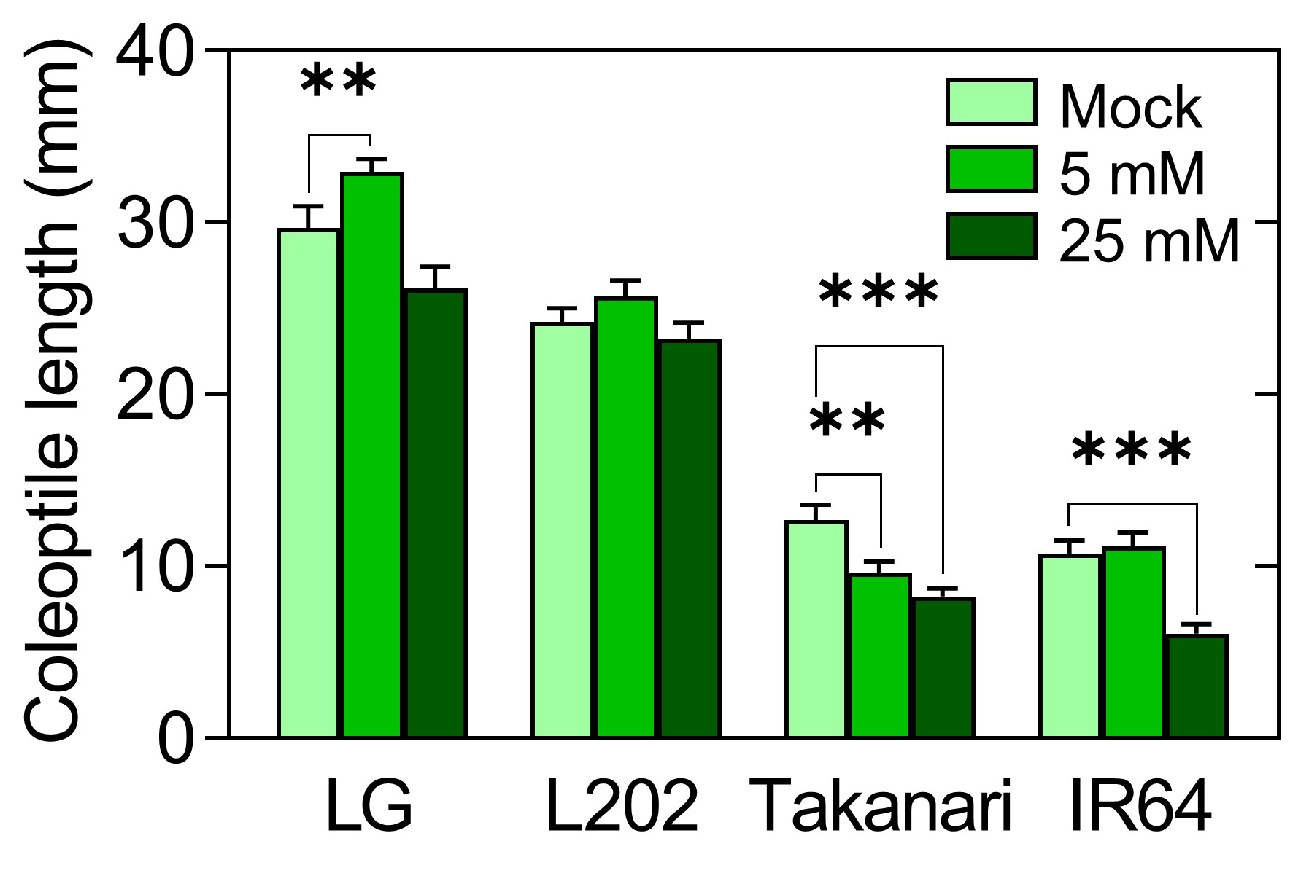
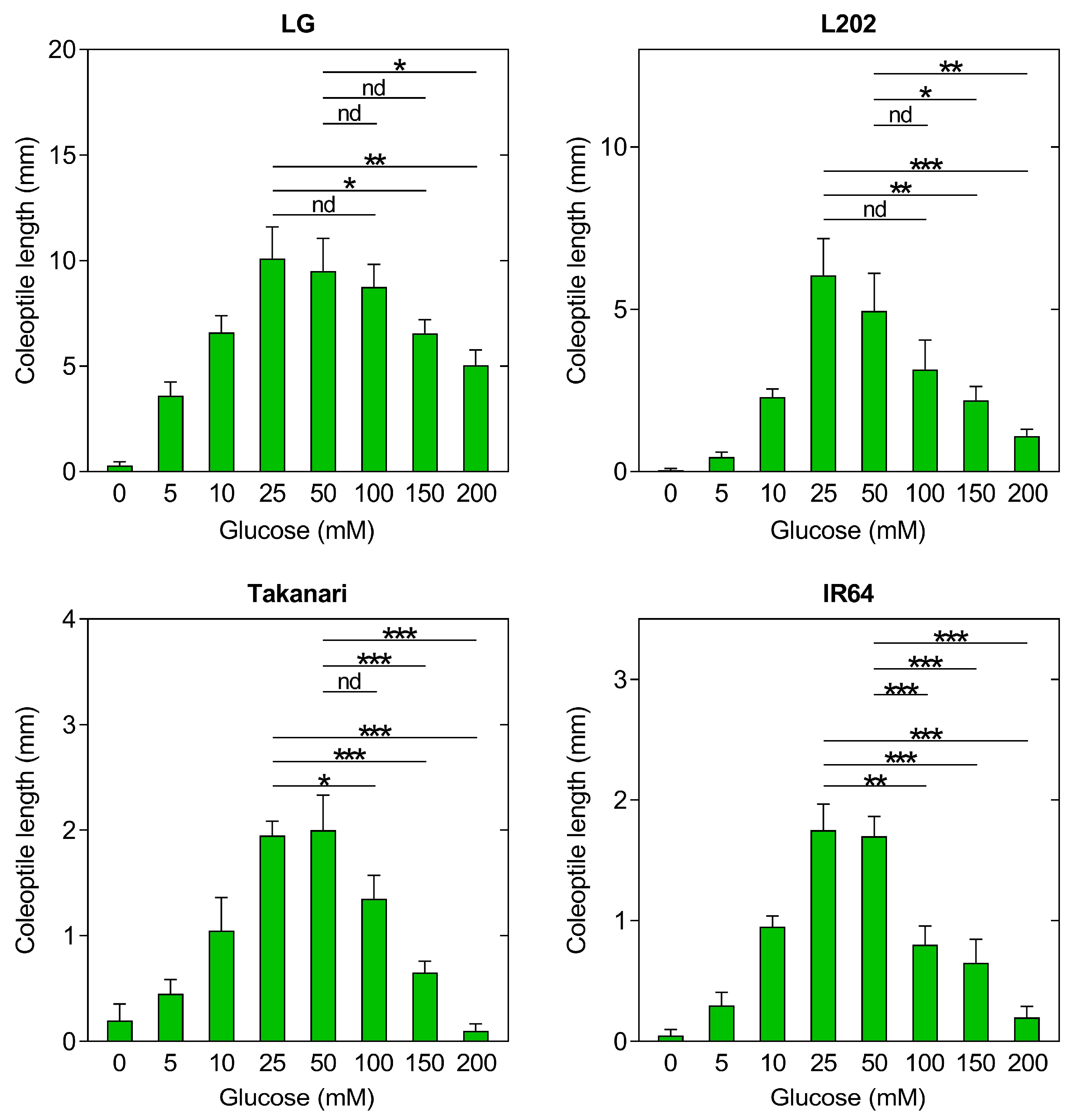
2.5. Effect of Exogenous Glucose on Coleoptile Elongation under Oxygen Deprivation
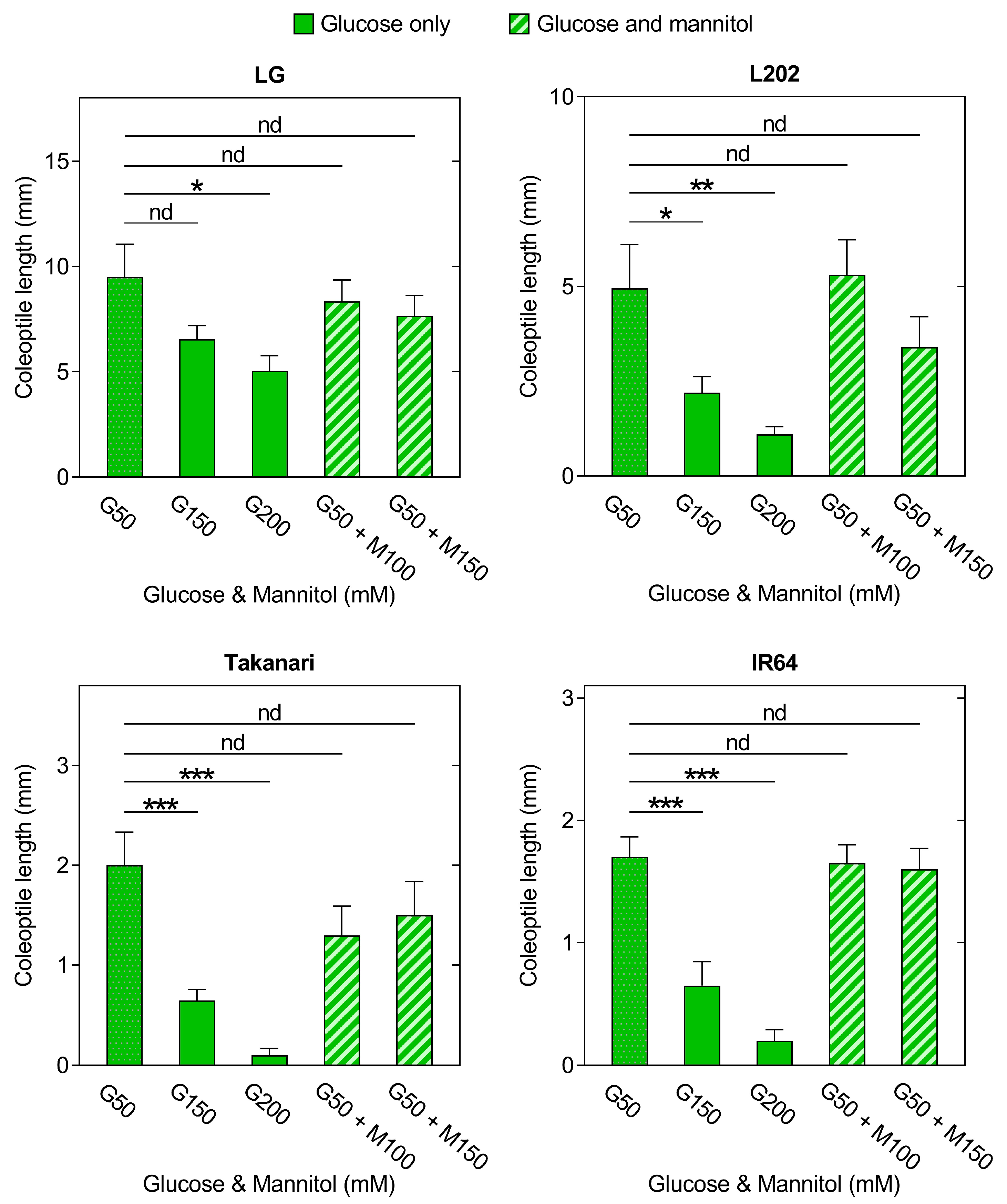
2.6. Impact of Glucose Sensitivity on Coleoptile Elongation under Oxygen Deprivation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Growth Conditions
5.2. Hormone and Carbohydrate Treatments
5.3. Carbohydrate Assays
5.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiono, K.; Koshide, A.; Iwasaki, K.; Oguri, K.; Fukao, T.; Larsen, M.; Glud, R.N. Imaging the Snorkel Effect during Submerged Germination in Rice: Oxygen Supply via the Coleoptile Triggers Seminal Root Emergence Underwater. Front. Plant Sci. 2022, 13, 946776. [Google Scholar] [CrossRef] [PubMed]
- Kordan, H.A. Anaerobiosis-induced etiolation in light-germinated rice seedlings. Ann. Bot. 1976, 40, 347–350. [Google Scholar] [CrossRef]
- Kordan, H.A.; Ashraf, M. Environmental Anoxia is Unnecessary for Inhibiting Chloroplast Photomorphogenesis in Rice Coleoptiles (Oryza sativa L.). J. Exp. Bot. 1990, 41, 435–440. [Google Scholar] [CrossRef]
- Perata, P.; Geshi, N.; Yamaguchi, J.; Akazawa, I. Effect of Anoxia on the Induction of Amylase in Cereal Seeds. Planta 1993, 191, 402–408. [Google Scholar] [CrossRef]
- Guglielminetti, L.; Yamaguchi, J.; Perata, P.; Alpi, A. Amylolytic Activities in Cereal Seeds under Aerobic and Anaerobic Conditions. Plant Physiol. 1995, 109, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms Associated with Tolerance to Flooding during Germination and Early Seedling Growth in Rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Khan, M.I.R.; Entila, F.; Dixit, S.; Pompe, P.C.; Panna Ali, M.; Pittendrigh, B.; Septiningsih, E.M.; Ismail, A.M. Responses of AG1 and AG2 QTL Introgression Lines and Seed Pre-Treatment on Growth and Physiological Processes during Anaerobic Germination of Rice under Flooding. Sci. Rep. 2020, 10, 10214. [Google Scholar] [CrossRef]
- Lu, C.A.; Lim, E.K.; Yu, S.M. Sugar Response Sequence in the Promoter of a Rice α-Amylase Gene Serves as a Transcriptional Enhancer. J. Biol. Chem. 1998, 273, 10120–10131. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.A.; Ho, T.H.D.; Ho, S.L.; Yu, S.M. Three Novel MYB Proteins with One DNA Binding Repeat Mediate Sugar and Hormone Regulation of α-Amylase Gene Expression. Plant Cell 2002, 14, 1963–1980. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.A.; Lin, C.C.; Lee, K.W.; Chen, J.L.; Huang, L.F.; Ho, S.L.; Liu, H.J.; Hsing, Y.I.; Yu, S.M. The SnRK1A Protein Kinase Plays a Key Role in Sugar Signaling during Germination and Seedling Growth of Rice. Plant Cell 2007, 19, 2484–2499. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated Responses to Oxygen and Sugar Deficiency Allow Rice Seedling to Tolerate Flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.R.; Lee, K.W.; Chen, C.Y.; Hong, Y.F.; Chen, J.L.; Lu, C.A.; Chen, K.T.; Ho, T.H.D.; Yu, S.M. SnRK1A-Interacting Negative Regulators Modulate the Nutrient Starvation Signaling Sensor SnRK1 in Source-Sink Communication in Cereal Seedlings under Abiotic Stress. Plant Cell 2014, 26, 808–827. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.S.; Ho, T.H.D.; Liu, L.; Lee, D.H.; Lee, C.H.; Chen, Y.R.; Lin, S.Y.; Lu, C.A.; Yu, S.M. Sugar Starvation-Regulated MYBS2 and 14-3-3 Protein Interactions Enhance Plant Growth, Stress Tolerance, and Grain Weight in Rice. Proc. Natl. Acad. Sci. USA 2019, 116, 21925–21935. [Google Scholar] [CrossRef] [Green Version]
- Angaji, S.A.; Septiningsih, E.M.; Mackill, D.J.; Ismail, A.M. QTLs Associated with Tolerance of Flooding during Germination in Rice (Oryza sativa L.). Euphytica 2010, 172, 159–168. [Google Scholar] [CrossRef]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A Trehalose-6-Phosphate Phosphatase Enhances Anaerobic Germination Tolerance in Rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef]
- Nghi, K.N.; Tondelli, A.; Valè, G.; Tagliani, A.; Marè, C.; Perata, P.; Pucciariello, C. Dissection of Coleoptile Elongation in Japonica Rice under Submergence through Integrated Genome-Wide Association Mapping and Transcriptional Analyses. Plant Cell Environ. 2019, 42, 1832–1846. [Google Scholar] [CrossRef] [PubMed]
- Septiningsih, E.M.; Ignacio, J.C.I.; Sendon, P.M.D.; Sanchez, D.L.; Ismail, A.M.; Mackill, D.J. QTL Mapping and Confirmation for Tolerance of Anaerobic Conditions during Germination Derived from the Rice Landrace Ma-Zhan Red. Theor. Appl. Genet. 2013, 126, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Khan, M.I.R.; Dixit, S.; Cruz, P.C.S.; Ismail, A.M. Growth, productivity and grain quality of AG1 and AG2 QTLs introgression lines under flooding in direct-seeded rice system. Field Crop. Res. 2020, 248, 107713. [Google Scholar] [CrossRef]
- Castano-Duque, L.; Ghosal, S.; Quilloy, F.A.; Mitchell-Olds, T.; Dixit, S. An Epigenetic Pathway in Rice Connects Genetic Variation to Anaerobic Germination and Seedling Establishment. Plant Physiol. 2021, 186, 1042–1059. [Google Scholar] [CrossRef]
- Kumar, V.; Ladha, J.K. Direct seeding of rice: Recent developments and future research needs. Adv. Agron. 2011, 111, 297–413. [Google Scholar]
- Yu, S.M.; Lee, H.T.; Lo, S.F.; Ho, T.H.D. How Does Rice Cope with Too Little Oxygen during Its Early Life? New Phytol. 2021, 229, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Iwata, N.; Shinada, H.; Kiuchi, H.; Sato, T.; Fujino, K. Mapping of QTLs Controlling Seedling Establishment Using a Direct Seeding Method in Rice. Breed. Sci. 2010, 60, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A Variable Cluster of Ethylene Response Factor-like Genes Regulates Metabolic and Developmental Acclimation Responses to Submergence in Rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Fukao, T.; Bailey-Serres, J. Submergence Tolerance Conferred by Sub1A Is Mediated by SLR1 and SLRL1 Restriction of Gibberellin Responses in Rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Lu, X.; Ma, B.; Chen, S.Y.; Zhang, J.S. Ethylene Signaling in Rice and Arabidopsis: Conserved and Diverged Aspects. Mol. Plant 2015, 8, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Sponsel, V.M. Signal achievements in gibberellin research: The second half-century. Annu. Plant Rev. 2016, 49, 1–36. [Google Scholar]
- Hedden, P. The Current Status of Research on Gibberellin Biosynthesis. Plant Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef]
- Watanabe, H.; Hase, S.; Saigusa, M. Effects of the Combined Application of Ethephon and Gibberellin on Growth of Rice (Oryza sativa L.) Seedlings. Plant Prod. Sci. 2007, 10, 468–472. [Google Scholar] [CrossRef] [Green Version]
- Magneschi, L.; Perata, P. Rice Germination and Seedling Growth in the Absence of Oxygen. Ann. Bot. 2009, 103, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Chen, P.W.; Yu, S.M. Metabolic Adaptation to Sugar/O2 Deficiency for Anaerobic Germination and Seedling Growth in Rice. Plant Cell Environ. 2014, 37, 2234–2244. [Google Scholar]
- Perata, P.; Pozueta-Romero, J.; Akazawa, T.; Yamaguchi, J. Effect of Anoxia on Starch Breakdown in Rice and Wheat Seeds. Planta 1992, 188, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Arenas-Huertero, F.; Arroyo, A.; Zhou, L.; Sheen, J.; León, P. Analysis of Arabidopsis glcose insensitive mutants, gin5 and gin6, reveals a central role of the plant hormone ABA in the regulation of plant vegetative development by sugar. Genes Dev. 2000, 14, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.; Zhou, L.; Rolland, F.; Hall, Q.; Cheng, W.-H.; Liu, Y.-X.; Hwang, I.; Jones, T.; Sheen, J. Role of the Arabidopsis Glucose Sensor HXK1 in Nutrient, Light, and Hormonal Signaling. Science 2003, 300, 332–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.I.; Ryoo, N.; Eom, J.S.; Lee, D.W.; Kim, H.B.; Jeong, S.W.; Lee, Y.H.; Kwon, Y.K.; Cho, M.H.; Bhoo, S.H.; et al. Role of the Rice Hexokinases OsHXK5 and OsHXK6 as Glucose Sensors. Plant Physiol. 2009, 149, 745–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, B.G.; Fukao, T. Plant Adaptation to Multiple Stresses during Submergence and Following Desubmergence. Int. J. Mol. Sci. 2015, 16, 30164–30180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, R.F. The effect of ethylene and other regulators on coleoptile growth of rice under anoxia. Plant Sci. 1991, 79, 57–62. [Google Scholar] [CrossRef]
- Loreti, E.; Yamaguchi, J.; Alpi, A.; Perata, P. Gibberellins Are Not Required for Rice Germination under Anoxia. Plant Soil 2003, 253, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Alpuerto, J.; Hussain, R.M.F.; Fukao, T. The key regulator of submergence tolerance, SUB1A, promotes photosynthetic and metabolic recovery from submergence damage in rice leaves. Plant Cell Environ. 2016, 39, 672–684. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirano, H.; Watanabe, T.; Fukuda, M.; Fukao, T. The Impact of Carbohydrate Management on Coleoptile Elongation in Anaerobically Germinating Seeds of Rice (Oryza sativa L.) under Light and Dark Cycles. Plants 2023, 12, 1565. https://doi.org/10.3390/plants12071565
Hirano H, Watanabe T, Fukuda M, Fukao T. The Impact of Carbohydrate Management on Coleoptile Elongation in Anaerobically Germinating Seeds of Rice (Oryza sativa L.) under Light and Dark Cycles. Plants. 2023; 12(7):1565. https://doi.org/10.3390/plants12071565
Chicago/Turabian StyleHirano, Haru, Takeru Watanabe, Mika Fukuda, and Takeshi Fukao. 2023. "The Impact of Carbohydrate Management on Coleoptile Elongation in Anaerobically Germinating Seeds of Rice (Oryza sativa L.) under Light and Dark Cycles" Plants 12, no. 7: 1565. https://doi.org/10.3390/plants12071565