2.1. Germination under Drought Stress and Silicon Application
There were clear differences between the germination energy (GE) of the individual species depending on the conditions of the induced drought as well as between the species under the same germination conditions (
Table 1). The seeds of
Lolium perenne alone and in a mixture with
Medicago x varia germinated the best under the control conditions (distilled water) (average 82.4%), whereas under the drought conditions, the GE decreased distinctly, especially when silicon was applied (the worst was at −0.6 MPa + Si, 29%). In addition, the
M. x varia seeds showed similar GE values regardless of the germination conditions (approx. 51–63%).
It was found that when the seeds of both species were germinated together in a mixture, the L. perenne showed higher GE values only under the control conditions and also when silicon (Si) was used in addition to water. The drought conditions inhibited the GE of this species to a greater extent than the seeds of L. perenne and M. x varia germinated separately. In contrast, the M. x varia seeds in such conditions showed better GE (56.4–79.5%) values, not only compared to L. perenne but also compared to germination in the control conditions and with Si addition (15.4 and 28.2%, respectively). As a result, the germination capacity (GC) was similar in the individual species, regardless of the germination conditions. For L. perenne, the GE value was at a high level of 88–100%, and for M. x varia it was 62.7–74.7%. In addition, when the seeds of both species were germinated as a mixture, the GC of L. perenne remained similar and high (85–97%), whereas, the seeds of M. x varia in the control conditions with Si addition showed significantly lower values of GC (F = 28.236; p < 0.000) than the other variants.
The
L. perenne seedlings were considered normal in 88 to about 99% of the germination variants without silicon (
Table 2). However, when Si was used with water (control + Si), seedlings that were not properly developed constituted 24%. The use of Si significantly increased the number of abnormal seedlings (
F = 33.159;
p < 0.000), especially under the drought conditions, i.e., −0.6 MPa + Si (73.3%). At the same time, seeds that did not germinate under drought conditions accounted for only 5–12%. A similar regularity was observed in the
M. x varia seedlings. Under the drought conditions, abnormal seedlings accounted for about 31–39%, while drought conditions and Si application increased their number (36 to 57% of the germinated seeds). Moreover, a significant share of healthy, non-germinating, and dead seeds were also observed, regardless of the germination variant (27–37%). The germination of both species as a mixture showed that normal seedlings of
L. perenne accounted for as much as 85–97% (similar values regardless of the conditions). The share of normal seedlings of
M. x varia was 14% lower in the drought conditions except for D2 + Si, where abnormal seedlings accounted for as much as 44%.
Marked changes in the number and length of roots and leaves developed by both the species under the drought conditions were found (
Table 3 and
Table 4). The
L. perenne seedlings developed longer roots in the drought conditions, especially in the −0.3 MPa variant (an increase of 36% compared to the control conditions). At the same time, the use of Si significantly inhibited the growth of roots (they were shorter by 30 mm compared to the control conditions and by approx. 53 mm under the drought conditions) (
F = 49.678;
p < 0.000). The
M. x varia reacted similarly to the drought conditions and developed seedlings with longer roots, especially at −0.3 MPa (47 mm), but the use of silicon during germination significantly reduced the length of the roots (up to 13 mm independent of the drought variant) (
F = 16,354;
p < 0.000). A similar reaction of both species to the application of drought conditions (longer roots of seedlings) and silicon application (shorter roots of seedlings) was also found. The drought conditions and the use of silicon also affected the length of the leaves of the
L. perenne and
M. x varia seedlings (when the seeds of both species were germinated as a mixture) but to a lesser extent than the length of roots. At the same time, these conditions had the least effect on the number of roots and leaves developed by the seedlings that were germinated separately and as a mixture.
The fresh weight of the seedlings of both species under the drought conditions was greater compared to the control conditions (
Table 5). However, the use of silicon under the control or drought conditions resulted in a lower fresh mass. The application of silicon in the control conditions resulted in a decrease in the fresh weight of the seedlings, especially
L. perenne (by approx. 40%). In turn, when the seeds of
L. perenne and
M. x varia were germinated separately under the drought conditions, the seedlings were characterized by a significantly higher dry weight (
F = 6.319;
p < 0.004), and the same was observed after the application of Si. When the seeds of both species were germinated as a mixture, no differences were found in the seedling weight compared to when they were germinated separately.
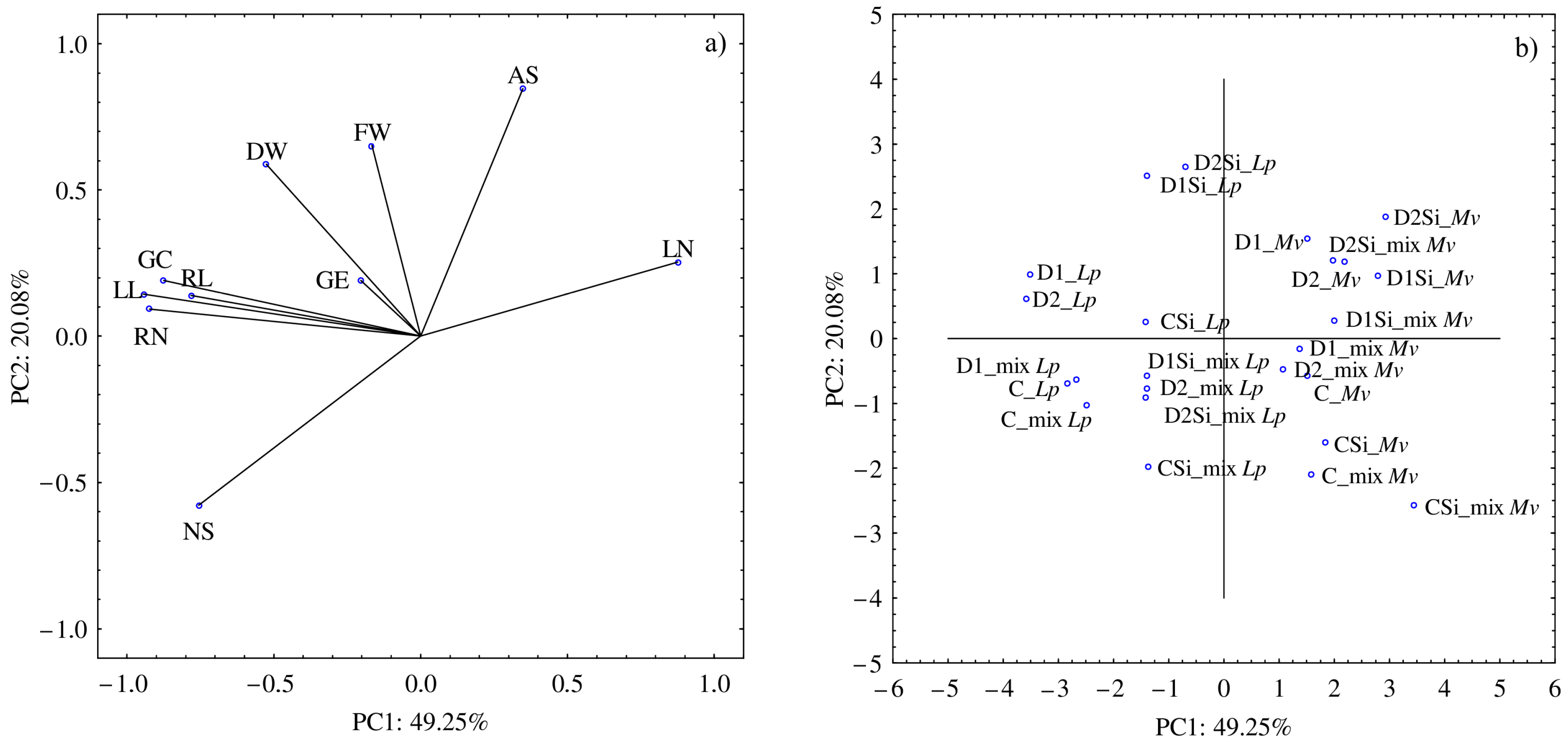
The PCA analysis showed that the first component accounted for 49.25% and the second accounted for 20.08% of the analyzed variability (
Figure 1a,b). The water conditions and the application of Si affected the GC, RN, RL, and LL the most. These parameters were significantly and positively correlated with each other. Moreover, a high NS was associated with a low LN. The highest values of NS were recorded in the
L. perenne in the mixture both in the control + Si and drought + Si conditions. In addition, the drought and Si conditions, especially D2 + Si, stimulated the development of AS in the
M. x varia. Simultaneously, the drought conditions combined with the application of Si resulted in a higher dry weight of the
L. perenne seedlings.
2.2. Germination with the addition of Root Extracts and with Variable pH
A clear effect of applying
M. sativa root extracts and changing the pH of the solution on the GE and GC of the
L. perenne and
M. sativa seeds, both alone and as a mixture, was found (
Table 6).
After five days, the GE of the L. perenne seeds under the control conditions (distilled water) was over 70% and was significantly higher compared to the remaining variants (15.5–53.5%). It was also found that the use of an alkaline solution (pH 6.5) in combination with a root extract of a lower concentration (LpH 6.5) contributed to a better GE for the L. perenne (average 53%) than under the conditions of the other variants with root extracts and solutions, and these results were significant compared to the application of a high concentration of extract (H). However, the GE of the M. sativa seeds in the conditions of the appropriate variants was higher than in the case of L. perenne (except for the control conditions) and varied from 53% to 68%. The effect of the root extracts and the pH of the solution was also visible in the case of the mixture. The GE of the L. perenne seeds was higher with an alkaline pH (62%) compared to the other variants of germination. In addition, the seeds of this species were characterized by a significantly better GE at a lower extract concentration (L) and at a higher extract concentration in combination with an acidic solution (HpH 5.0) (by about 28% and 25%, respectively) compared to when the seeds were germinated individually.
In turn, the application of alkaline pH in conditions in combination with low concentration of root extract had a negative effect on the GE of the L. perenne seeds. The M. sativa seeds germinated the best under the control conditions (86%) and with both root extract applications (59–62%). The use of an acidic solution caused the worst GE values, especially in combination with a high concentration of root extract (20%), whereas the GC of the L. perenne seeds was the highest under the control conditions and amounted to about 90%. The seeds germinated slightly worse in the solution with a pH of 6.5, and the same was observed in the combination with a low concentration of root extract (82% and 84.5%, respectively). On the other hand, the use of an acidic solution (pH 5.0) in combination with the root extract limited the germination of the seeds (an average of 61%). In turn, the GC of M. sativa was visibly lower (70.5%) in comparison with L. perenne. Moreover, the use of a high concentration of root extract in combination with an acidic solution (HpH 5.0) and especially an alkaline solution (HpH 6.5) caused a marked reduction in the germination of the seeds (by 10% and 17%) compared to the control conditions. However, when the seeds of both species were germinated as a mixture, the GC of L. perenne was significantly worse (73%), while that of M. sativa was higher (90%) in the control conditions. In the remaining variants, the GC of the L. perenne seeds was higher than that of M. sativa (except for with a high concentration of extracts). Simultaneously, the GC of the M. sativa seeds in the mixture with L. perenne was similar to that of the seeds of these species when they were germinated separately (except for control conditions).
Large differences in the morphology of seedlings of the assessed species were found, and these depended on the variants used during seed germination (
Table 7). A lower concentration of
M. sativa root extract (L) had a positive effect on the length of the
L. perenne and
M. sativa roots, both when germinated individually andas a mixture. The application of the remaining variants during the germination of the
L. perenne seeds caused a considerable reduction in the length of the roots (except under the control conditions). The shortest roots were developed when an acidic solution was used, especially in the LpH 5.0 variant (approx. 15 mm), whereas, the growth of
M. sativa was stimulated under both root extract applications, and the length of the roots varied from 54 to 60 mm. The other variants used during germination resulted in a clearly limited root growth. A significant stimulating effect of the root extracts was also found according to the germination of
L. perenne and
M. sativa as a mixture, especially for the application of a low concentration of extract (the roots were longer by 115% and 23% compared to the control, respectively).
However, the use of the remaining variants had an inhibitory effect on the growth of the roots of M. sativa, where they were significantly shorter, and this was also observed compared to the control. The length of the L. perenne leaves, as in the case of the roots, was the greatest for the application of a low concentration of extract (57 mm) as well as for the control conditions (about 54 mm). It was found that the use of the remaining variants inhibited the growth of leaves but to a lesser extent than that of roots. The shortest leaves were developed under acidic conditions of the solution and under acidic conditions combined with the application of a low concentration of extract (LpH 5.0). However, the length of the M. sativa leaves was the greatest when the root extracts were used (regardless of the concentration). On the other hand, the seedlings obtained under alkaline or acidic solution conditions led to the development of shorter leaves compared to the other variants. A beneficial effect of the root extracts on the leaf length was also noted in the case of seed germination as a mixture, but only for L. perenne.
The effects of the applied conditions on the fresh and dry weight of the seedlings were found (
Table 8). The fresh weight of the
L. perenne seedlings was the highest when the seeds were germinated under the conditions of a low concentration of
M. sativa root extract (it was 25% higher compared to the control conditions). However, the use of this extract in combination with an acidic solution resulted in a significantly lower weight (about 13 mg per plant). In turn, the fresh weight of
M. sativa was negatively affected by both the higher concentration of the extract and the acidic solution (this was significant compared to the control conditions). In the case of the germination the seeds of both species as a mixture, the use of the
M. sativa root extract had a significantly positive effect on the fresh weight of the
L. perenne seedlings, while the use of an acidic solution had a negative effect. On the other hand, the
M. sativa seedlings obtained a significantly higher weight when a high concentration of extract was used (compared to the control conditions).
The dry weight of the L. perenne seedlings was similar (it ranged from 2.77 to 3.43 mg per plant), except for the pH 5.0 variant, where it was significantly higher (4.13 mg per plant). Moreover, the M. sativa seedlings reached the highest weight (significant in relation to the control conditions) with the use of an alkaline solution, and the lowest was obtained with the use of a high concentration of extract and in the HpH 6.5 conditions. In the mixture, a beneficial effect of a low concentration of extract in combination with an acidic solution (LpH 5.0) was found only in terms of the dry weight of the L. perenne plants, while the weight of the M. sativa seedlings was similar for all the germination variants (2.05–2.72 mg per plant).
The results obtained showed that the GC of
L. perenne as well as
M. sativa, especially when germinated as a mixture with
L. perenne, were inhibited in response to the different concentrations of
M. sativa root extract and the pH of the solution (
Table 9). Similarly, the high concentration of extract induced a decrease in the RL and LL of the seedlings of
M. sativa and
L. perenne in the mixture. However, the treatment with a low concentration of extract caused a visible increase in the RL as well as LL (except for
M. sativa in a mixture). Moreover, all the extracts and solutions stimulated the DW of
M. sativa (except the treatment with a high concentration of extract) as well as the variant with
Ms germinated as a mixture.
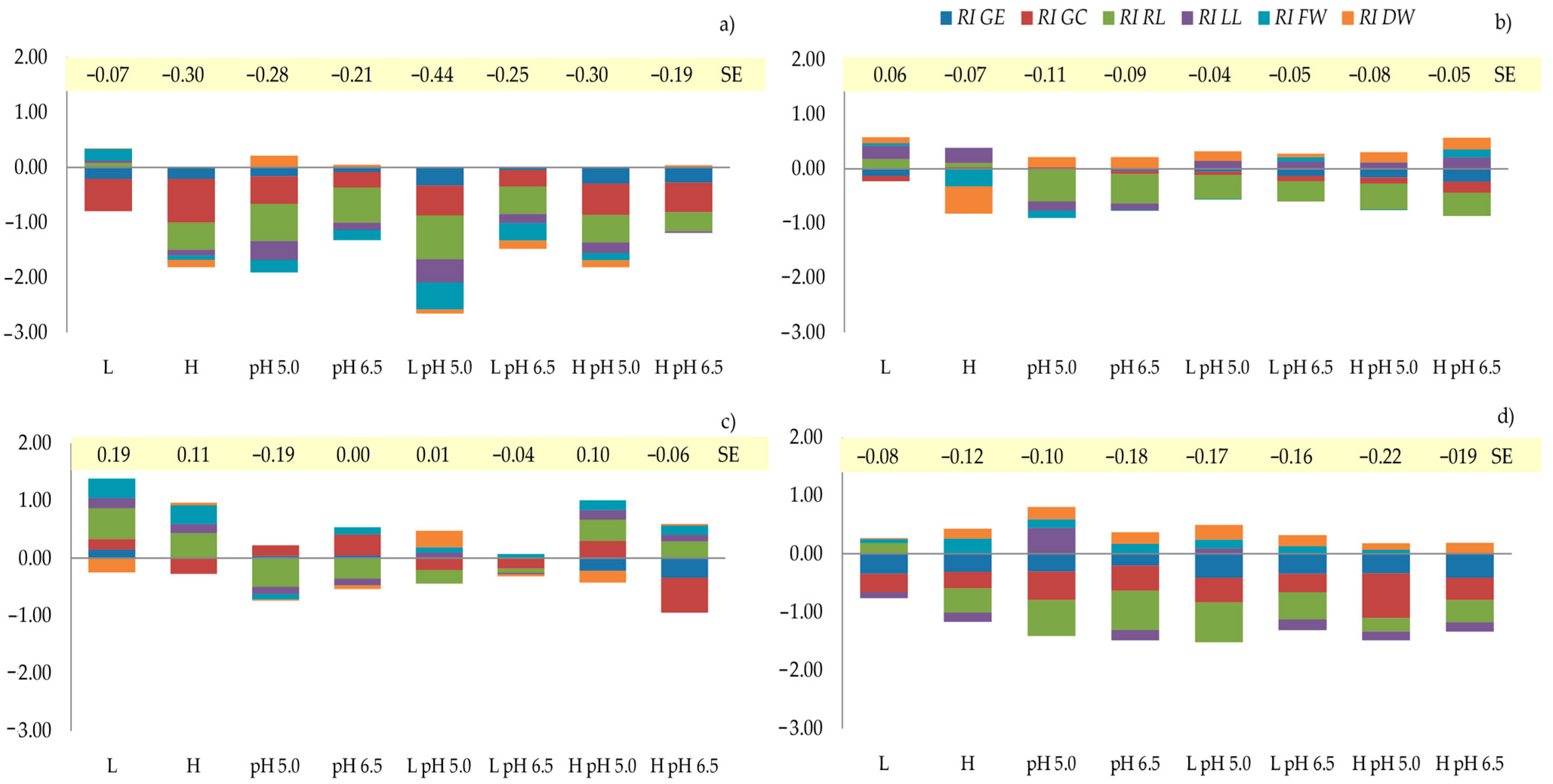
An allelopathic synthesis effect (
SE) was obtained for each assessed plant (which was also observed when the plants were germinated as a mixture) by combining the
RI values of the seed germination and growth of the seedlings (
Figure 2). The values of this indicator were low, with the highest ones being found for
L. perenne. It was found that the S
E value for
L. perenne was negative for all the variants, which was mainly determined by the
RI RL and
RI GC indices. In turn, for the
L. perenne germinated in a mixture, the highest values of
SE (negative and positive) depended primarily on the
RI RL index (
Table 9,
Figure 2). For
M. sativa, the
SE values were only positive with the variant where a low concentration of the extract was used, despite the inhibitory effect of this extract on seed germination. On the other hand, in the mix
Ms variant, all the
SE values were negative despite the beneficial impact of evaluated variants on the FW and DW of the seedlings.





{kind=link}
{kind=link}