
Differences in Physiological Characteristics, Seed Germination, and Seedling Establishment in Response to Salt Stress between Dimorphic Seeds in the Halophyte Suaeda liaotungensis
Abstract
:1. Introduction
2. Results
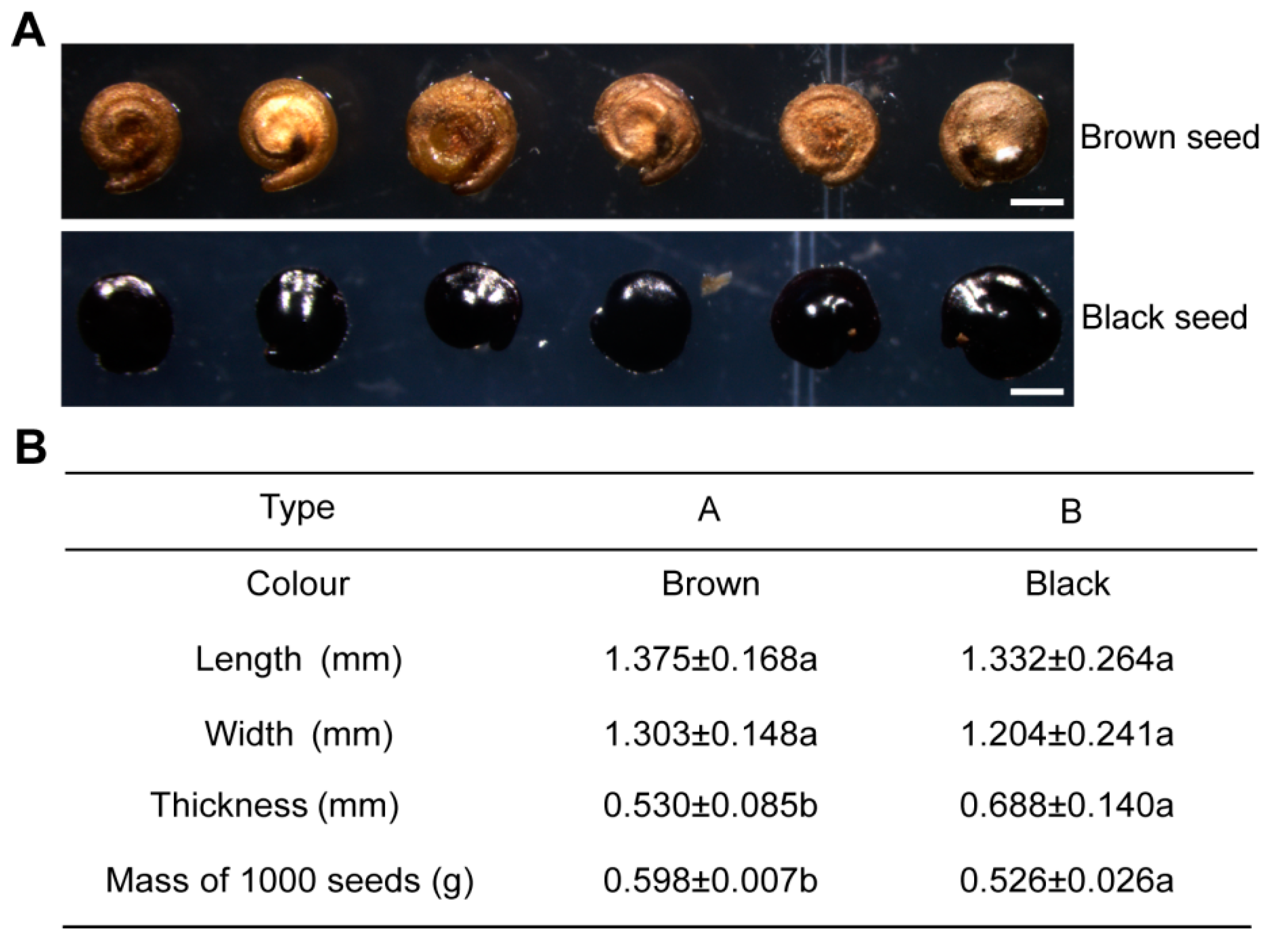
2.1. Morphological Characteristics of Dimorphic Seeds in S. liaotungensis
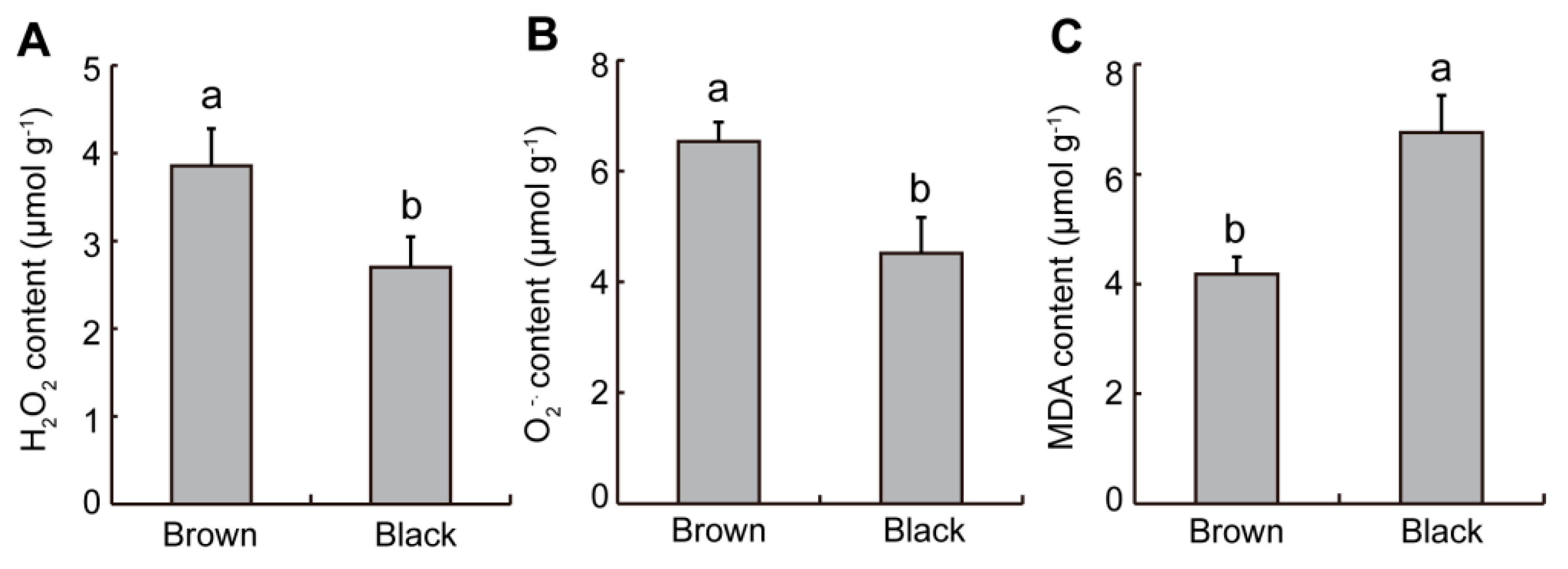
2.2. Difference in ROS Levels from Dimorphic Seeds in S. liaotungensis
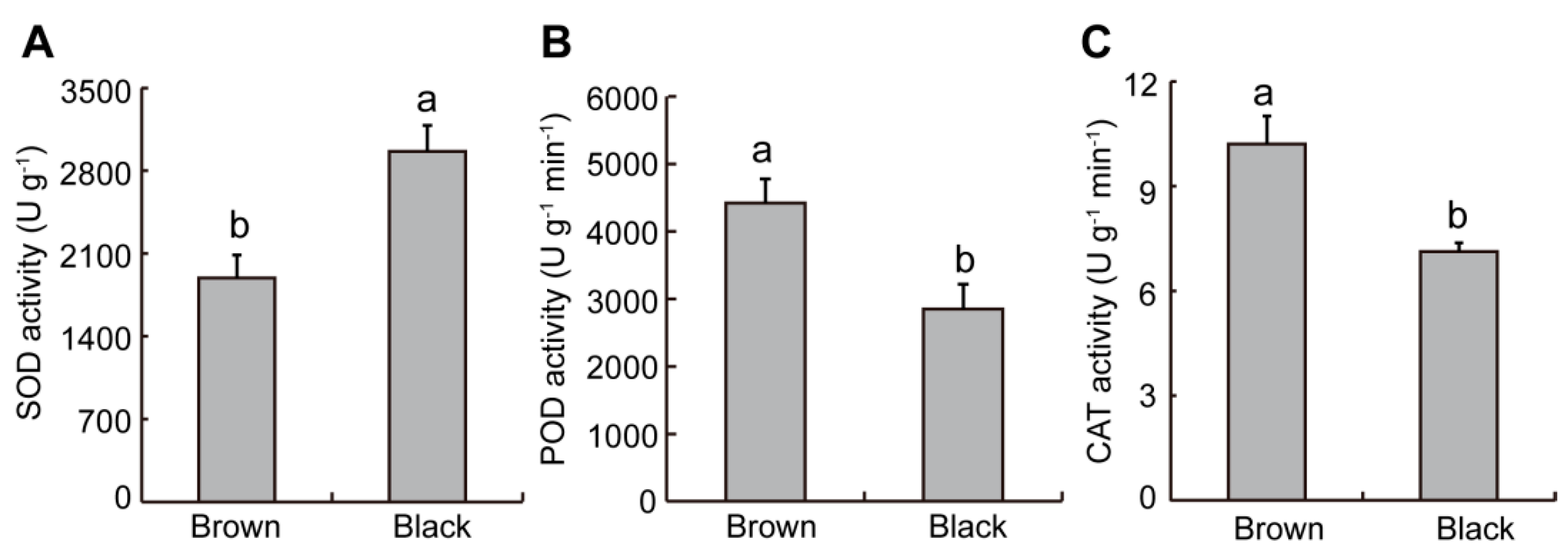
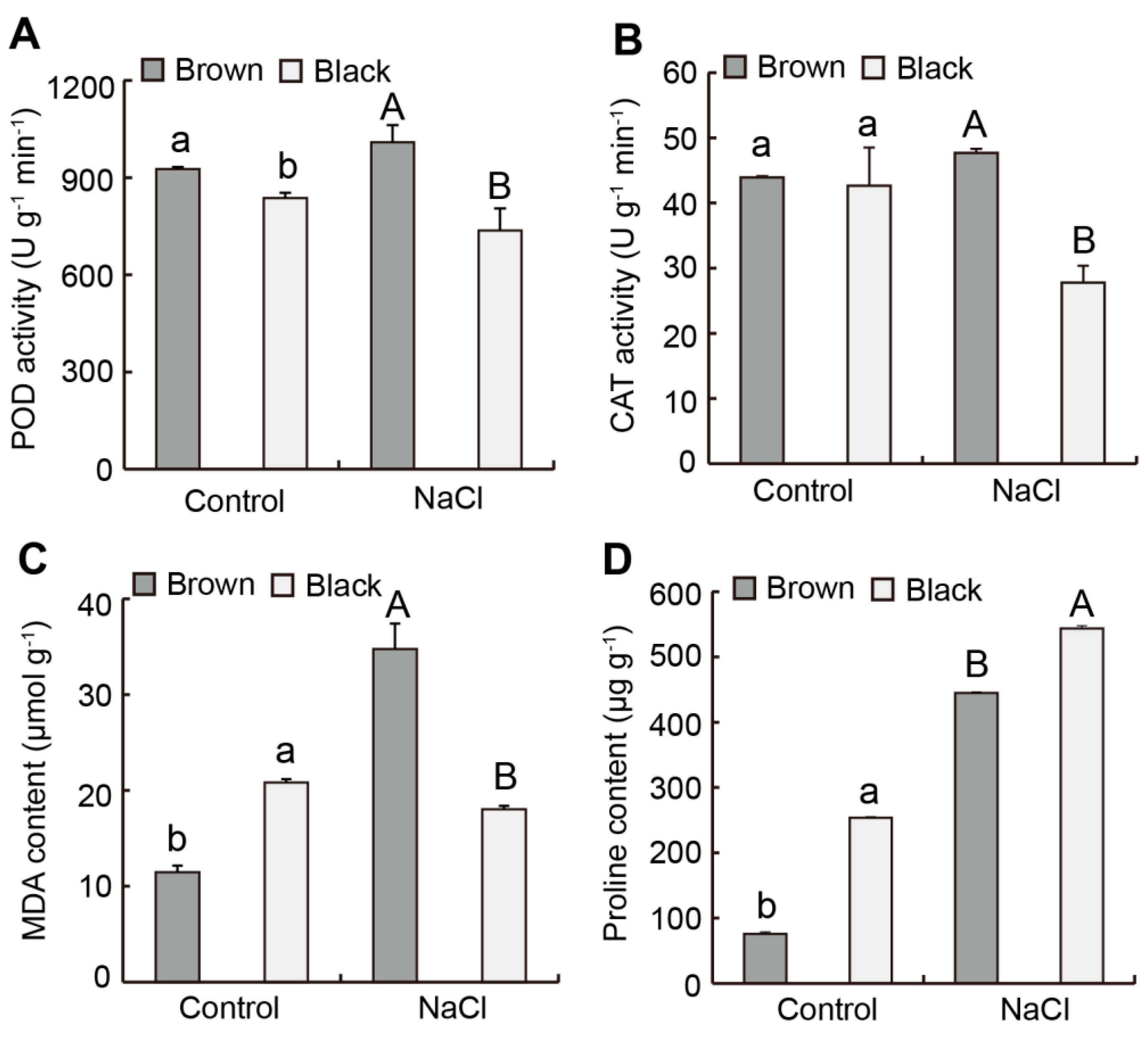
2.3. Difference in Antioxidant Enzyme Activity from Dimorphic Seeds in S. liaotungensis
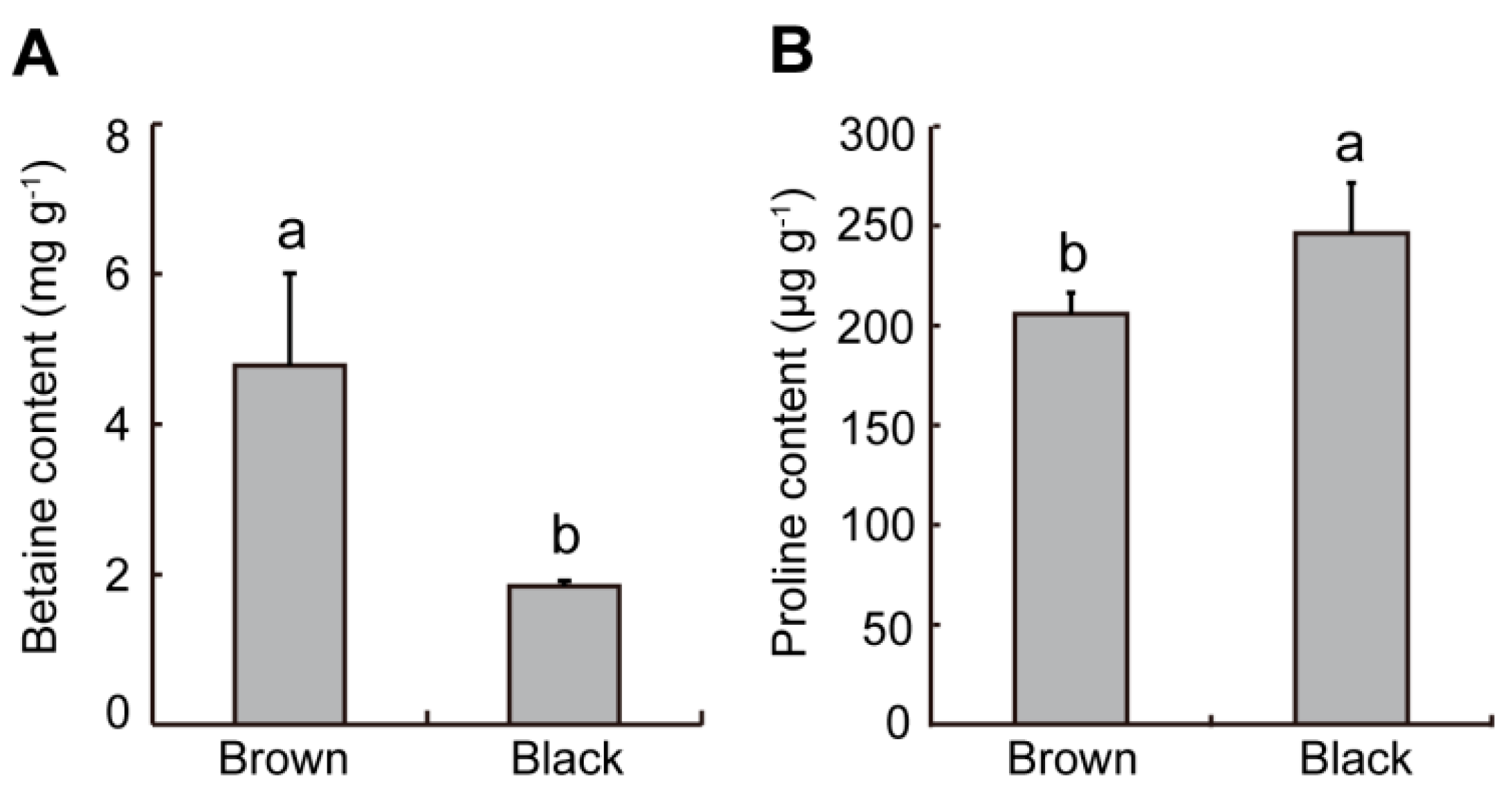
2.4. Difference in Osmotic Regulating Substances from Dimorphic Seeds in S. liaotungensis
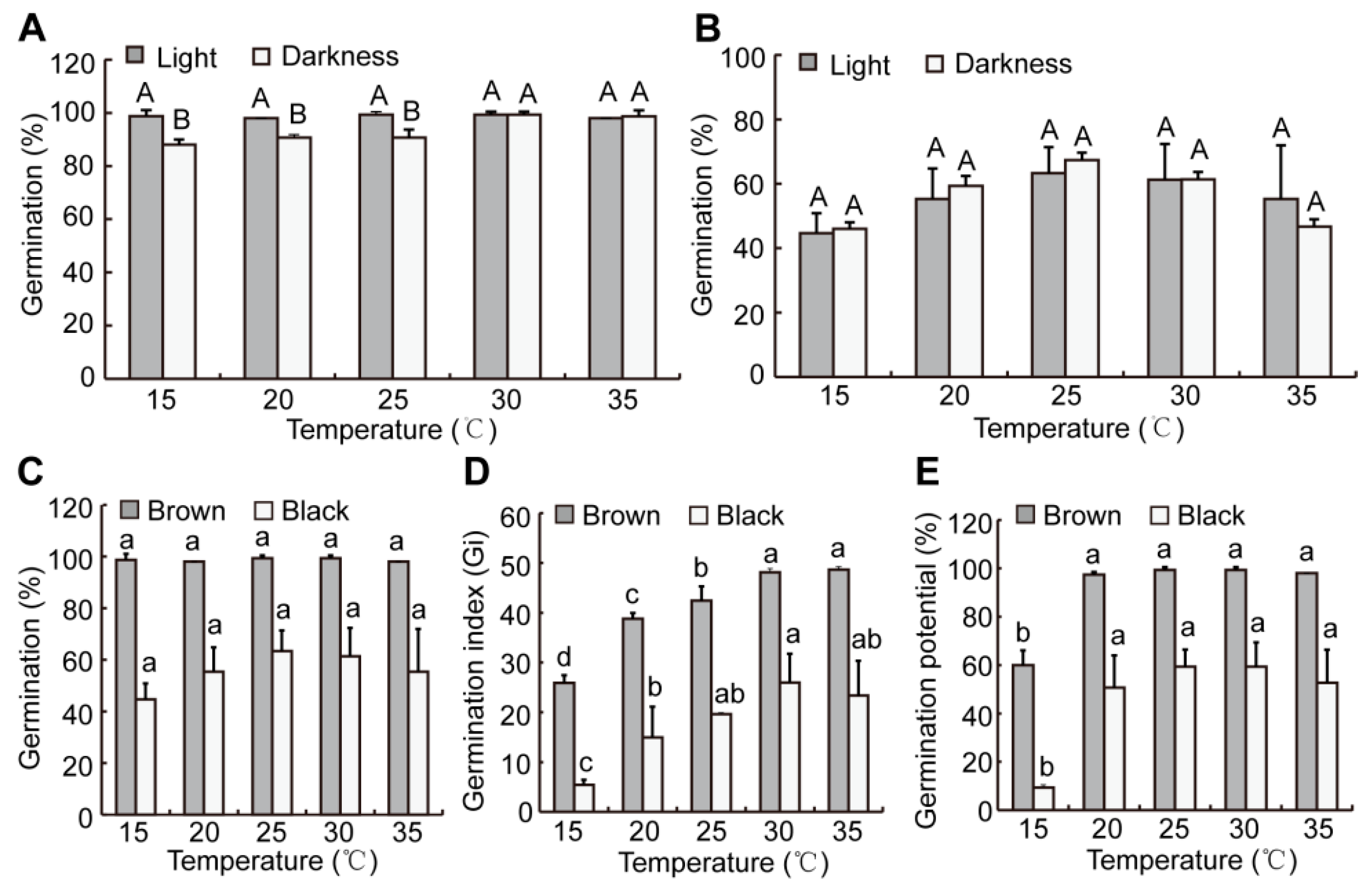
2.5. Effects of Light and Temperature on Seed Germination in S. liaotungensis
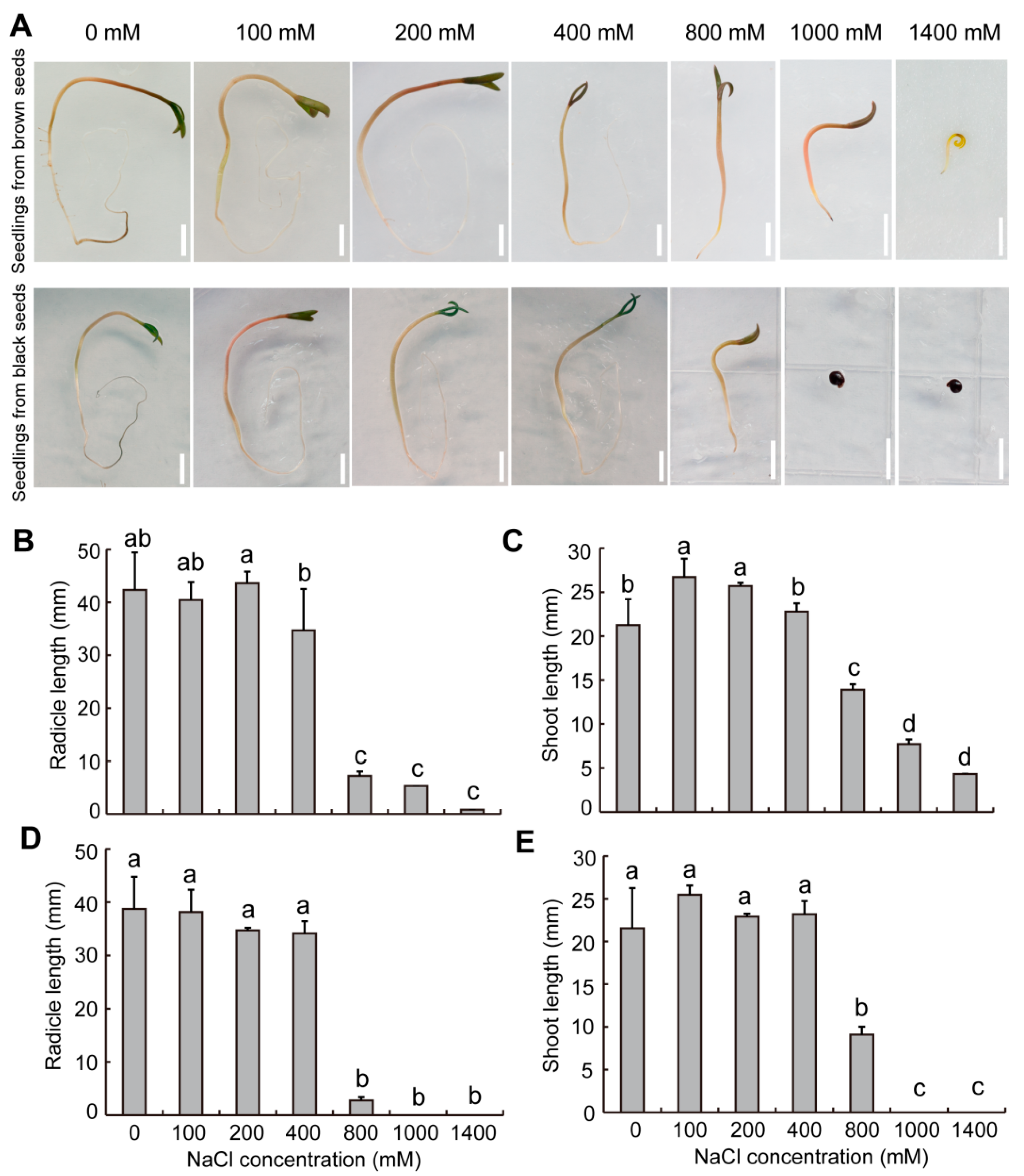
2.6. Effect of Salinity on Seed Germination in S. liaotungensis
2.7. Effect of Salinity on the Physiological Indexes of Dimorphic Seeds during Germination in S. liaotungensis
2.8. Effect of Salinity on Seedling Growth in S. liaotungensis
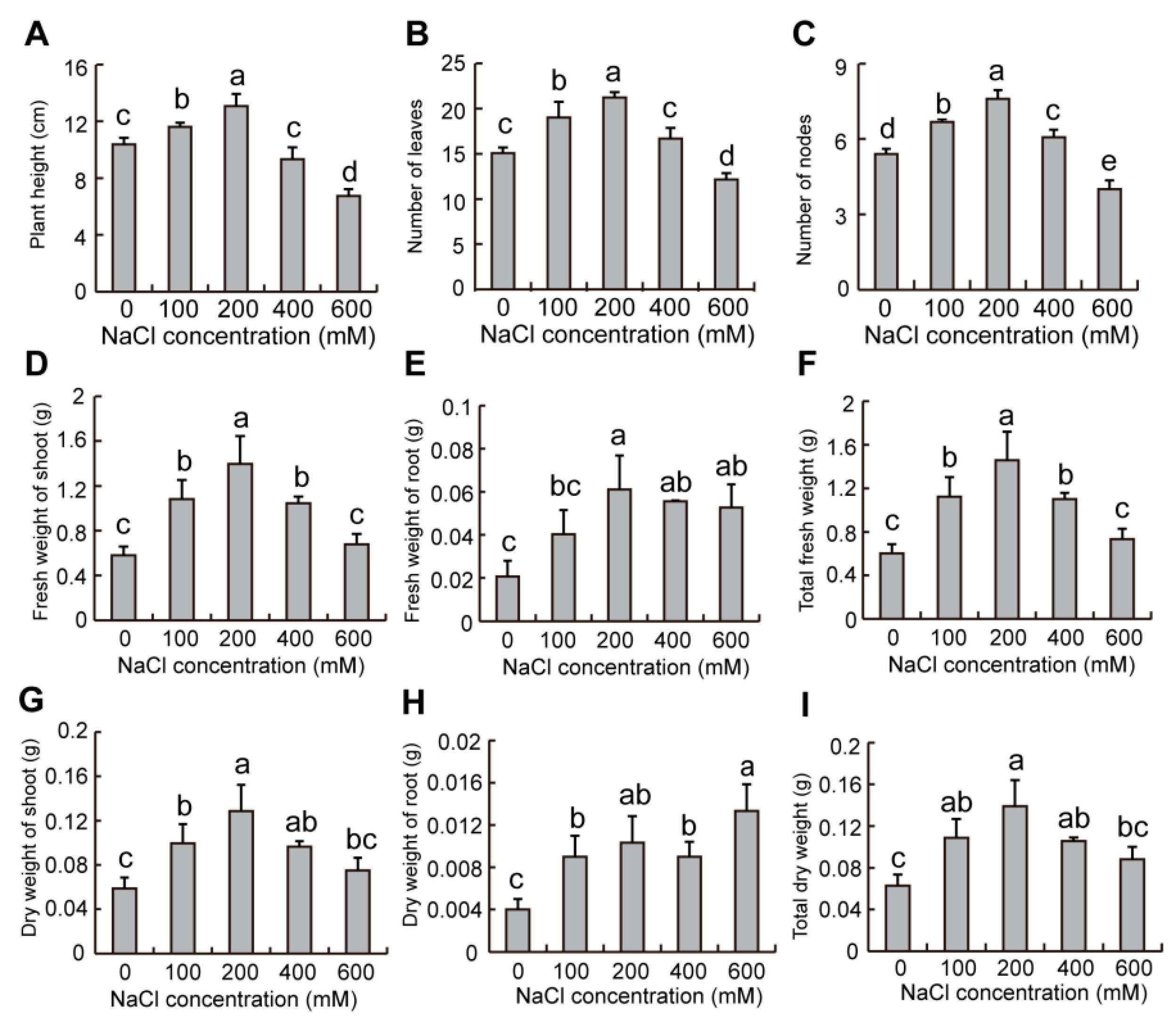
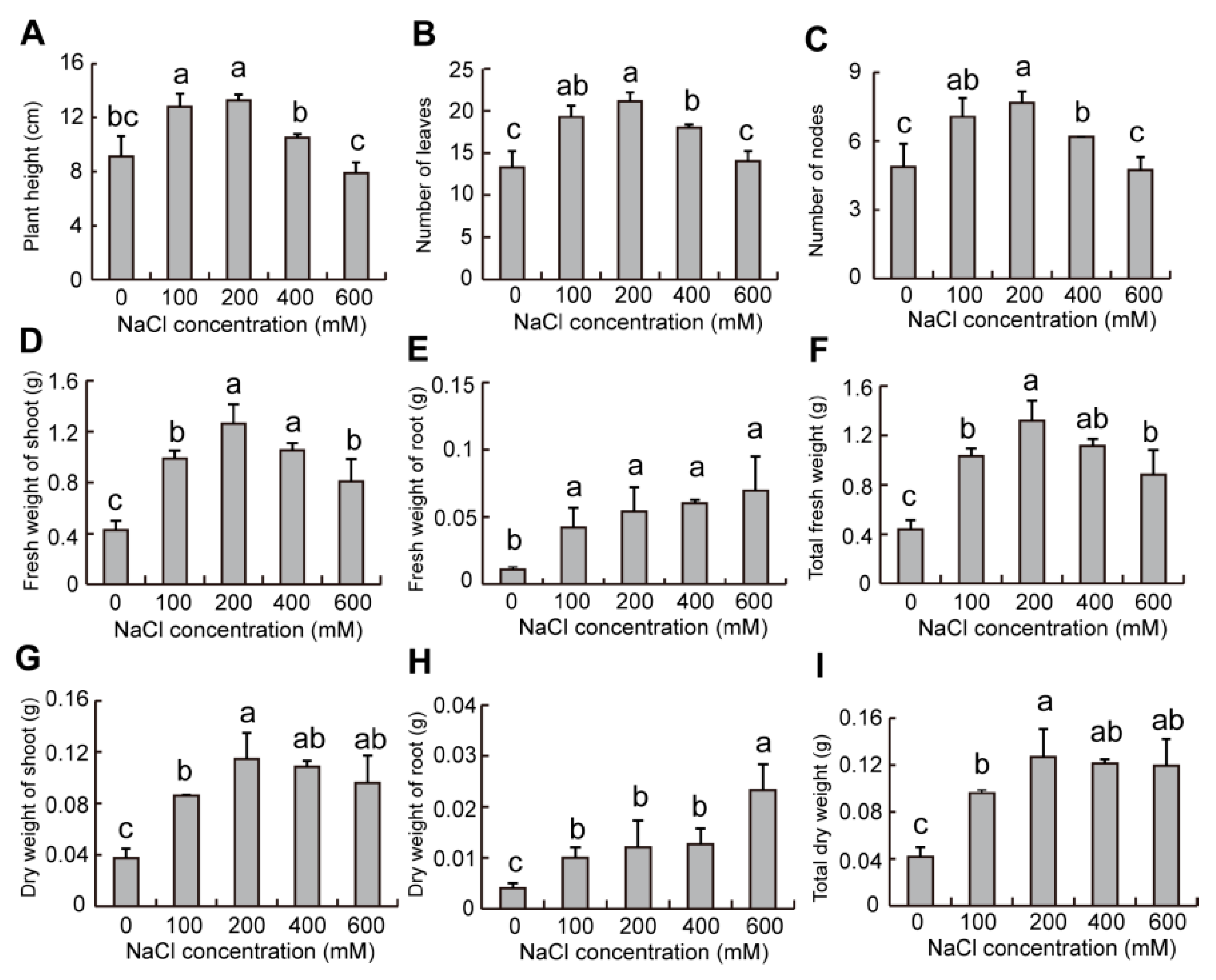
2.9. Effect of Salinity on Growth of Plants in S. liaotungensis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Morphological Characteristics of Dimorphic Seeds
4.3. Determination of Physiological Indexes
4.4. Seed Germination
4.5. Seedling Growth
4.6. Plant Growth
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Zelm, E.; Zhang, Y.X.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Panuccio, M.R.; Jacobsen, S.E.; Akhtar, S.S.; Muscolo, A. Effect of saline water on seed germination and early seedling growth of the halophyte quinoa. AoB Plants 2014, 6, plu047. [Google Scholar] [CrossRef]
- Hariadi, Y.; Marandon, K.; Tian, Y.; Jacobsen, S.E.; Shabala, S. Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 2011, 62, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungar, I.A. Population ecology of halophyte seeds. Bot. Rev. 1987, 53, 301–334. [Google Scholar] [CrossRef]
- Ungar, I.A. Salinity, temperature, and growth regulator effects on seed germination of Salicornia europaea L. Aquat. Bot. 1977, 3, 329–335. [Google Scholar] [CrossRef]
- Galatino, M.I.; Van der Valk, A.G. Seed germination traits of annuals and emergents recruited during drawdowns in the Delta marsh, Manitoba, Canada. Aquat. Bot. 1986, 26, 89–102. [Google Scholar]
- Mohammad, S.; Sen, D.N. A report on polymorphic seeds in halophytes Trianthema triquetra L. in Indian desert. Curr. Sci. 1988, 57, 616–617. [Google Scholar]
- Sorensen, A.E. Somatic polymorphism and seed dispersal. Nature 1978, 276, 174–176. [Google Scholar] [CrossRef]
- Khan, M.A.; Gul, B.; Weber, D.J. Germination of dimorphic seeds of Suaeda moquinii under high salinity stress. Aust. J. Bot. 2001, 49, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Huang, Z.Y.; Baskin, C.C.; Baskin, J.M.; Dong, M. Germination of dimorphic seeds of the desert annual halophyte Suaeda aralocaspica (Chenopodiaceae), a C4 plant without Kranz anatomy. Ann. Bot. 2008, 102, 757–769. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar]
- Wang, H.L.; Tian, C.Y.; Wang, L. Germination of dimorphic seeds of Suaeda aralocaspica in response to light and salinity conditions during and after cold stratification. PeerJ 2017, 5, e3671. [Google Scholar] [CrossRef] [Green Version]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 2014; pp. 150–162. [Google Scholar]
- Zhang, H.; Hu, M.F.; Ma, H.Y.; Jiang, L.; Zhao, Z.Y.; Ma, J.B.; Wang, L. Differential responses of dimorphic seeds and seedlings to abiotic stresses in the halophyte Suaeda salsa. Front. Plant Sci. 2021, 12, 630338. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Shi, W.W.; Liu, R.R.; Xu, Y.G.; Sui, N.; Zhou, J.C.; Feng, G. The role of the seed coat in adaptation of dimorphic seeds of the euhalophyte Suaeda salsa to salinity. Plant Spec. Biol. 2017, 32, 107–114. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Zhang, H.; Tian, Y.; Guan, B.; Zhou, D.; Sun, Z.; Baskin, C.C. The best salt solution parameter to describe seed/seedling responses to saline and sodic salts. Plant Soil 2018, 426, 313–325. [Google Scholar] [CrossRef]
- Weber, D.J. Adaptive mechanisms of halophytes in desert regions. In Salinity and Water Stress; Springer: Dordrecht, The Netherlands, 2009; Volume 44, pp. 179–185. [Google Scholar]
- El-Keblawy, A.; Al-Rawai, A. Effects of salinity, temperature and light on germination of invasive Prosopis juliflora (Sw.) DC. J. Arid. Environ. 2005, 61, 555–565. [Google Scholar] [CrossRef]
- Bhatt, A.; Bhat, N.R.; Suleiman, M.K.; Santo, A. Effects of storage, mucilage presence, photoperiod, thermoperiod and salinity on germination of Farsetia aegyptia Turra (Brassicaceae) seeds: Implications for restoration and seed banks in Arabian Desert. Plant Biosyst. 2019, 153, 280–287. [Google Scholar] [CrossRef]
- Shang, C.; Wang, L.; Tian, C.Y.; Song, J. Heavy metal tolerance and potential for remediation of heavy metal-contaminated saline soils for the euhalophyte Suaeda salsa. Plant Signal Behav. 2020, 15, 1805902. [Google Scholar] [CrossRef]
- Wu, D.D.; Sun, Y.H.; Wang, H.F.; Shi, H.; Su, M.X.; Shan, H.Y.; Li, T.T.; Li, Q.L. The SlNAC8 gene of the halophyte Suaeda liaotungensis enhances drought and salt stress tolerance in transgenic Arabidopsis thaliana. Gene 2018, 662, 10–20. [Google Scholar] [CrossRef]
- Li, Q.L.; Lai, L.D.; Yu, X.D.; Wang, J.Y. The SlCMO gene driven by its own promoter enhances salt tolerance of transgenic tomato without affecting growth and yield. Plant Mol. Biol. Rep. 2018, 36, 596–604. [Google Scholar] [CrossRef]
- Du, X.R.; Su, M.X.; Jiao, Y.; Xu, S.X.; Song, J.Q.; Wang, H.F.; Li, Q.L. A transcription factor SlNAC10 gene of Suaeda liaotungensis regulates proline synthesis and enhances salt and drought tolerance. Int. J. Mol. Sci. 2022, 23, 9625. [Google Scholar] [CrossRef]
- Shao, L.; Jiang, H. Effect of season and variation in salinity on rhizosphere fungal diversity of Suaeda liaotungensis. Acta Ecol. Sin. 2016, 36, 1050–1057. [Google Scholar]
- Li, J.G.; Wang, Y.; Pritchard, H.W.; Wang, X.F. The fluxes of H2O2 and O2 can be used to evaluate seed germination and vigor of Caragana korshinskii. Planta 2014, 239, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Kamal, M.M.; Erazo, C.; Tanino, K.K.; Kawamura, Y.; Kasuga, J.; Laarveld, B.; Olkowski, A.; Uemura, M. A single seed treatment mediated through reactive oxygen species increases germination, growth performance, and abiotic stress tolerance in Arabidopsis and rice. Biosci. Biotechnol. Biochem. 2020, 84, 2597–2608. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandák, B.; Pyšek, P. The effects of light quality, nitrate concentration and presence of bracteoles on germination of different fruit types in the heterocarpous Atriplex sagittata. J. Ecol. 2001, 89, 149–158. [Google Scholar] [CrossRef]
- Mandák, B.; Pyšek, P. Fruit dispersal and seed banks in Atriplex sagittata: The role of heterocarpy. J. Ecol. 2001, 89, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, A.; Murata, N. Genetic engineering of glycinebetaine synthesis in plants: Current status and implications for enhancement of stress tolerance. J. Exp. Bot. 2000, 51, 81–88. [Google Scholar] [CrossRef]
- Cao, J.; Lv, X.Y.; Chen, L.; Xing, J.J.; Lan, H.Y. Effects of salinity on the growth, physiology and relevant gene expression of an annual halophyte grown from heteromorphic seeds. AoB Plants 2015, 7, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisar, F.; Gul, B.; Khan, M.A.; Hameed, A. Heteromorphic seeds of coastal halophytes Arthrocnemum macrostachyum and A. indicum display differential patterns of hydrogen peroxide accumulation, lipid peroxidation and antioxidant activities under increasing salinity. Plant Physiol. Biochem. 2019, 144, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. Comptes Rendus Biol. 2008, 31, 806–814. [Google Scholar] [CrossRef]
- Bailly, C. The signalling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Müller, K.; Linkies, A.; Vreeburg, R.A.M.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress seed germination and elongation growth. Plant Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [Green Version]
- Li, X.R.; Yao, S.X.; Chen, S.S.; Lan, H.Y. Physiological responses to salt stress of plants derived from heteromorphic seeds of Chenopodium album. Acta Ecol. Sin. 2015, 35, 8139–8147. [Google Scholar]
- Banu, N.A.; Hoque, A.; Watanabe-Sugimoto, M.; Matsuoka, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Proline and glycinebetaine induce antioxidant defense gene expression and suppress cell death in cultured tobacco cells under salt stress. J. Plant Physiol. 2009, 166, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Li, W.Q.; Yamaguchi, S.; Khan, M.A.; An, P.; Liu, X.J.; Tran, L.P. Roles of gibberellins and abscisic acid in regulating germination of Suaeda salsa dimorphic seeds under salt stress. Front. Plant Sci. 2016, 6, 1235. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Baskin, C.C.; Baskin, J.M.; Yang, F.; Huang, Z. Comparison of germination and seed bank dynamics of dimorphic seeds of the cold desert halophyte Suaeda corniculata subsp. mongolica. Ann. Bot. 2012, 110, 1545–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Gui, B.; Weber, D.J. Temperature and high salinity effects in germinating dimorphic seeds of Atriplex rose. West. N. Am. Nat. 2004, 64, 193–201. [Google Scholar]
- Lee, M.H.; Cho, E.J.; Wi, S.G.; Bae, H.; Kim, J.E.; Cho, J.Y.; Lee, S.; Kim, J.H.; Chung, B.Y. Divergences in morphological changes and antioxidant responses in salt-tolerant and salt-sensitive rice seedlings after salt stress. Plant Physiol. Biochem. 2013, 70, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Aycan, M.; Baslam, M.; Asiloglu, R.; Mitsui, T.; Yildiz, M. Development of new high-salt tolerant bread wheat (Triticum aestivum L.) genotypes and insight into the tolerance mechanisms. Plant Physiol. Biochem. 2021, 166, 314–327. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (Oryza sativa L.) varieties. Biomed Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [Green Version]
- Mo, X.; Qian, J.Y.; Liu, P.; Zeng, H.L.; Chen, G.H.; Wang, Y. Exogenous betaine enhances the protrusion vigor of rice seeds under heat stress by regulating plant hormone signal transduction and its interaction network. Antioxidants 2022, 11, 1792. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.C.; Fu, T.T.; Sui, N.; Guo, J.R.; Feng, G.; Fan, J.L.; Song, J. The role of salinity in seed maturation of the euhalophyte Suaeda salsa. Plant Biosyst. 2016, 150, 83–90. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Cambrollé, J.; Luque, T.; Figueroa, M.E.; Davy, A.J. Carry-over of differential salt tolerance in plants grown from dimorphic seeds of Suaeda splendens. Ann. Bot. 2008, 102, 103–112. [Google Scholar] [CrossRef]
- Guo, J.R.; Du, M.; Tian, H.Y.; Wang, B.S. Exposure to high salinity during seed development markedly enhances seedling emergence and fitness of the progeny of the extreme halophyte Suaeda salsa. Front. Plant Sci. 2020, 11, 1291. [Google Scholar] [CrossRef]
- Brennan, T.; Frenkel, C. Involvement of hydrogen peroxide in the regulation of senescence in pear. Plant Physiol. 1977, 59, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K. Chromium-mediated oxidative stress and ultrastructural changes in root cells of developing rice seedlings. J. Plant Physiol. 2007, 164, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Q.; Liu, X.J.; Li, X.X.; Wang, H.F.; Chu, R.W.; Qu, F.F.; Zhang, S.X.; Li, Q.L. Transcriptome analysis reveals genes and pathways associated with salt tolerance during seed germination in Suaeda liaotungensis. Int. J. Mol. Sci. 2022, 23, 12229. [Google Scholar] [CrossRef] [PubMed]
- Song, S.Y.; Chen, Y.; Chen, J.; Dai, X.Y.; Zhang, W.H. Physiological mechanisms underlying OsNAC5-dependent tolerance of rice plants to abiotic stress. Planta 2011, 234, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, L.H.; Zhao, J.F.; Song, Y.; Zhang, C.J.; Guo, Y. Identification of an apoplastic protein involved in the initial phase of salt stress response in rice root by two-dimensional electrophoresis. Plant Physiol. 2009, 149, 916–928. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Pan, K.W.; Wang, J.C.; Ma, Y.H. Effects of phenolic acids on seed germination and seedling antioxidant enzyme activity of Alfalfa. Acta Ecol. Sin. 2006, 26, 3393–3403. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl Concentration (mM) | Brown Seed | Black Seed | ||||
|---|---|---|---|---|---|---|
| Germination Percentage (%) | Germination Recovery Percentage (%) | Final Germination Percentage (%) | Germination Percentage (%) | Germination Recovery Percentage (%) | Final Germination Percentage (%) | |
| 0 | 100.0 ± 0.0 Aa | 0.0 ± 0.0 Ac | 100.0 ± 0.0 Aa | 49.3 ± 4.6 Ba | 0.0 ± 0.0 Ac | 49.3 ± 4.6 Ba |
| 100 | 96.0 ± 6.9 Aa | 0.0 ± 0.0 Ac | 96.0 ± 6.9 Aab | 44.0 ± 2.0 Ba | 0.0 ± 0.0 Ac | 44.0 ± 2.0 Ba |
| 200 | 98.0 ± 0.0 Aa | 0.0 ± 0.0 Ac | 98.0 ± 0.0 Aab | 52.7 ± 7.0 Ba | 0.7 ± 1.2 Ac | 53.3 ± 6.1 Ba |
| 400 | 89.3 ± 4.2 Aa | 0.0 ± 0.0 Ac | 89.3 ± 4.2 Ab | 26.7 ± 18.1 Bb | 22.7 ± 13.3 Bb | 49.3 ± 5.0 Ba |
| 800 | 61.3 ± 7.6 Ab | 32.0 ± 10.6 Ab | 93.3 ± 3.1 Ab | 4.0 ± 5.3 Bc | 45.3 ± 1.2 Aa | 49.3 ± 4.2 Ba |
| 1000 | 36.0 ± 9.2 Ac | 48.7 ± 5.8 Aa | 84.7 ± 4.2 Ab | 1.3 ± 1.2 Bc | 49.3 ± 10.1 Aa | 50.7 ± 9.0 Ba |
| 1400 | 19.3 ± 12.1 Ad | 50.7 ± 9.0 Aa | 70.0 ± 11.1 Ac | 0.0 ± 0.0 Bc | 43.3 ± 3.1 Aa | 43.3 ± 3.1 Ba |
| NaCl Concentration (mM) | Brown Seed | Black Seed |
|---|---|---|
| 0 | 46.2 ± 0.6 Aa | 19.8 ± 2.3 Ba |
| 100 | 43.3 ± 3.1 Aa | 18.4 ± 0.7 Bab |
| 200 | 35.2 ± 5.5 Ab | 14.6 ± 3.1 Bb |
| 400 | 26.2 ± 1.2 Ac | 3.9 ± 4.1 Bc |
| 800 | 12.6 ± 2.3 Ad | 0.5 ± 0.6 Bcd |
| 1000 | 7.4 ± 2.8 dAe | 0.2 ± 0.2 Bcd |
| 1400 | 5.3 ± 4.2 Ae | 0.0 ± 0.0 Ad |
| NaCl Concentration (mM) | Seedlings from Brown Seeds | Seedlings from Black Seeds |
|---|---|---|
| 0 | 100.0 ± 0.0 Aa | 100.0 ± 0.0 Aa |
| 100 | 96.5 ± 8.3 Aab | 99.4 ± 12.4 Aa |
| 200 | 104.4 ± 12.1 Aa | 90.8 ± 11.8 Aa |
| 400 | 81.8 ± 12.2 Ab | 89.1 ± 10.8 Aa |
| 800 | 15.9 ± 0.3 Ac | 6.8 ± 0.4 Bb |
| 1000 | 11.8 ± 1.5 Ac | 0.0 ± 0.0 Bb |
| 1400 | 1.8 ± 0.1 Ac | 0.0 ± 0.0 Bb |
| NaCl Concentration (mM) | Seedlings from Brown Seeds | Seedlings from Black Seeds |
|---|---|---|
| 0 | 100.0 ± 0.0 Ab | 100.0 ± 0.0 Aa |
| 100 | 126.6 ± 11.8 Aa | 121.1 ± 19.8 Aa |
| 200 | 122.3 ± 14.4 Aa | 109.3 ± 20.2 Aa |
| 400 | 108.2 ± 10.7 Ab | 109.9 ± 15.2 Aa |
| 800 | 63.5 ± 7.5 Ac | 39.5 ± 4.5 Bb |
| 1000 | 35.1 ± 3.3 Ad | 0.0 ± 0.0 Bc |
| 1400 | 19.7 ± 2.7 Ad | 0.0 ± 0.0 Bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Wang, H.; Chu, R.; Zhao, L.; Li, X.; An, S.; Qiang, M.; Du, W.; Li, Q. Differences in Physiological Characteristics, Seed Germination, and Seedling Establishment in Response to Salt Stress between Dimorphic Seeds in the Halophyte Suaeda liaotungensis. Plants 2023, 12, 1408. https://doi.org/10.3390/plants12061408
Song J, Wang H, Chu R, Zhao L, Li X, An S, Qiang M, Du W, Li Q. Differences in Physiological Characteristics, Seed Germination, and Seedling Establishment in Response to Salt Stress between Dimorphic Seeds in the Halophyte Suaeda liaotungensis. Plants. 2023; 12(6):1408. https://doi.org/10.3390/plants12061408
Chicago/Turabian StyleSong, Jieqiong, Hongfei Wang, Ruowen Chu, Lantong Zhao, Xinxin Li, Shuo An, Mengke Qiang, Wanying Du, and Qiuli Li. 2023. "Differences in Physiological Characteristics, Seed Germination, and Seedling Establishment in Response to Salt Stress between Dimorphic Seeds in the Halophyte Suaeda liaotungensis" Plants 12, no. 6: 1408. https://doi.org/10.3390/plants12061408