
Conocarpus lancifolius (Combretaceae): Pharmacological Effects, LC-ESI-MS/MS Profiling and In Silico Attributes


 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Sample Preparation
2.2. Chemicals and Reagents
2.3. Estimation of Total Phenolic Content (TPC)
2.4. Estimation of Total Flavonoid Content (TFC)
2.5. Antioxidant Assays
2.5.1. DPPH Radical Scavenging Assay
2.5.2. Total Antioxidant Capacity (TAC)
2.5.3. Total Reducing Power Assay
2.6. Acetylcholinesterase (AChE) Inhibitory Activity
2.7. Animals and Their Housing Conditions
2.8. Acute Toxicity Study
2.9. Subacute Toxicity Study
2.10. Cardioprotective Studies
2.10.1. Isoproterenol-Induced Chronic Myocardial Infarction (MI)
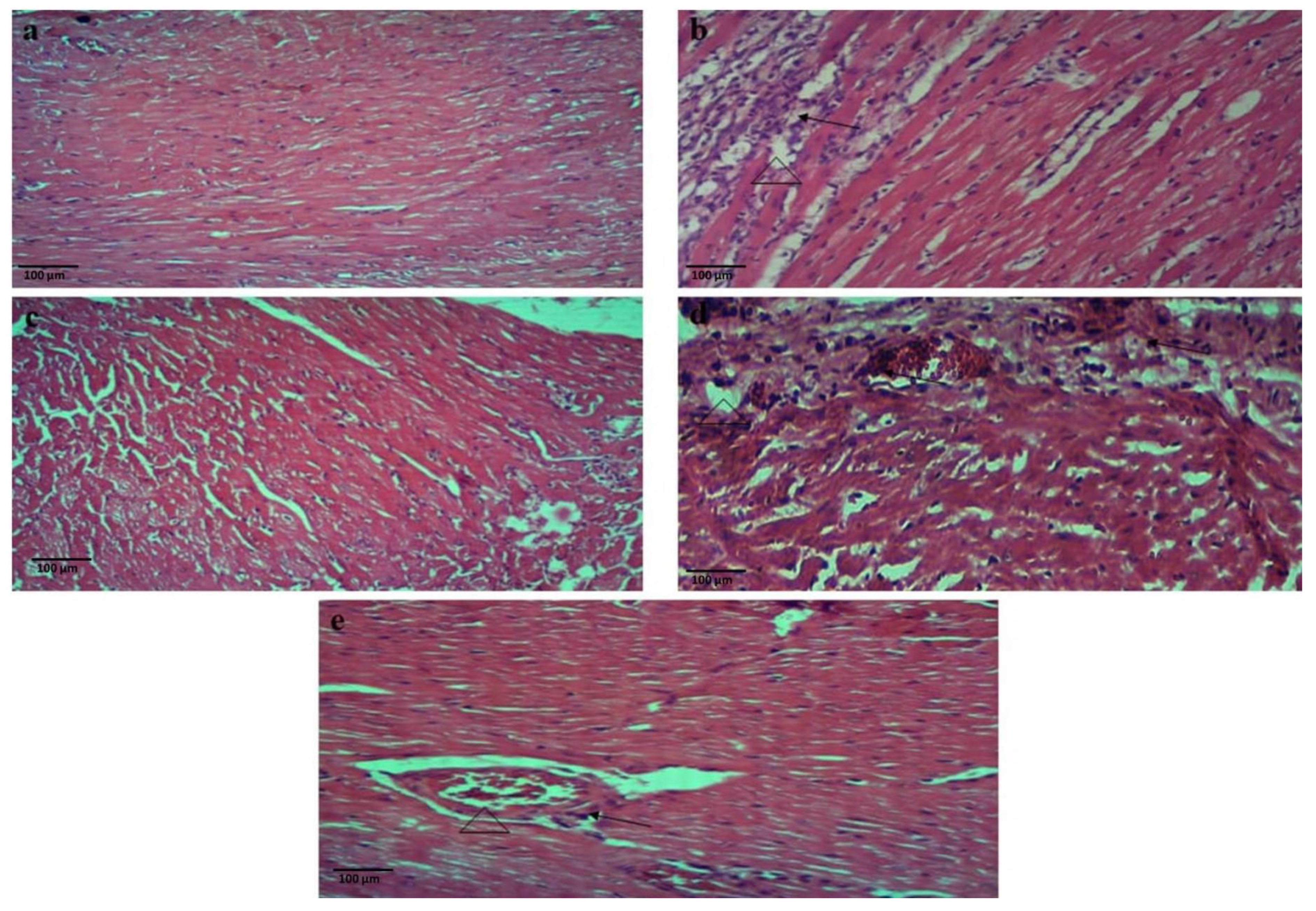
2.10.2. Blood Collection and Histopathological Examinations
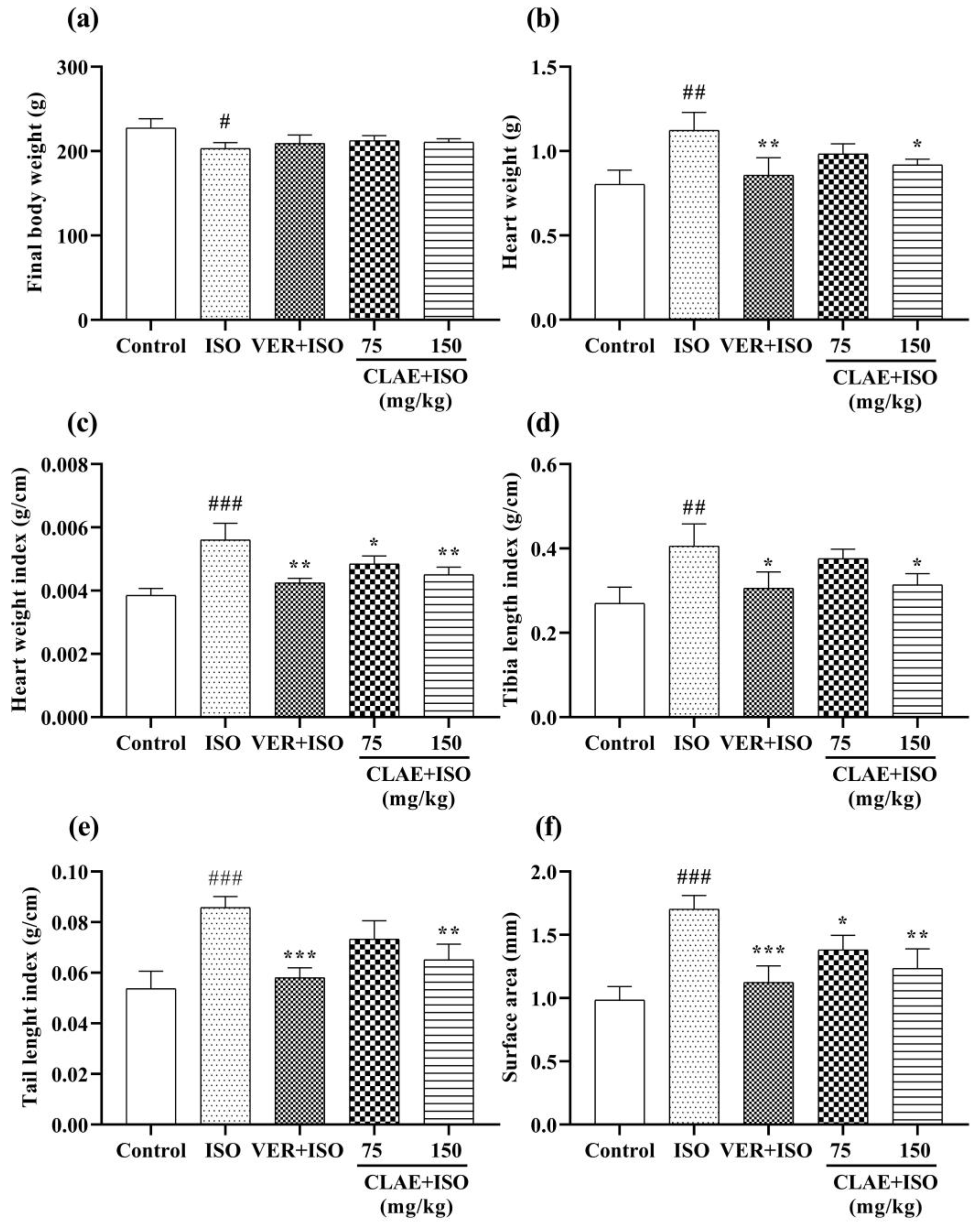
2.10.3. Measurement of Cardiac Biometric Indices
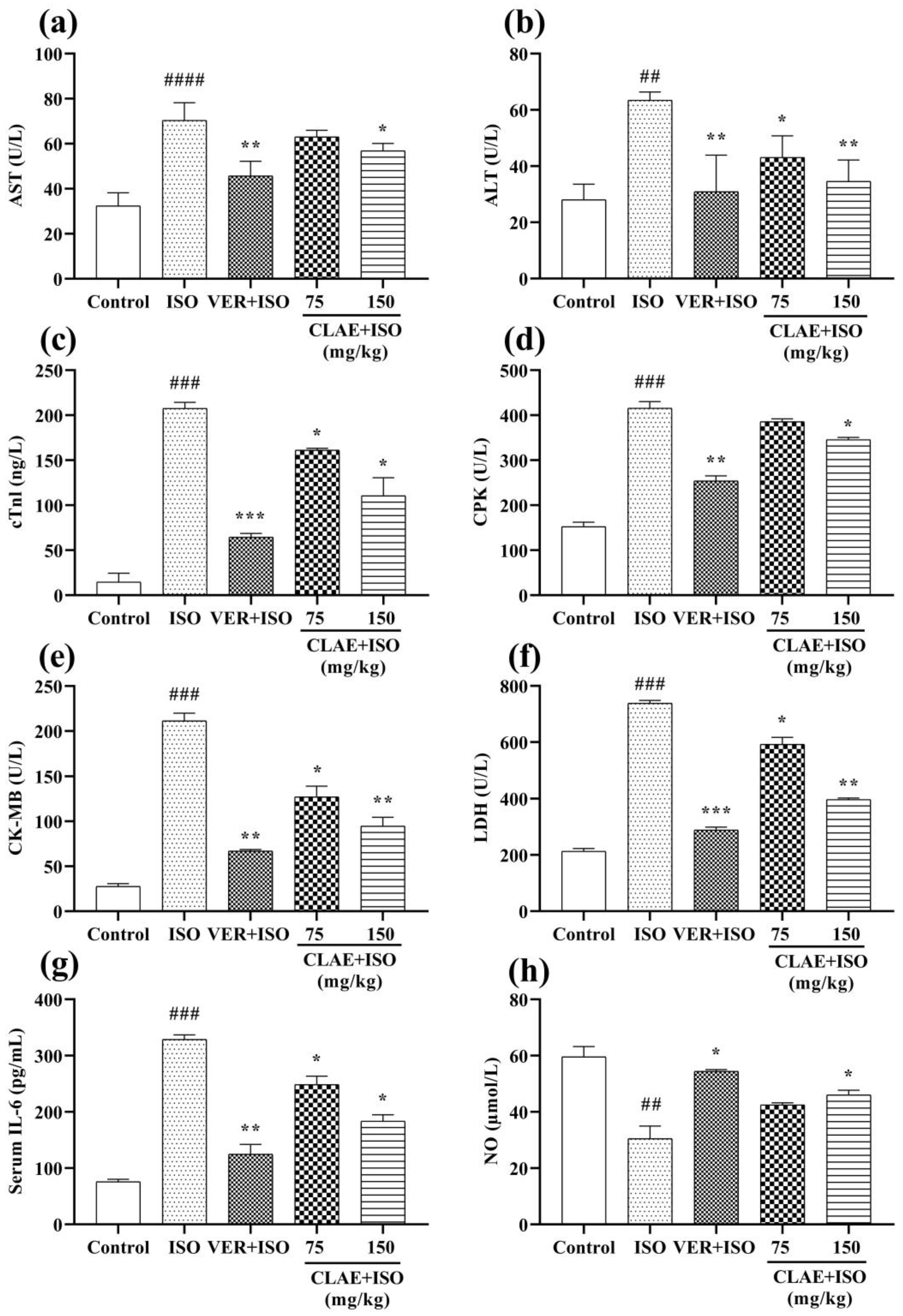
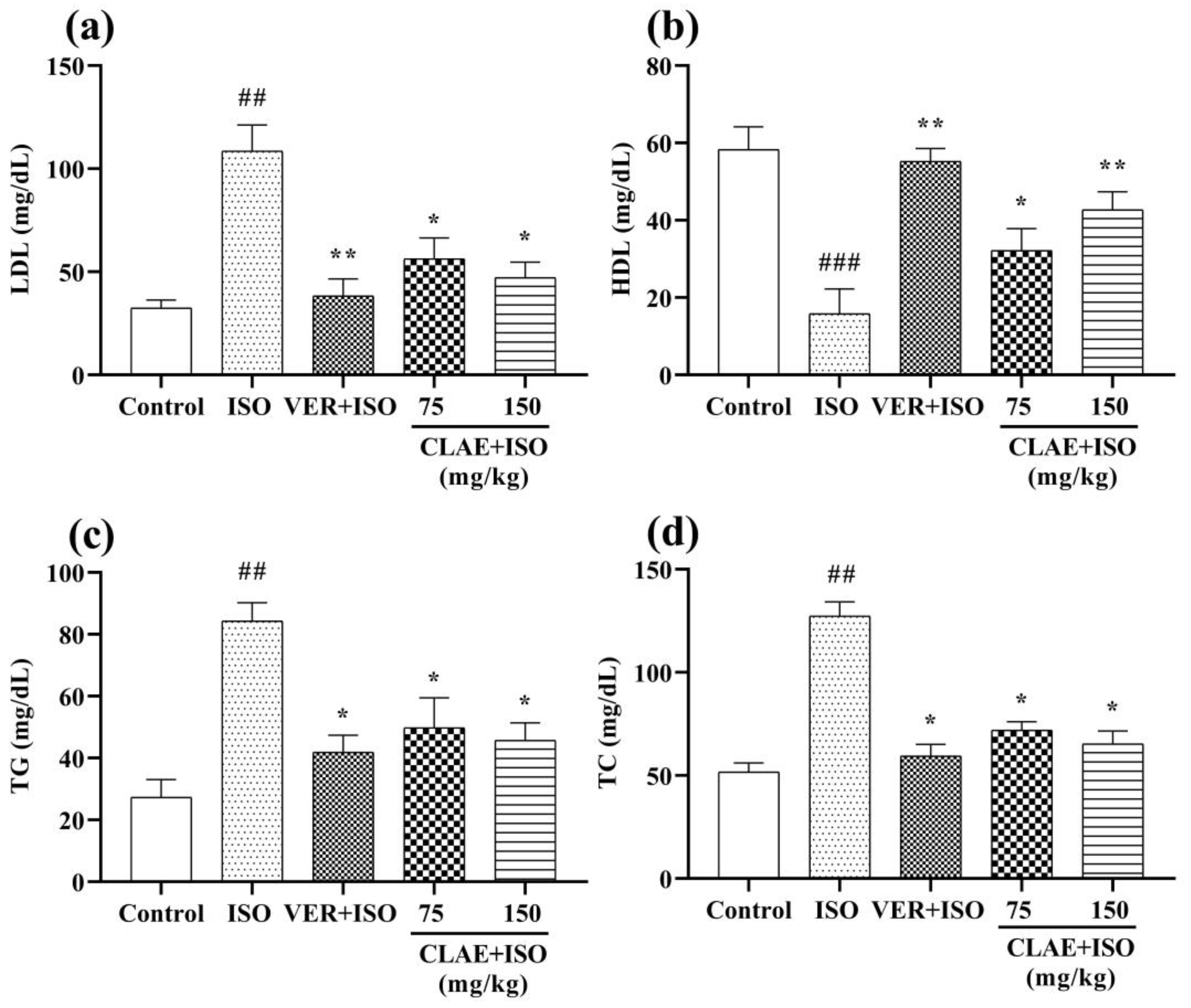
2.10.4. Estimation of Heart Biochemical Markers
2.11. Behavior Studies
2.11.1. Experimental Design
2.11.2. Noise Stress Induction Procedure
2.12. Behavior Studies for Anxiety
2.12.1. Open Field Test (OFT)
2.12.2. Elevated Plus Maze (EPM) Test
2.13. Behavior Studies for Depression
Forced Swimming Test (FST)
2.14. Behavior Studies for Memory and Learning
2.14.1. Novel Object Recognition (NOR) Test
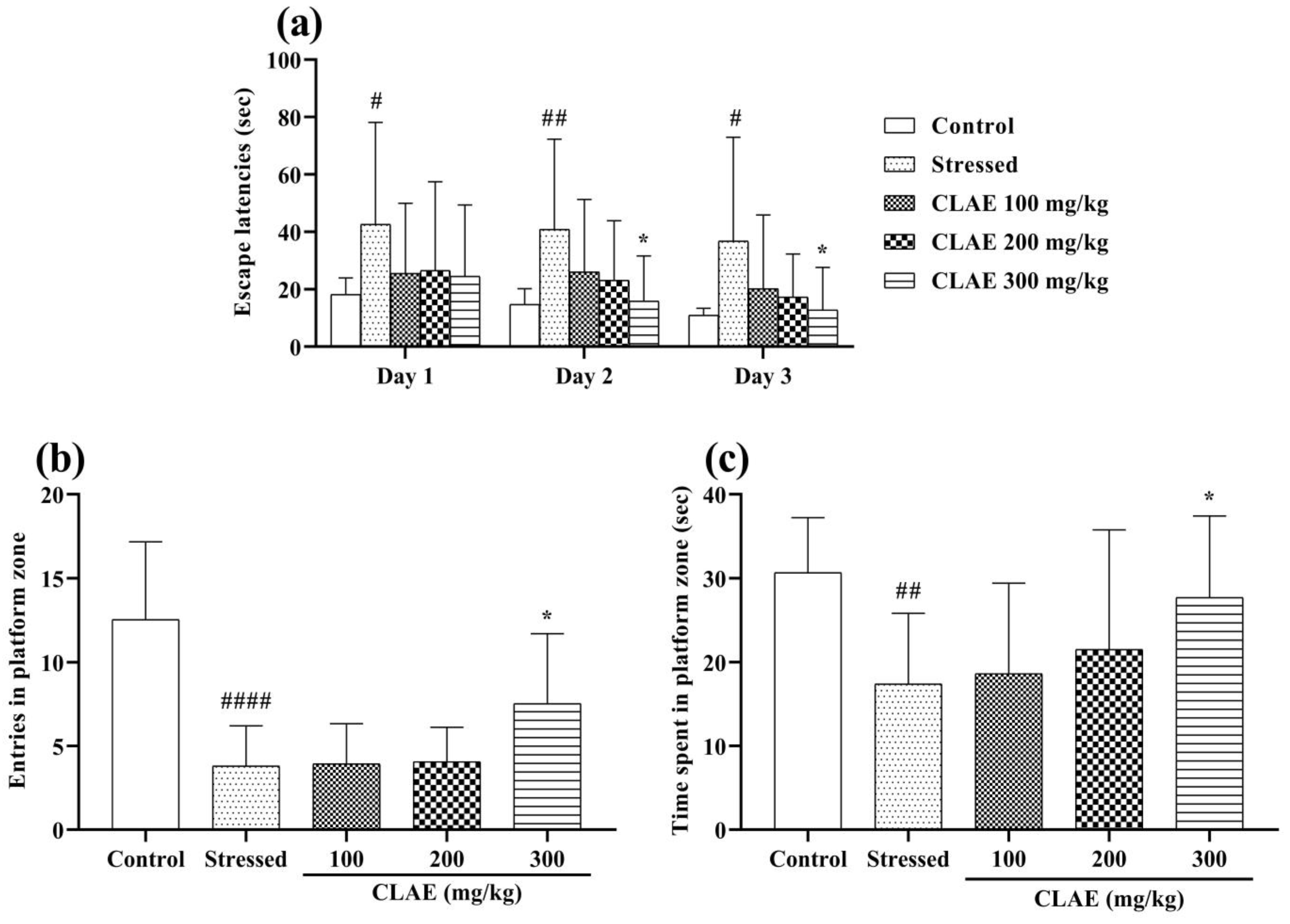
2.14.2. Morris Water Maze (MWM) Test
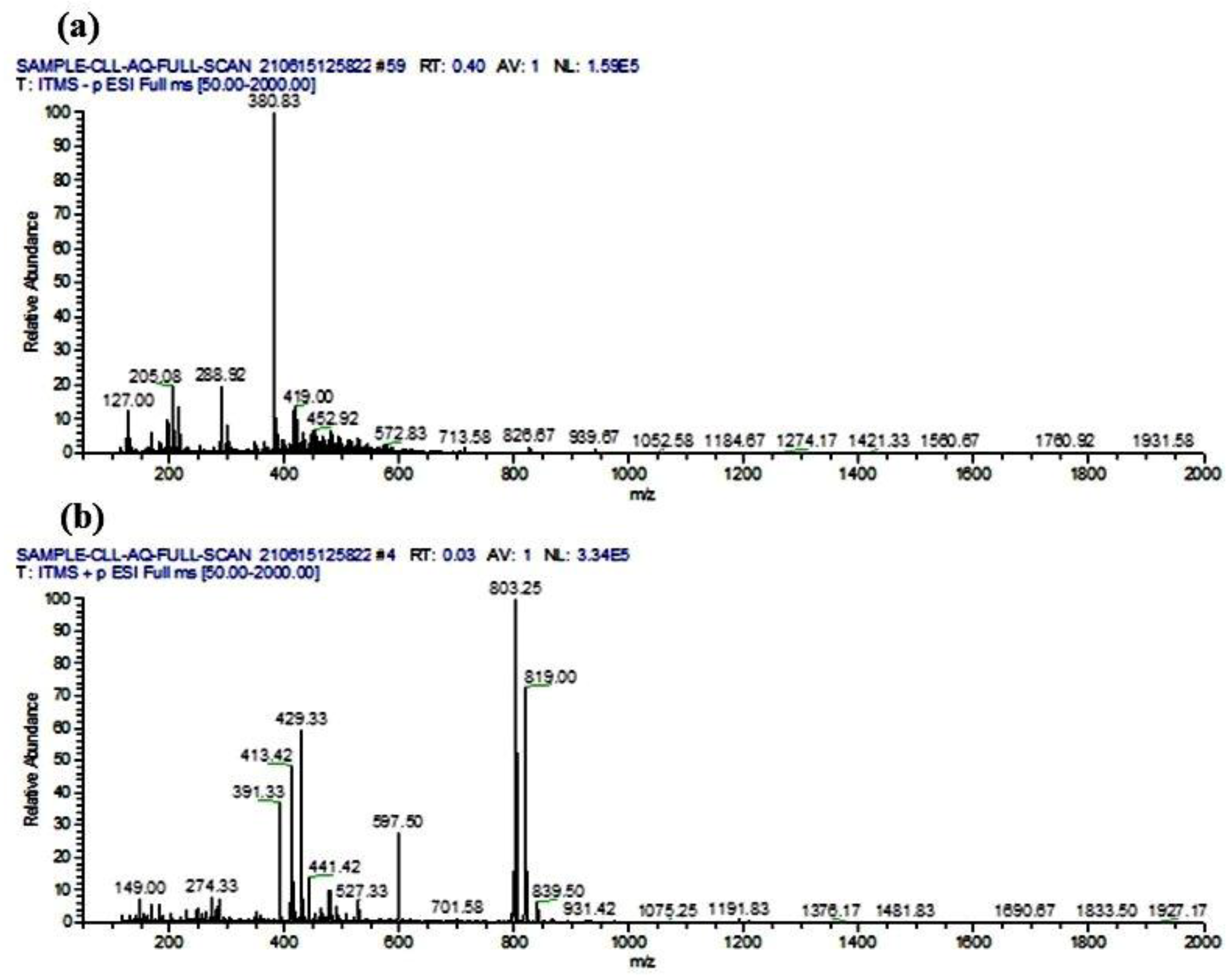
2.15. LC-ESI-MS/MS Analysis
2.16. Drug-Likeness and ADMET Predictions
2.17. Molecular Docking Approach
2.17.1. Ligand Preparation
2.17.2. Protein Preparation
2.17.3. Receptor Grid Generation and Molecular Docking
2.17.4. Inhibition Constant (Ki)
2.18. Network Analysis
2.18.1. Screening of Ingredients-Related and Disease-Associated Potential Targets
2.18.2. Enrichment Analysis and Network Construction
2.19. Statistical Analysis
3. Results
3.1. Determination of TPC, TFC and Antioxidant Capacities
3.2. AChE Inhibitory Activity
3.3. Single-Dose Acute Toxicity
3.4. Repeated-Dose Subacute Toxicity
3.5. ISO-Induced Chronic Myocardial Injury
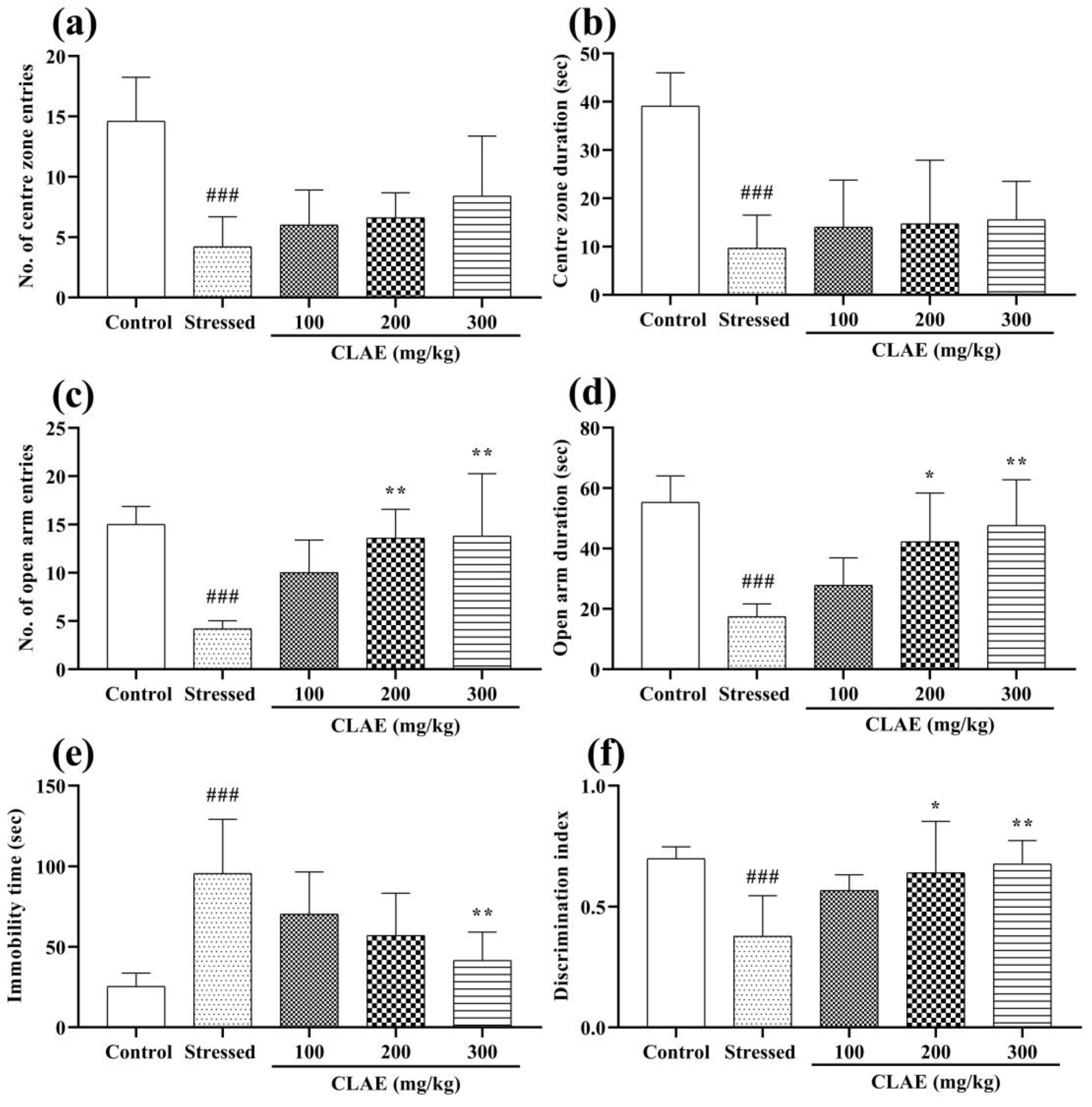
3.6. Behavior Studies for Anxiety
3.6.1. Open Field Test (OFT)
3.6.2. Elevated Plus Maze (EPM) Test
3.7. Behavior Studies for Depression
Forced Swimming Test (FST)
3.8. Behavior Tests for Memory and Learning
3.8.1. Novel Object Recognition (NOR) Test
3.8.2. Morris Water Maze (MWM) Test
3.9. Phytocompounds Identification Using LC-ESI-MS/MS Analysis
3.10. Drug-Likeness and ADMET Studies
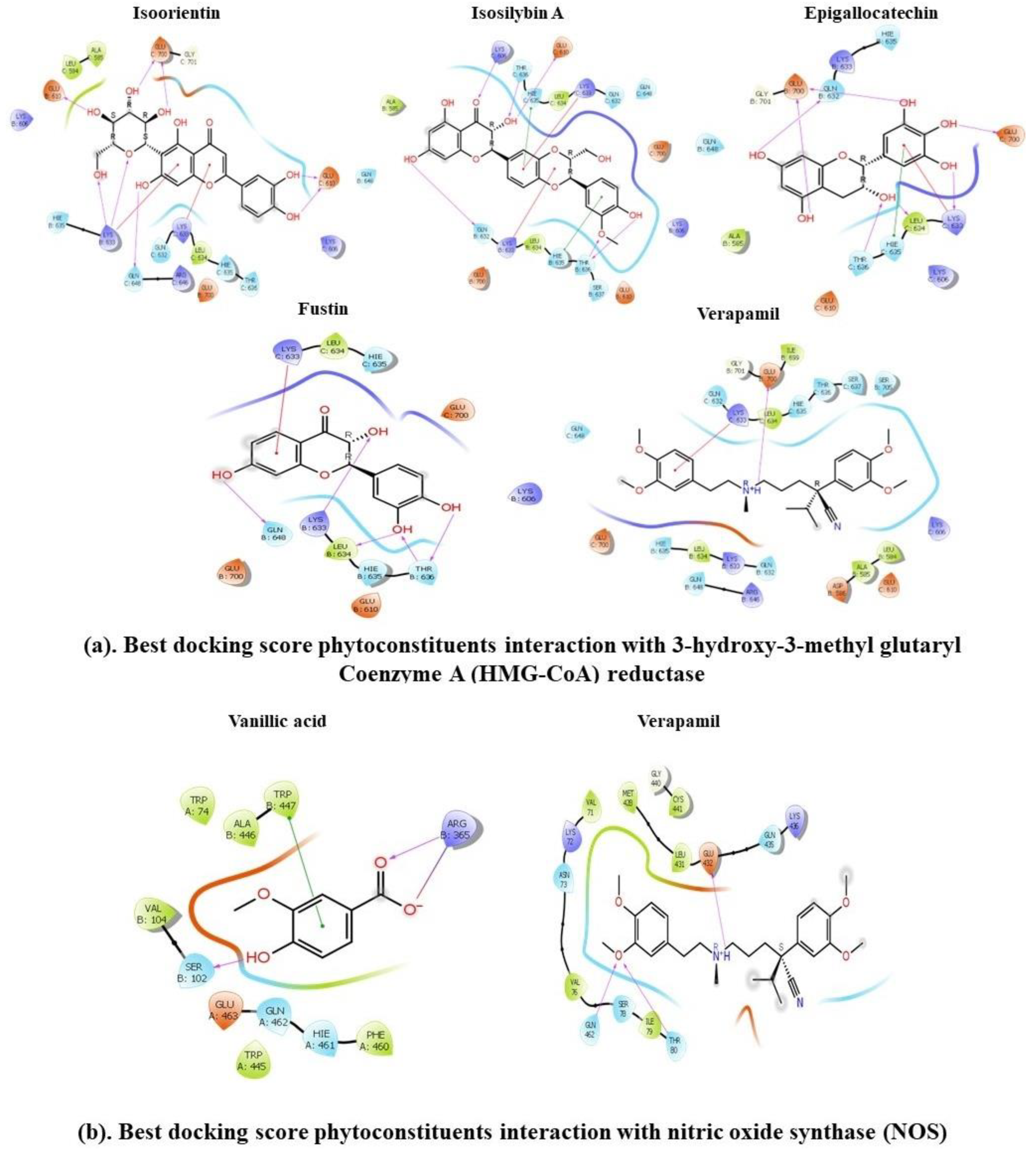
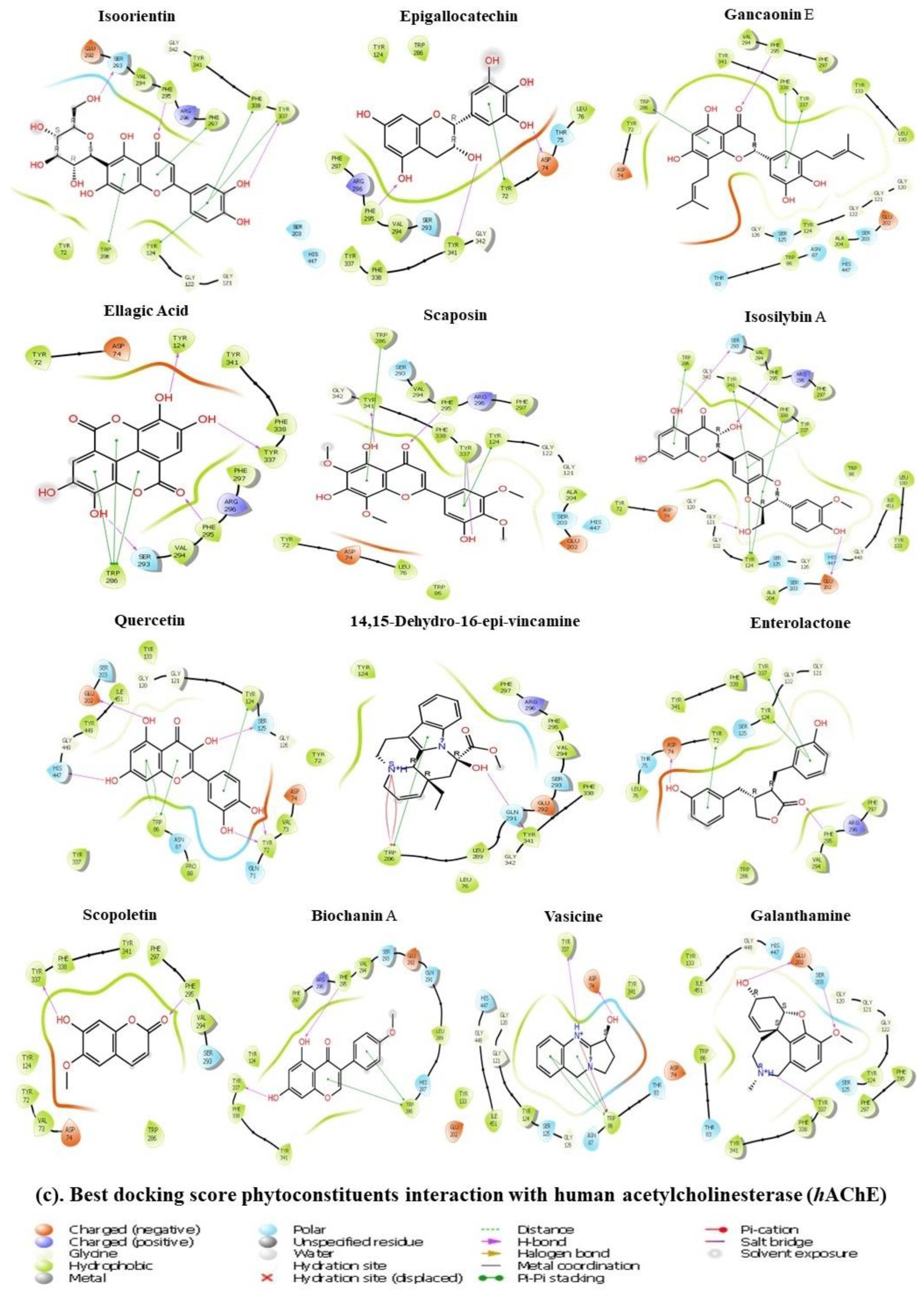
3.11. Molecular Docking Approach
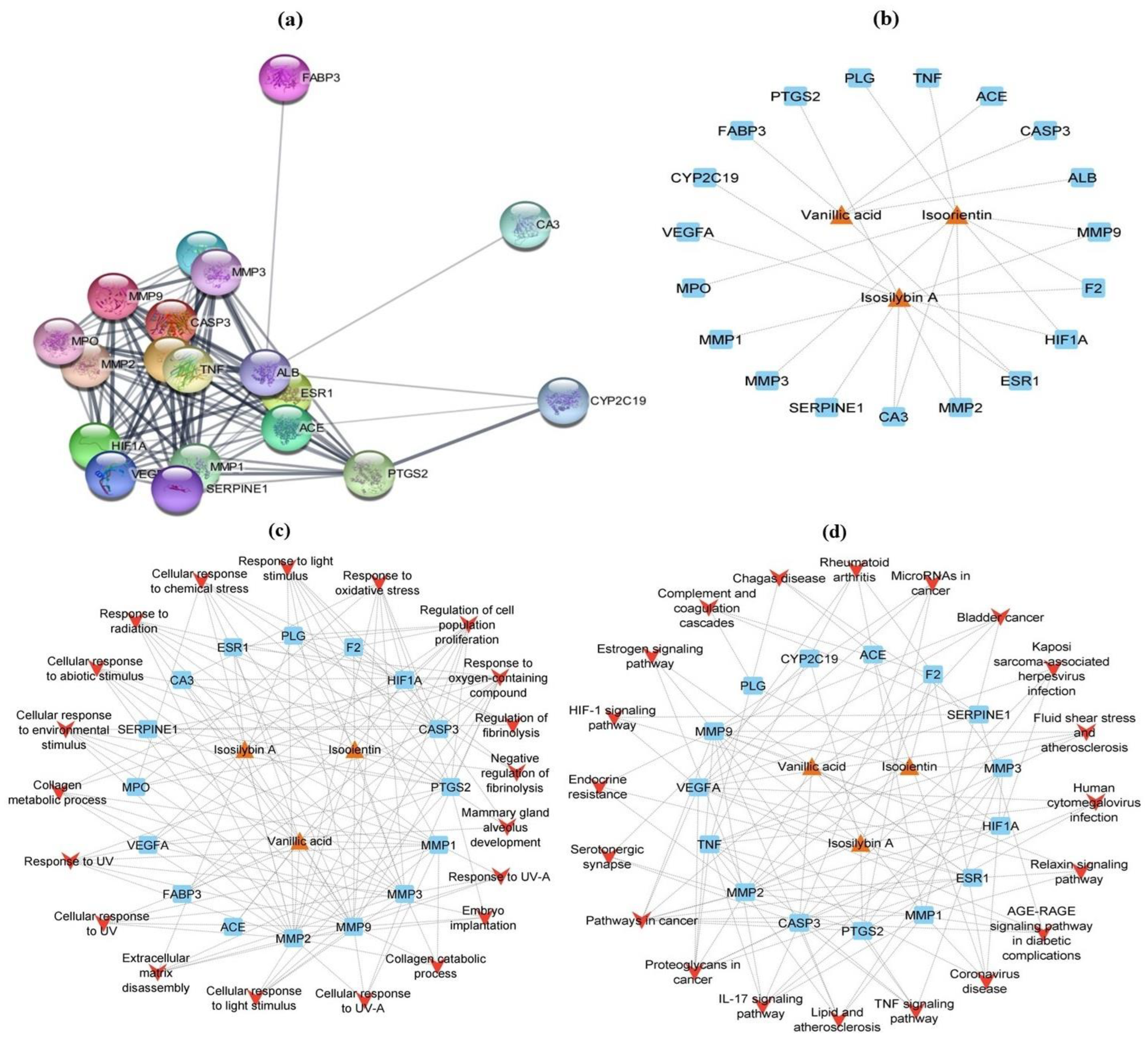
3.12. Network Analysis
3.12.1. Screening of Putative Targets
3.12.2. GO and KEGG Enrichment Analysis
3.12.3. Network Construction
4. Discussion
4.1. TPC, TFC and Antioxidant Capacities
4.2. Single-Dose Acute Toxicity
4.3. Repeated-Dose Subacute Toxicity
4.4. ISO-Induced Chronic Myocardial Injury
4.5. Behavioral Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaduganathan, M.; Mensah, G.A.; Turco, J.V.; Fuster, V.; Roth, G.A. The Global Burden of Cardiovascular Diseases and Risk: A Compass for Future Health. J. Am. Coll. Cardiol. 2022, 80, 2361–2371. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Pandey, M.M.; Rawat, A. Traditional herbs: A remedy for cardiovascular disorders. Phytomedicine 2016, 23, 1082–1089. [Google Scholar] [CrossRef]
- Samad, N.; Yasmin, F.; Khaliq, S.; Ahmad, S.; Azizuddin; Mustafa, S. Diet supplements of banana fruit pulp mitigates repeated noise stress induced behavioral deficits and oxidative stress. Pak. J. Pharm. Sci. 2020, 33, 2293–2300. [Google Scholar]
- Sikandaner, H.E.; Park, S.Y.; Kim, M.J.; Park, S.N.; Yang, D.W. Neuroprotective effects of sildenafil against oxidative stress and memory dysfunction in mice exposed to noise stress. Behav. Brain Res. 2017, 319, 37–47. [Google Scholar] [CrossRef]
- Muhorakeye, O.; Biracyaza, E. Exploring barriers to mental health services utilization at Kabutare District Hospital of Rwanda: Perspectives from patients. Front. Psychol. 2021, 12, 638377. [Google Scholar] [CrossRef]
- Malik, H.; Javaid, S.; Fawad Rasool, M.; Samad, N.; Rizwan Ahamad, S.; Alqahtani, F.; Imran, I. Amelioration of scopolamine-induced amnesic, anxiolytic and antidepressant effects of Ficus benghalensis in behavioral experimental models. Medicina 2020, 56, 144. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Parajuli, K. Reactive oxygen species: A key hallmark of cardiovascular disease. Adv. Med. 2016, 2016, 9152732. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodzadeh, Y.; Mahmoudi, J.; Gorgani-Firuzjaee, S.; Mohtavinejad, N.; Namvaran, A. Effects of N-acetylcysteine on noise exposure-induced oxidative stress and depressive-and anxiety-like behaviors in adult male mice. Basic. Clin. Neurosci. J. 2021, 12, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Hovatta, I.; Juhila, J.; Donner, J. Oxidative stress in anxiety and comorbid disorders. Neurosci. Res. 2010, 68, 261–275. [Google Scholar] [CrossRef]
- Khurm, M.; Wang, X.; Zhang, H.; Hussain, S.N.; Qaisar, M.N.; Hayat, K.; Saqib, F.; Zhang, X.; Zhan, G.; Guo, Z. The genus Cassia L.: Ethnopharmacological and phytochemical overview. Phytother. Res. 2020, 35, 2336–2385. [Google Scholar] [CrossRef]
- Afifi, H.S.; Al Marzooqi, H.M.; Tabbaa, M.J.; Arran, A.A. Phytochemicals of Conocarpus spp. as a natural and safe source of phenolic compounds and antioxidants. Molecules 2021, 26, 1069. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.M.A.; Akram, M.; Riaz, M.; Munir, N.; Rasool, G. Cardioprotective potential of plant-derived molecules: A scientific and medicinal approach. Dose-Response 2019, 17, 1559325819852243. [Google Scholar] [CrossRef]
- Khan, I.A.; Hussain, M.; Munawar, S.H.; Iqbal, M.O.; Arshad, S.; Manzoor, A.; Shah, M.A.; Abbas, K.; Shakeel, W.; Syed, S.K. Jasminum sambac: A Potential candidate for drug development to cure cardiovascular ailments. Molecules 2021, 26, 5664. [Google Scholar] [CrossRef] [PubMed]
- Batool, R.; Khan, M.R.; Sajid, M.; Ali, S.; Zahra, Z. Estimation of phytochemical constituents and in vitro antioxidant potencies of Brachychiton populneus (Schott & Endl.) R.Br. BMC Chem. 2019, 13, 32. [Google Scholar] [CrossRef]
- Samad, N.; Tariq, S.; Mahmood, A.; Imran, I.; Alqahtani, F. Reversal of repeated noise stress-induced behavioral, cognitive, neuroendocrine response and oxidative parameters by Abelmoschus esculentus (Okra) root powder in rats. Pak. J. Pharm. Sci. 2020, 33, 1221–1232. [Google Scholar]
- Wang, S.; Yu, Y.; Feng, Y.; Zou, F.; Zhang, X.; Huang, J.; Zhang, Y.; Zheng, X.; Huang, X.-F.; Zhu, Y.; et al. Protective effect of the orientin on noise-induced cognitive impairments in mice. Behav. Brain Res. 2016, 296, 290–300. [Google Scholar] [CrossRef]
- Sundaramahalingam, M.; Ramasundaram, S.; Rathinasamy, S.D.; Natarajan, R.P.; Somasundaram, T. Role of Acorus calamus and alpha-asarone on hippocampal dependent memory in noise stress exposed rats. Pak. J. Biol. Sci. 2013, 16, 770–778. [Google Scholar] [CrossRef]
- Ravindran, R.; Devi, R.S.; Samson, J.; Senthilvelan, M. Noise-stress-induced brain neurotransmitter changes and the effect of Ocimum sanctum (Linn) treatment in albino rats. J. Pharmacol. Sci. 2005, 98, 354–360. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R.; Othman, Z.; Abdul Aziz, C.B. Neuroprotective effects of Tualang honey against oxidative stress and memory decline in young and aged rats exposed to noise stress. J. Taibah Univ. Sci. 2018, 12, 273–284. [Google Scholar] [CrossRef]
- Samad, N.; Imran, A.; Bhatti, S.A.; Imran, I.; Alqahtani, F.; Alasmari, A.F.; Sivandzade, F. Vitamin D2 protects acute and repeated noise stress induced behavioral, biochemical, and histopathological alterations: Possible antioxidant effect. Saudi J. Biol. Sci. 2022, 29, 601–609. [Google Scholar] [CrossRef]
- Ali, H.M.; Salem, M.Z.; Abdel-Megeed, A. In-vitro antibacterial activities of alkaloids extract from leaves of Conocarpus lancifolius Engl. J. Pure Appl. Microbiol. 2013, 7, 1903–1907. [Google Scholar]
- Saadullah, M.; Chaudary, B.A.; Uzair, M. Antioxidant, phytotoxic and antiurease activities, and total phenolic and flavonoid contents of Conocarpus lancifolius (Combretaceae). Trop. J. Pharm. Res. 2016, 15, 555–561. [Google Scholar] [CrossRef]
- Raza, S.A.; Chaudhary, A.R.; Mumtaz, M.W.; Adnan, A.; Mukhtar, H.; Akhtar, M.T. Metabolite profiling and antidiabetic attributes of ultrasonicated leaf extracts of Conocarpus lancifolius. Asian Pac. J. Trop. Biomed. 2020, 10, 353–360. [Google Scholar]
- Al-Tameemi, M.; Turke, T.; Hussain, A.; Abdulhameed, H.R.; Numan, E.M.; Abdolaah, S.S.; Ghiad, S.; Muhammed, A.A.; Ban; Nasir, A. Studies on acute toxicity (LD50) and histopathological effects of methanolic and aqueous Conocarpus lancifolius extracts in mice. Int. J. Pharm. Pharm. Res. 2016, 7, 243–253. [Google Scholar]
- Mohammed, S.A.; Mousa, H.M.; Alwan, A.H. Determination of hemolytic cytotoxicity and antibacterial activity of Conocarpus lancifolius aqueous leaves extract. IOP Conf. Ser. Mater. Sci. Eng. 2019, 571, 012045. [Google Scholar] [CrossRef]
- Saadullah, M.; Chaudary, B.A.; Uzair, M.; Afzal, K. Anti-diabetic potential of Conocarpus lancifolius. Bangladesh J. Pharmacol. 2014, 9, 244–249. [Google Scholar] [CrossRef]
- Al-Taweel, A.M.; Perveen, S.; Fawzy, G.A.; Mehmood, R.; Khan, A.; Khan, S.I. New ellagic acid derivative from the fruits of heat-tolerant plant Conocarpus lancifolius Engl. and their anti-inflammatory, cytotoxic, PPAR agonistic activities. Pak. J. Pharm. Sci. 2016, 29, 1833–1837. [Google Scholar]
- Al-Musayeib, N.M.; Mothana, R.A.; Al-Massarani, S.; Matheeussen, A.; Cos, P.; Maes, L. Study of the in vitro antiplasmodial, antileishmanial and antitrypanosomal activities of medicinal plants from Saudi Arabia. Molecules 2012, 17, 11379–11390. [Google Scholar] [CrossRef]
- Raza, S.A.; Ahmad, M.; Mumtaz, M.W.; Bashir, S.; Ch, A.R. In vitro pharmacological attributes and metabolite’s fingerprinting of Conocarpus lancifolius. Bol. Latinoam. Caribe Plantas Med. Aromat. 2021, 20, 660–671. [Google Scholar] [CrossRef]
- Saadullah, M.; Asif, M.; Sattar, A.; Rehman, K.; Shah, S.; Saleem, M.; Shah, A.; Wajid, M.; Rasool, A.; Uzair, M.; et al. Cytotoxic and antioxidant potentials of ellagic acid derivatives from Conocarpus lancifolius (Combretaceae). Trop. J. Pharm. Res. 2020, 19, 1037–1080. [Google Scholar] [CrossRef]
- Velioglu, Y.; Mazza, G.; Gao, L.; Oomah, B. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Ul-Haq, I.; Ullah, N.; Bibi, G.; Kanwal, S.; Ahmad, M.S.; Mirza, B. Antioxidant and cytotoxic activities and phytochemical analysis of Euphorbia wallichii root extract and its fractions. Iran. J. Pharm. Res. 2012, 11, 241–249. [Google Scholar]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colometric methods. J. Food Drug Anal. 2002, 10, 3. [Google Scholar] [CrossRef]
- Phull, A.R.; Abbas, Q.; Ali, A.; Raza, H.; Kim, S.J.; Zia, M.; Haq, I.U. Antioxidant, cytotoxic and antimicrobial activities of green synthesized silver nanoparticles from crude extract of Bergenia ciliata. Future J. Pharm. Sci. 2016, 2, 31–36. [Google Scholar] [CrossRef]
- Siddhuraju, P.; Mohan, P.; Becker, K. Studies on the antioxidant activity of Indian Laburnum (Cassia fistula L.): A preliminary assessment of crude extracts from stem bark, leaves, flowers and fruit pulp. Food Chem. 2002, 79, 61–67. [Google Scholar] [CrossRef]
- Umamaheswari, M.; Chatterjee, T. In vitro antioxidant activities of the fractions of Coccinia grandis L. leaf extract. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 61–73. [Google Scholar] [CrossRef]
- Mathew, M.; Subramanian, S. In vitro screening for anti-cholinesterase and antioxidant activity of methanolic extracts of ayurvedic medicinal plants used for cognitive disorders. PLoS ONE 2014, 9, e86804. [Google Scholar] [CrossRef]
- Institute of Laboratory Animal Resources: ILAR. Animal Welfare Information Center Newsletter. Available online: https://www.nal.usda.gov/programs/awic (accessed on 8 January 2023).
- Figueredo, K.C.; Guex, C.G.; Reginato, F.Z.; da Silva, A.R.H.; Cassanego, G.B.; Lhamas, C.L.; Boligon, A.A.; Lopes, G.H.H.; de Freitas Bauermann, L. Safety assessment of Morus nigra L. leaves: Acute and subacute oral toxicity studies in Wistar rats. J. Ethnopharmacol. 2018, 224, 290–296. [Google Scholar] [CrossRef]
- Organization for Economic Cooperation and Development: Paris. Available online: http://www.oecd-ilibrary.org (accessed on 11 January 2023).
- Saqib, F.; Ali, A.; Ahmedah, H.T.; Irimie, C.A.; Toma, S.I.; Popovici, B.E.; Moga, M.; Irimie, M. Cardioprotective, hypotensive and toxicological studies of Populus ciliata (Wall. ex Royle). Biomed. Pharmacother. 2021, 142, 112065. [Google Scholar] [CrossRef]
- Selhorst e Silva Beserra, A.M.; Vilegas, W.; Tangerina, M.M.P.; Ascêncio, S.D.; Soares, I.M.; Pavan, E.; Damazo, A.S.; Ribeiro, R.V.; de Oliveira Martins, D.T. Chemical characterisation and toxicity assessment in vitro and in vivo of the hydroethanolic extract of Terminalia argentea Mart. leaves. J. Ethnopharmacol. 2018, 227, 56–68. [Google Scholar] [CrossRef]
- Eladwy, R.A.; Mantawy, E.M.; El-Bakly, W.M.; Fares, M.; Ramadan, L.A.; Azab, S.S. Mechanistic insights to the cardioprotective effect of blueberry nutraceutical extract in isoprenaline-induced cardiac hypertrophy. Phytomedicine 2018, 51, 84–93. [Google Scholar] [CrossRef]
- Elasoru, S.E.; Rhana, P.; de Oliveira Barreto, T.; de Souza, D.L.N.; Menezes-Filho, J.E.R.; Souza, D.S.; Moreira, M.V.L.; Campos, M.T.G.; Adedosu, O.T.; Roman-Campos, D.; et al. Andrographolide protects against isoproterenol-induced myocardial infarction in rats through inhibition of L-type Ca2+ and increase of cardiac transient outward K+ currents. Eur. J. Pharmacol. 2021, 906, 174194. [Google Scholar] [CrossRef]
- Wahid, M.; Saqib, F.; Chicea, L.; Ahmedah, H.T.; Sajer, B.H.; Marc, R.A.; Pop, O.L.; Moga, M.; Gavris, C. Metabolomics analysis delineates the therapeutic effects of hydroethanolic extract of Cucumis sativus L. seeds on hypertension and isoproterenol-induced myocardial infarction. Biomed. Pharmacother. 2022, 148, 112704. [Google Scholar] [CrossRef]
- Yin, F.; Spurgeon, H.A.; Rakusan, K.; Weisfeldt, M.L.; Lakatta, E.G. Use of tibial length to quantify cardiac hypertrophy: Application in the aging rat. Am. J. Physiol. Heart Circ. Physiol. 1982, 243, H941–H947. [Google Scholar] [CrossRef] [PubMed]
- Nalban, N.; Sangaraju, R.; Alavala, S.; Mir, S.M.; Jerald, M.K.; Sistla, R. Arbutin attenuates isoproterenol-induced cardiac hypertrophy by inhibiting TLR-4/NF-κB pathway in mice. Cardiovasc. Toxicol. 2020, 20, 235–248. [Google Scholar] [CrossRef]
- Imran, I.; Javaid, S.; Waheed, A.; Rasool, M.F.; Majeed, A.; Samad, N.; Saeed, H.; Alqahtani, F.; Ahmed, M.M.; Alaqil, F.A. Grewia asiatica berry juice diminishes anxiety, depression, and scopolamine-induced learning and memory impairment in behavioral experimental animal models. Front. Nutr. 2021, 7, 587367. [Google Scholar] [CrossRef]
- Javaid, U.; Javaid, S.; Ashraf, W.; Rasool, M.F.; Noman, O.M.; Alqahtani, A.S.; Majeed, A.; Shakeel, W.; Albekairi, T.H.; Alqahtani, F.; et al. Chemical profiling and dose-dependent assessment of fear reducing and memory-enhancing effects of Solanum virginianum in rats. Dose-Response 2021, 19, 1559325821998486. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.S.; Ashraf, W.; Javaid, S.; Rasool, M.F.; Rahman, H.M.A.; Saleem, H.; Anjum, S.M.M.; Siddique, F.; Morales-Bayuelo, A.; Kaya, S.; et al. Chemical characterization and evaluation of the neuroprotective potential of Indigofera sessiliflora through in-silico studies and behavioral tests in scopolamine-induced memory compromised rats. Saudi J. Biol. Sci. 2021, 28, 4384–4398. [Google Scholar] [CrossRef]
- Haider, S.; Naqvi, F.; Batool, Z.; Tabassum, S.; Perveen, T.; Saleem, S.; Haleem, D.J. Decreased hippocampal 5-HT and DA levels following sub-chronic exposure to noise stress: Impairment in both spatial and recognition memory in male rats. Sci. Pharm. 2012, 80, 1001–1011. [Google Scholar] [CrossRef]
- Bonaccorsi, J.; Cintoli, S.; Mastrogiacomo, R.; Baldanzi, S.; Braschi, C.; Pizzorusso, T.; Cenni, M.C.; Berardi, N. System consolidation of spatial memories in mice: Effects of enriched environment. Neural Plast. 2013, 2013, 956312. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shen, P.; He, T.; Chang, Y.; Shi, L.; Tao, S.; Li, X.; Xun, Q.; Guo, X.; Yu, Z.; et al. Noise induced hearing loss impairs spatial learning/memory and hippocampal neurogenesis in mice. Sci. Rep. 2016, 6, 20374. [Google Scholar] [CrossRef] [PubMed]
- Aleem, A.; Shahnaz, S.; Javaid, S.; Ashraf, W.; Rasool, M.F.; Ahmad, T.; Alotaibi, A.F.; Albeshri, K.S.; Alqahtani, F.; Imran, I. Chronically administered Agave americana var. marginata extract ameliorates diabetes mellitus, associated behavioral comorbidities and biochemical parameters in alloxan-induced diabetic rats. Saudi Pharm. J. 2022, 30, 1373–1386. [Google Scholar] [CrossRef]
- Qamar, M.; Akhtar, S.; Ismail, T.; Sestili, P.; Tawab, A.; Ahmed, N. Anticancer and anti-inflammatory perspectives of Pakistan’s indigenous berry Grewia asiatica Linn (Phalsa). J. Berry. Res. 2020, 10, 115–131. [Google Scholar] [CrossRef]
- Qamar, M.; Akhtar, S.; Ismail, T.; Yuan, Y.; Ahmad, N.; Tawab, A.; Ismail, A.; Barnard, R.T.; Cooper, M.A.; Blaskovich, M.A.; et al. Syzygium cumini (L.), Skeels fruit extracts: In vitro and in vivo anti-inflammatory properties. J. Ethnopharmacol. 2021, 271, 113805. [Google Scholar] [CrossRef] [PubMed]
- Wahid, M.; Saqib, F.; Ahmedah, H.T.; Gavris, C.M.; De Feo, V.; Hogea, M.; Moga, M.; Chicea, R. Cucumis sativus L. seeds ameliorate muscular spasm-induced gastrointestinal and respiratory disorders by simultaneously inhibiting calcium mediated signaling pathway. Pharmaceuticals 2021, 14, 1197. [Google Scholar] [CrossRef]
- Ferrari, I.V. Open access in silico tools to predict the ADMET profiling and PASS (Prediction of Activity Spectra for Substances of Bioactive compounds of Garlic (Allium sativum L.). BioRxiv 2021. [Google Scholar] [CrossRef]
- Anza, M.; Endale, M.; Cardona, L.; Cortes, D.; Eswaramoorthy, R.; Zueco, J.; Rico, H.; Trelis, M.; Abarca, B. Antimicrobial activity, in silico molecular docking, ADMET and DFT analysis of secondary metabolites from roots of three Ethiopian medicinal plants. Adv. Appl. Bioinform. Chem. 2021, 14, 117–132. [Google Scholar] [CrossRef]
- Sirous, H.; Chemi, G.; Campiani, G.; Brogi, S. An integrated in silico screening strategy for identifying promising disruptors of p53-MDM2 interaction. Comput. Biol. Chem. 2019, 83, 107105. [Google Scholar] [CrossRef]
- Krushna, G.S.; Shivaranjani, V.L.; Umamaheswari, J.; Srinivasulu, C.; Hussain, S.A.; Kareem, M.A.; Reddy, V.D.; Ali, D.; Lokhande, K.B.; Swamy, K.V.; et al. In vivo and molecular docking studies using whole extract and phytocompounds of Aegle marmelos fruit protective effects against isoproterenol-induced myocardial infarction in rats. Biomed. Pharmacother. 2017, 91, 880–889. [Google Scholar] [CrossRef]
- Paul, R.; Borah, A. Global loss of acetylcholinesterase activity with mitochondrial complexes inhibition and inflammation in brain of hypercholesterolemic mice. Sci. Rep. 2017, 7, 17922. [Google Scholar] [CrossRef]
- Wahid, M.; Saqib, F. Scientific basis for medicinal use of Citrullus lanatus (Thunb.) in diarrhea and asthma: In vitro, in vivo and in silico studies. Phytomedicine 2022, 98, 153978. [Google Scholar] [CrossRef]
- Wen, J.; Zou, W.; Wang, R.; Liu, H.; Yang, Y.; Li, H.; Wei, S.; Li, R.; Cai, H.; Wang, J.; et al. Cardioprotective effects of Aconiti Lateralis, Radix Praeparata combined with Zingiberis Rhizoma on doxorubicin-induced chronic heart failure in rats and potential mechanisms. J. Ethnopharmacol. 2019, 238, 111880. [Google Scholar] [CrossRef]
- Pan, H.-T.; Xi, Z.-Q.; Wei, X.-Q.; Wang, K. A network pharmacology approach to predict potential targets and mechanisms of “Ramulus Cinnamomi (cassiae)–Paeonia lactiflora” herb pair in the treatment of chronic pain with comorbid anxiety and depression. Ann. Med. 2022, 54, 413–425. [Google Scholar] [CrossRef]
- Xiong, W.-C.; Wu, H.-Z.; Xiong, Y.-Y.; Liu, B.; Xie, Z.-T.; Wu, S.-T.; Yao, Y.-F.; Yang, Y.-F. Network pharmacology-based research of active components of Albiziae flos and mechanisms of its antidepressant effect. Curr. Med. Sci. 2020, 40, 123–129. [Google Scholar] [CrossRef]
- Raafat, K. Identification of phytochemicals from North African plants for treating Alzheimer’s diseases and of their molecular targets by in silico network pharmacology approach. J. Tradit. Complement. Med. 2021, 11, 268–278. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Xiao, P.-T.; Liu, S.-Y.; Kuang, Y.-J.; Jiang, Z.-M.; Lin, Y.; Xie, Z.-S.; Liu, E.-H. Network pharmacology analysis and experimental validation to explore the mechanism of sea buckthorn flavonoids on hyperlipidemia. J. Ethnopharmacol. 2021, 264, 113380. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qin, X.-M.; Tian, J.-S.; Gao, X.-X.; Du, G.-H.; Zhou, Y.-Z. Integrated network pharmacology and metabolomics to dissect the combination mechanisms of Bupleurum chinense DC-Paeonia lactiflora Pall herb pair for treating depression. J. Ethnopharmacol. 2021, 264, 113281. [Google Scholar] [CrossRef] [PubMed]
- Wahid, M.; Saqib, F.; Akhtar, S.; Ali, A.; Wilairatana, P.; Mubarak, M.S. Possible mechanisms underlying the antispasmodic, bronchodilator, and antidiarrheal activities of polarity–Based extracts of Cucumis sativus L. Seeds in in silico, in vitro, and in vivo studies. Pharmaceuticals 2022, 15, 641. [Google Scholar] [CrossRef]
- Silvestre, S.M.; Almeida, P.J.; El-Shishtawy, R. Natural products as a source for new leads in gout treatment. Evid.-Based Complement. Altern. Med. 2020, 2020, 8274975. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.K.D.N.; de Almeida, V.S.; de Araujo, D.R.C.; Harand, W.; de Araújo Soares, A.K.; Moreira, L.R.; de Lorena, V.M.B.; Magalhães, L.P.M.; Ximenes, R.M.; de Sena, K.X.D.F.R.; et al. Evaluation of cytotoxic, immunomodulatory and antibacterial activities of aqueous extract from leaves of Conocarpus erectus Linnaeus (Combretaceae). J. Pharm. Pharmacol. 2018, 70, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Naseer, S.; Iqbal, J.; Naseer, A.; Kanwal, S.; Hussain, I.; Tan, Y.; Aguilar-Marcelino, L.; Cossio-Bayugar, R.; Zajac, Z.; Bin Jardan, Y.A.; et al. Deciphering chemical profiling, pharmacological responses and potential bioactive constituents of Saussurea lappa Decne. Extracts through in vitro approaches. Saudi J. Biol. Sci. 2022, 29, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Farkas, O.; Jakus, J.; Héberger, K. Quantitative structure–Antioxidant activity relationships of flavonoid compounds. Molecules 2004, 9, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Ovais, M.; Ullah, I.; Ahmed, J.; Shahid, M. Flavonoids as Prospective Neuroprotectants and Their Therapeutic Propensity in Aging Associated Neurological Disorders. Front. Aging Neurosci. 2019, 11, 155. [Google Scholar] [CrossRef]
- Park, S.-J.; Kim, S.-D.; Kwag, E.-B.; Park, J.H.; Yoo, H.-S. Acute and subchronic toxicological evaluation of the herbal product HAD-B1 in rats. Evid. Based Complement. Altern. Med. 2021, 2021, 9970822. [Google Scholar] [CrossRef]
- Sireeratawong, S.; Lertprasertsuke, N.; Srisawat, U.; Thuppia, A.; Ngamjariyawat, A.; Suwanlikhid, N.; Jaijoy, K. Acute and subchronic toxicity study of the water extract from Tiliacora triandra (Colebr.) Diels in rats. Songklanakarin J. Sci. Technol. 2008, 30, 611–619. [Google Scholar]
- Gibson, G.G.; Skett, P. Introduction to Drug Metabolism, 3rd ed.; Nelson Thornes Ltd.: Hampshire, UK, 2001; p. 256. [Google Scholar]
- Ugwah-Oguejiofor, C.J.; Okoli, C.O.; Ugwah, M.O.; Umaru, M.L.; Ogbulie, C.S.; Mshelia, H.E.; Umar, M.; Njan, A.A. Acute and sub-acute toxicity of aqueous extract of aerial parts of Caralluma dalzielii N. E. Brown in mice and rats. Heliyon 2019, 5, e01179. [Google Scholar] [CrossRef]
- Kifayatullah, M.; Mustafa, M.S.; Sengupta, P.; Sarker, M.M.R.; Das, A.; Das, S.K. Evaluation of the acute and sub-acute toxicity of the ethanolic extract of Pericampylus glaucus (Lam.) Merr. in BALB/c mice. J. Acute Dis. 2015, 4, 309–315. [Google Scholar] [CrossRef]
- Nfozon, J.; Tume, C.; Kdjo, N.; Boyom, F.; Leonard, S.; Dzoyem, J.; Metinous, S. Acute and sub-chronic toxicity evaluation of Triplotaxis stellulifera (Benth.) hutch and Crasssocephalum bougheyanum CD Adams methanol extract on mice. Biochem. Anal. Biochem. 2019, 8, 1–10. [Google Scholar]
- Benrahou, K.; Mrabti, H.N.; Assaggaf, H.M.; Mortada, S.; Salhi, N.; Rouas, L.; El Bacha, R.; Dami, A.; Masrar, A.; Alshahrani, M.M.; et al. Acute and subacute toxicity studies of Erodium guttatum extracts by oral administration in rodents. Toxins 2022, 14, 735. [Google Scholar] [CrossRef]
- Kharchoufa, L.; Bouhrim, M.; Bencheikh, N.; El Assri, S.; Amirou, A.; Yamani, A.; Choukri, M.; Mekhfi, H.; Elachouri, M. Acute and subacute toxicity studies of the aqueous extract from Haloxylon scoparium Pomel (Hammada scoparia (Pomel)) by oral administration in rodents. Biomed. Res. Int. 2020, 2020, 4020647. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, M.; Ostad, S.N.; Asemi, Z.; Mahboubi, M.; Hejazi, S.; Sharafati-Chaleshtori, R.; Rashidi, A.; Akbari, H.; Sharifi, N. Sub-chronic oral toxicity of Cuminum cyminum L.’s essential oil in female Wistar rats. Regul. Toxicol. Pharmacol. 2017, 88, 138–143. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, M.; Song, J.; Shi, Y.; Qin, X.; Gao, Z.; Lv, Y.; Du, G. The cardioprotective effects of the new crystal form of puerarin in isoproterenol-induced myocardial ischemia rats based on metabolomics. Sci. Rep. 2020, 10, 17787. [Google Scholar] [CrossRef]
- Paulino, E.T.; Ferreira, A.K.B.; da Silva, J.C.G.; Costa, C.D.F.; Smaniotto, S.; de Araujo-Junior, J.X.; Júnior, E.F.S.; Bortoluzzi, J.H.; Ribeiro, Ê.A.N. Cardioprotective effects induced by hydroalcoholic extract of leaves of Alpinia zerumbet on myocardial infarction in rats. J. Ethnopharmacol. 2019, 242, 112037. [Google Scholar] [CrossRef]
- Xie, J.; Cha, S.K.; An, S.W.; Kuro-o, M.; Birnbaumer, L.; Huang, C.L. Cardioprotection by Klotho through downregulation of TRPC6 channels in the mouse heart. Nat. Commun. 2012, 3, 1238. [Google Scholar] [CrossRef]
- Elhadidy, M.G.; Elalfy, M.M.; Saad, L. Possible Effect of Abscisic acid on Nitric Oxide in Isoproterenol-Induced Myocardial Infarction in Rats. Bull. Egypt. Soc. Physiol. Sci. 2020, 40, 17–30. [Google Scholar] [CrossRef]
- Patel, V.; Upaganlawar, A.; Zalawadia, R.; Balaraman, R. Cardioprotective effect of melatonin against isoproterenol induced myocardial infarction in rats: A biochemical, electrocardiographic and histoarchitectural evaluation. Eur. J. Pharmacol. 2010, 644, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; McGaffin, K.R.; Pastor-Soler, N.M.; Ahmad, F. SGLT1 is a novel cardiac glucose transporter that is perturbed in disease states. Cardiovasc. Res. 2009, 84, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Feriani, A.; Tir, M.; Aldahmash, W.; Mnafgui, K.; Hichem, A.; Gómez-Caravaca, A.M.; del Mar Contreras, M.; Taamalli, A.; Alwasel, S.; Segura-Carretero, A.; et al. In vivo evaluation and molecular docking studies of Schinus molle L. fruit extract protective effect against isoproterenol-induced infarction in rats. Environ. Sci. Pollut. Res. 2022, 29, 80910–80925. [Google Scholar] [CrossRef]
- Aboubakr, M.; Elsayd, F.; Soliman, A.; Fadl, S.E.; El-Shafey, A.; Abdelhiee, E.Y. L-Carnitine and vitamin E ameliorate cardiotoxicity induced by tilmicosin in rats. Environ. Sci. Pollut. Res. 2020, 27, 23026–23034. [Google Scholar] [CrossRef]
- Shi, L.; Du, X.; Zuo, B.; Hu, J.; Cao, W. Qige Huxin Formula Attenuates Isoprenaline-Induced Cardiac Fibrosis in Mice via Modulating Gut Microbiota and Protecting Intestinal Integrity. Evid. Based Complement. Altern. Med. 2022, 2022, 2894659. [Google Scholar] [CrossRef] [PubMed]
- Ismail, D.I.; ShamsEldeen, A.M.; Rashed, L.A.; Shama, A.A.E.D.; Ashour, S.S.; Aboulkhair, A.G. Cardioprotective Potential of Zinc and Vitamin E Against Isoprenaline-Induced Myocardial Infarction in Albino Rats by Targeting Autophagy: A Histological and Biochemical Study. Egypt. J. Histol. 2021, 44, 450–464. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, R.; Zhang, J.; Liang, T.; Guo, J.; Sun, T.; Fu, X.; Wang, L.; Zhang, L. Pinoresinol diglucoside (PDG) attenuates cardiac hypertrophy via AKT/mTOR/NF-κB signaling in pressure overload-induced rats. J. Ethnopharmacol. 2021, 272, 113920. [Google Scholar] [CrossRef]
- Akter, N.; Chowdhury, F.I.; Selim, S.; Nayan, S.I.; Khan, F.; Subhan, N.; Hossain, H.; Rahman, M.M.; Haque, M.A.; Alam, M.A. Polyphenolics in ramontchi protect cardiac tissues via suppressing isoprenaline-induced oxidative stress and inflammatory responses in Long-Evans rats. J. Funct. Foods 2020, 75, 104250. [Google Scholar] [CrossRef]
- Lubos, E.; Handy, D.E.; Loscalzo, J. Role of oxidative stress and nitric oxide in atherothrombosis. Front. Biosci. 2008, 13, 5323–5344. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, L.; Lu, S. Evaluation of antioxidant and immunity activities of quercetin in isoproterenol-treated rats. Molecules 2012, 17, 4281–4291. [Google Scholar] [CrossRef] [PubMed]
- Avci, E.; Dolapoglu, A.; Akgun, D.E. Role of Cholesterol as a Risk Factor in Cardiovascular Diseases. In Cholesterol—Good, Bad, and the Heart, 1st ed.; Nagpanl, M.L., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Ramirez, A.; Hu, P.P. Low High-Density Lipoprotein and Risk of Myocardial Infarction. Clin. Med. Insights Cardiol. 2015, 9, 113–117. [Google Scholar] [CrossRef]
- Venkadeswaran, K.; Thomas, P.A.; Geraldine, P. An experimental evaluation of the anti-atherogenic potential of the plant, Piper betle, and its active constitutent, eugenol, in rats fed an atherogenic diet. Biomed. Pharmacother. 2016, 80, 276–288. [Google Scholar] [CrossRef]
- Shaito, A.; Thuan, D.T.B.; Phu, H.T.; Nguyen, T.H.D.; Hasan, H.; Halabi, S.; Abdelhady, S.; Nasrallah, G.K.; Eid, A.H.; Pintus, G. Herbal Medicine for Cardiovascular Diseases: Efficacy, Mechanisms, and Safety. Front. Pharmacol. 2020, 11, 422. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, X.; Hu, M.; Zhang, D.; Yuan, S.; Li, P.; Feng, L. Medicinal and edible plants in the treatment of dyslipidemia: Advances and prospects. Chin. Med. 2022, 17, 113. [Google Scholar] [CrossRef]
- Moris, D.; Spartalis, M.; Spartalis, E.; Karachaliou, G.-S.; Karaolanis, G.I.; Tsourouflis, G.; Tsilimigras, D.I.; Tzatzaki, E.; Theocharis, S. The role of reactive oxygen species in the pathophysiology of cardiovascular diseases and the clinical significance of myocardial redox. Ann. Transl. Med. 2017, 5, 326. [Google Scholar] [CrossRef]
- Qi, C.; Liu, X.; Xiong, T.; Wang, D. Tempol prevents isoprenaline-induced takotsubo syndrome via the reactive oxygen species/mitochondrial/anti-apoptosis/p38 MAPK pathway. Eur. J. Pharmacol. 2020, 886, 173439. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.E.; Hare, J.M. Xanthine oxidoreductase and cardiovascular disease: Molecular mechanisms and pathophysiological implications. J. Physiol. 2004, 555, 589–606. [Google Scholar] [CrossRef]
- Shanmugam, P.; Valente, A.J.; Prabhu, S.D.; Venkatesan, B.; Yoshida, T.; Delafontaine, P.; Chandrasekar, B. Angiotensin-II type 1 receptor and NOX2 mediate TCF/LEF and CREB dependent WISP1 induction and cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol. 2011, 50, 928–938. [Google Scholar] [CrossRef]
- Li, S.; Liu, H.; Lin, Z.; Li, Z.; Chen, Y.; Chen, B.; Huang, L.; Lin, X.; Yao, H. Isoorientin attenuates doxorubicin-induced cardiac injury via the activation of MAPK, Akt, and Caspase-dependent signaling pathways. Phytomedicine 2022, 101, 154105. [Google Scholar] [CrossRef] [PubMed]
- Zholobenko, A.; Modriansky, M. Silymarin and its constituents in cardiac preconditioning. Fitoterapia 2014, 97, 122–132. [Google Scholar] [CrossRef]
- Taleb, A.; Ahmad, K.A.; Ihsan, A.U.; Qu, J.; Lin, N.; Hezam, K.; Koju, N.; Hui, L.; Qilong, D. Antioxidant effects and mechanism of silymarin in oxidative stress induced cardiovascular diseases. Biomed. Pharmacother. 2018, 102, 689–698. [Google Scholar] [CrossRef]
- Liu, X.; Hu, M.; Ye, C.; Liao, L.; Ding, C.; Sun, L.; Liang, J.; Chen, Y. Isosilybin regulates lipogenesis and fatty acid oxidation via the AMPK/SREBP-1c/PPARα pathway. Chem. Biol. Interact. 2022, 368, 110250. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Jiao, S.; Qin, M.; Hu, W.; Yi, B.; Liu, D. Vanillic acid alleviates acute myocardial hypoxia/reoxygenation injury by inhibiting oxidative stress. Oxidative Med. Cell. Longev. 2020, 2020, 8348035. [Google Scholar] [CrossRef]
- Stanely Mainzen Prince, P.; Rajakumar, S.; Dhanasekar, K. Protective effects of vanillic acid on electrocardiogram, lipid peroxidation, antioxidants, proinflammatory markers and histopathology in isoproterenol induced cardiotoxic rats. Eur. J. Pharmacol. 2011, 668, 233–240. [Google Scholar] [CrossRef]
- Wang, S.; Sun, X.; Wang, Z.; Zhou, S.; Su, S.; Nan, X.; Lu, D.; Li, Z. Vanillic acid attenuates monocrotaline-induced pulmonary arterial hypertension by enhancing NO signaling pathways. Nat. Prod. Commun. 2022, 17, 1934578X221128411. [Google Scholar] [CrossRef]
- Lim, J.; Kweon, K.; Kim, H.-W.; Cho, S.W.; Park, J.; Sim, C.S. Negative impact of noise and noise sensitivity on mental health in childhood. Noise Health 2018, 20, 199–211. [Google Scholar]
- Kent, M.; Bardi, M.; Hazelgrove, A.; Sewell, K.; Kirk, E.; Thompson, B.; Trexler, K.; Terhune-Cotter, B.; Lambert, K. Profiling coping strategies in male and female rats: Potential neurobehavioral markers of increased resilience to depressive symptoms. Horm. Behav. 2017, 95, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Mineur, Y.S.; Obayemi, A.; Wigestrand, M.B.; Fote, G.M.; Calarco, C.A.; Li, A.M.; Picciotto, M.R. Cholinergic signaling in the hippocampus regulates social stress resilience and anxiety- and depression-like behavior. Proc. Natl. Acad. Sci. USA 2013, 110, 3573–3578. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Zakaria, R.; AbdAziz, C.; Othman, Z.; Al-Rahbi, B. Tualang honey improves memory performance and decreases depressive-like behavior in rats exposed to loud noise stress. Noise Health 2015, 17, 83–89. [Google Scholar] [CrossRef]
- Sturman, O.; Germain, P.-L.; Bohacek, J. Exploratory rearing: A context- and stress-sensitive behavior recorded in the open-field test. Stress 2018, 21, 443–452. [Google Scholar] [CrossRef]
- Ma, L. Depression, anxiety, and apathy in mild cognitive impairment: Current perspectives. Front. Aging Neurosci. 2020, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Samson, J.; Sheeladevi, R.; Ravindran, R. Oxidative stress in brain and antioxidant activity of Ocimum sanctum in noise exposure. Neurotoxicology 2007, 28, 679–685. [Google Scholar] [CrossRef]
- Manikandan, S.; Devi, R.S. Antioxidant property of α-Asarone against noise-stress-induced changes in different regions of rat brain. Pharmacol. Res. 2005, 52, 467–474. [Google Scholar] [CrossRef]
- Endo, T.; Nakagawa, T.; Iguchi, F.; Kita, T.; Okano, T.; Sha, S.-H.; Schacht, J.; Shiga, A.; Kim, T.S.; Ito, J. Elevation of superoxide dismutase increases acoustic trauma from noise exposure. Free Radic. Biol. Med. 2005, 38, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Flores, N.P.; Bona, N.P.; Luduvico, K.P.; de Souza Cardoso, J.; Soares, M.S.P.; Gamaro, G.D.; Spanevello, R.M.; Lencina, C.L.; Gazal, M.; Stefanello, F.M. Eugenia uniflora fruit extract exerts neuroprotective effect on chronic unpredictable stress-induced behavioral and neurochemical changes. J. Food Biochem. 2020, 44, e13442. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, S.; Padma, M.K.; Srikumar, R.; Jeya Parthasarathy, N.; Muthuvel, A.; Sheela Devi, R. Effects of chronic noise stress on spatial memory of rats in relation to neuronal dendritic alteration and free radical-imbalance in hippocampus and medial prefrontal cortex. Neurosci. Lett. 2006, 399, 17–22. [Google Scholar] [CrossRef]
- Wankhar, D.; Sheela Devi, R.; Ashok, I. Emblica officinalis outcome on noise stress induced behavioural changes in Wistar albino rats. Biomed. Prev. Nutr. 2014, 4, 219–224. [Google Scholar] [CrossRef]
- Samad, N.; Muneer, A.; Ullah, N.; Zaman, A.; Ayaz, M.M.; Ahmad, I. Banana fruit pulp and peel involved in antianxiety and antidepressant effects while invigorate memory performance in male mice: Possible role of potential antioxidants. Pak. J. Pharm. Sci. 2017, 30, 989–995. [Google Scholar]
- Nandini, H.S.; Naik, P.R. Antidiabetic, antihyperlipidemic and antioxidant effect of Vincamine, in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2019, 843, 233–239. [Google Scholar] [CrossRef]
- Vas, Á.; Gulyás, B. Eburnamine derivatives and the brain. Med. Res. Rev. 2005, 25, 737–757. [Google Scholar] [CrossRef]
- Tan, X.; Liang, Z.; Li, Y.; Zhi, Y.; Yi, L.; Bai, S.; Forest, K.H.; Nichols, R.A.; Dong, Y.; Li, Q.X. Isoorientin, a GSK-3β inhibitor, rescues synaptic dysfunction, spatial memory deficits and attenuates pathological progression in APP/PS1 model mice. Behav. Brain Res. 2021, 398, 112968. [Google Scholar] [CrossRef]
- Ko, Y.H.; Kwon, S.H.; Lee, S.Y.; Jang, C.G. Isoorientin improves scopolamine-induced cognitive impairments by restoring the cholinergic system, antioxidant defense, and p-CREB/BDNF signaling in the hippocampus and frontal cortex. Arch. Pharm. Res. 2019, 42, 722–731. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, A.K.; Kumar, R.; Jamieson, S.; Pandey, A.K.; Bishayee, A. Neuroprotective potential of Ellagic Acid: A Critical Review. Adv. Nutr. 2021, 12, 1211–1238. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Quispe, C.; Hossain, R.; Islam, M.T.; Al-Harrasi, A.; Al-Rawahi, A.; Martorell, M.; Mamurova, A.; Seilkhan, A.; Altybaeva, N.; et al. Neuropharmacological Effects of Quercetin: A Literature-Based Review. Front. Pharmacol. 2021, 12, 665031. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Sun, T.; Yang, L.; Liu, A.; Liu, Q.Q.; Tian, Q.Q.; Wang, Y.; Zhao, M.G.; Yang, Q. Scopoletin ameliorates anxiety-like behaviors in complete Freund’s adjuvant-induced mouse model. Mol. Brain 2020, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhao, W.; Ge, C.; Li, X.; Sun, Z. Scopoletin attenuates intracerebral hemorrhage-induced brain injury and improves neurological performance in rats. Neuroimmunomodulation 2021, 28, 74–81. [Google Scholar] [CrossRef]
- Hornick, A.; Lieb, A.; Vo, N.P.; Rollinger, J.M.; Stuppner, H.; Prast, H. The coumarin scopoletin potentiates acetylcholine release from synaptosomes, amplifies hippocampal long-term potentiation and ameliorates anticholinergic- and age-impaired memory. Neuroscience 2011, 197, 280–292. [Google Scholar] [CrossRef]
- Ali, S.K.; Hamed, A.R.; Soltan, M.M.; El-Halawany, A.M.; Hegazy, U.M.; Hussein, A.A. Kinetics and molecular docking of Vasicine from Adhatoda vasica: An acetylcholinesterase inhibitor for Alzheimer’s disease. S. Afr. J. Bot. 2016, 104, 118–124. [Google Scholar] [CrossRef]
- Liu, W.; Shi, X.; Yang, Y.; Cheng, X.; Liu, Q.; Han, H.; Yang, B.; He, C.; Wang, Y.; Jiang, B.; et al. In vitro and in vivo metabolism and inhibitory activities of Vasicine, a potent acetylcholinesterase and butyrylcholinesterase inhibitor. PLoS ONE 2015, 10, e0122366. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.H.; Nguyen, N.P.K.; Tran, K.N.; Shin, H.M.; Yang, I.J. Network pharmacology and experimental validation to investigate the antidepressant potential of Atractylodes lancea (Thunb.) DC. Life 2022, 12, 1925. [Google Scholar] [CrossRef]
- Saadullah, M.; Asif, M.; Ch, A.B.; Yaseen, H.S.; Uzair, M.; Afzal, K. Isolation, characterization and preliminary cytotoxic and antifungal evaluations of novel Lancifoliate isolated from methanol extract of Conocarpus lancifolius. Anti-Cancer Agents Med. Chem. 2020, 20, 1664–1672. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TPC | TFC | DPPH Assay | TAC | TRP | AChE Assay |
|---|---|---|---|---|---|---|
| (µg GAE/mg) | (µg QE/mg) | IC50 (µg/mL) | (µg AAE/mg) | (µg GAE/mg) | IC50 (µg/mL) | |
| 250 µg/mL | 250 µg/mL | |||||
| CLAE | 67.70 ± 0.15 | 47.54 ± 0.45 | 16.66 ± 0.42 | 77.33 ± 0.41 | 79.11 ± 0.67 | 110.13 ± 1.71 |
| - | - | 8.67 ± 1.83 a | - | - | 6.48 ± 1.29 b |
| Parameters | Control | Single-Dose Acute | Repeated-Dose Subacute | |
|---|---|---|---|---|
| Observed | Toxicity | Toxicity | ||
| 2000 mg/kg | 400 mg/kg | 800 mg/kg | ||
| Final body weight (g) | 33.84 ± 2.68 | 34.76 ± 3.43 | 32.42 ± 1.73 | 30.41 ± 3.20 |
| Organ weight | ||||
| Heart (g) | 0.269 ± 0.021 | 0.272 ± 0.015 | 0.261 ± 0.007 | 0.265 ± 0.030 |
| Kidney (g) | 0.342 ± 0.039 | 0.354 ± 0.023 | 0.341 ± 0.037 | 0.339 ± 0.015 |
| Liver (g) | 2.844 ± 0.224 | 2.821 ± 0.269 | 2.711 ± 0.192 | 2.674 ± 0.167 |
| Parameters | Control | Repeated-Dose Subacute Toxicity CLAE | |
| 400 mg/kg | 800 mg/kg | ||
| Biochemicals | |||
| Albumin (g/dL) | 1.732 ± 0.959 | 2.128 ± 1.326 | 2.792 ± 0.755 |
| Albumin/Globulin ratio | 2.112 ± 0.568 | 2.226 ± 0.347 | 2.718 ± 0.283 |
| Total Protein (g/dL) | 3.852 ± 1.927 | 4.412 ± 1.103 | 5.458 ± 0.362 |
| Alkaline Phosphatase (U/L) | 287.5 ± 18.97 | 303.6 ± 28.75 | 334.4 ± 41.91 |
| Alanine Transaminase (U/L) | 30.67 ± 8.491 | 39.36 ± 7.826 | 44.05 ± 9.680 |
| Aspartate Transaminase (U/L) | 98.76 ± 28.39 | 123.3 ± 19.19 | 141.2 ± 24.03 |
| Uric Acid (mg/dL) | 0.800 ± 0.100 | 1.028 ± 0.279 | 1.054 ± 0.365 |
| Creatinine (mg/dL) | 0.976 ± 0.392 | 1.288 ± 0.352 | 1.724 ± 0.630 |
| Total bilirubin (mg/dL) | 0.216 ± 0.055 | 0.232 ± 0.049 | 0.284 ± 0.102 |
| Total Cholesterol (mg/dL) | 79.80 ± 15.40 | 80.26 ± 17.91 | 83.41 ± 11.82 |
| Triglycerides (mg/dL) | 122.3 ± 27.46 | 132.4 ± 11.85 | 162.2 ± 39.85 |
| Sodium (mmol/L) | 110.7 ± 33.13 | 104.5 ± 23.16 | 151.2 ± 25.31 |
| Potassium (mmol/L) | 6.380 ± 1.108 | 7.176 ± 0.367 | 9.082 ± 0.128 |
| Chloride (mmol/L) | 96.00 ± 11.77 | 113.3 ± 20.01 | 127 ± 26.93 |
| Hematological | |||
| RBCs (106/µL) | 8.660 ± 2.195 | 9.640 ± 0.555 | 11.32 ± 0.978 |
| Hemoglobin (g/dL) | 13.24 ± 1.414 | 15.40 ± 1.517 | 20.38 ± 5.364 |
| Hematocrit (%) | 45.69 ± 2.355 | 48.26 ± 6.017 | 57.00 ± 10 |
| Mean corpuscular volume (f/L) | 54.20 ± 10.50 | 59.83 ± 12.79 | 71.19 ± 18.36 |
| Mean corpuscular hemoglobin (pg) | 17.80 ± 1.834 | 18.58 ± 2.924 | 22.61 ± 4.479 |
| MCHC (%) | 31.17 ± 6.058 | 32 ± 3.245 | 35.82 ± 2.305 |
| WBCs (105/µL) | 4.643 ± 0.750 | 5.622 ± 0.809 | 5.671 ± 0.398 |
| Neutrophils (%) | 38.58 ± 3.846 | 42.80 ± 8.956 | 47.22 ± 5.516 |
| Lymphocytes (%) | 49.62 ± 6.243 | 52.34 ± 3.553 | 60.95 ± 10.71 |
| Eosinophils (%) | 1.060 ± 0.357 | 0.998 ± 0.416 | 1.080 ± 0.266 |
| Platelets (105/µL) | 5.560 ± 0.931 | 6.977 ± 0.584 | 7.368 ± 1.227 |
| Comp. No. | R.T. (min) | Analysis Mode | m/z | Product Ions (m/z) | Accurate Mass | Chemical Class | Proposed Compounds | References |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.45 | [M − H]− | 130.67 | 129, 113.92, 101.08, 99, 85, | 131.09463 | Amino acid | L-Leucine | MassBank |
| 83.08, 69 | KO001262 | |||||||
| 2 | 0.59 | [M − H]− | 133 | 115, 105, 97, 89, 71, 59.08 | 134.02152 | Carboxylic | Malic acid | MassBank |
| acid | KO001305 | |||||||
| 3 | 0.74 | [M − H]− | 166.92 | 152, 139.08, 131, 123, 108, | 168.04230 | Phenolic | Vanillic acid | MassBank |
| 92.92, 73.08, 59.17 | acid | ML005851 | ||||||
| 4 | 0.81 | [M − H]− | 169 | 155.67, 141.08, 131.08, 125, | 170.02152 | Phenolic | Gallic acid | MassBank |
| 113, 92.83, 81.08, 68.75 | acid | KO000889 | ||||||
| 5 | 1.73 | [M − H]− | 196.75 | 182.08, 169.08, 161, 152.92, | 198.05282 | Phenolic | Syringic acid | MassBank |
| 124.92, 109, 96.92 | acid | KO001814 | ||||||
| 6 | 2.45 | [M − H]− | 207.08 | 192, 171, 169, 163, 149.08, | 208.213 | Phenolic | 3,4-Dimethoxy- | MassBank |
| 132.92, 121, 89.08, 71.25 | acid | cinnamic acid | PR306017 | |||||
| 7 | 3.19 | [M − H]− | 275 | 260.08, 257, 239.08, 229.08, | 276.09977 | Coumarin | Capensine | MassBank |
| 201, 165.08, 147.17, 113 | BML00229 | |||||||
| 8 | 3.25 | [M − H]− | 287.08 | 268.92, 253.83, 250.92, 249, | 288.06339 | Flavonoid | Fustin | MassBank |
| 198.92, 154.92, 135, 126.83 | BS003856 | |||||||
| 9 | 3.74 | [M − H]− | 297.17 | 281.08, 279.17, 253.25, | 298.33890 | Lignan | Enterolactone | MassBank |
| 209.25, 197.08, 183, 153.25 | PR308012 | |||||||
| 10 | 3.83 | [M − H]− | 301.03 | 284, 257.08, 229, 201, 125, | 302.04265 | Flavonoid | Quercetin | MassBank |
| 117 | PB002411 | |||||||
| 11 | 3.89 | [M − H]− | 305 | 273.17, 244.83, 230.50, | 306.07395 | Catechin | Epigallocatechin | MassBank |
| 225.08, 207.17, 181.08, 103 | BS003952 | |||||||
| 12 | 4.19 | [M − H]− | 349.08 | 331, 313, 269.33, 241, | 350.20932 | Diterpene | Andrographolide | MassBank |
| 150.92, 138.92, 124.75 | BML80746 | |||||||
| 13 | 4.24 | [M − H]− | 351 | 333, 314.92, 306.92, 276.83, | 352.434 | Alkaloid | 14,15-Dehydro-16- | MassBank |
| 224.92, 222.83, 169, 152.67 | epi-vincamine | PR304643 | ||||||
| 14 | 4.55 | [M − H]− | 380.75 | 362, 299.92, 255.83, 129.58, | 381.17875 | Alkaloid | Otosenine | MassBank |
| 126.92 | BML81861 | |||||||
| 15 | 4.97 | [M − H]− | 383.17 | 365, 347, 339.25, 306.92, | 384.476 | Alkaloid | Voacristine | MassBank |
| 248.92, 221.92, 176.75 | PR304465 | |||||||
| 16 | 5.24 | [M − H]− | 385 | 367, 348.92, 327.25, 287.17, | 386.353 | Glycoside | 1-O- | MassBank |
| 237.08, 190.92, 154.83 | Sinapoylglucose | PR309003 | ||||||
| 17 | 5.35 | [M − H]− | 389.25 | 370.83, 352.92, 350.92, | 390.0951 | Flavonoid | Scaposin | MassBank |
| 345.17, 315, 271, 241, 153 | BS003673 | |||||||
| 18 | 5.79 | [M − H]− | 417.33 | 398.92, 380.92, 373.08, | 418.354 | Flavonoid | Kaempferol-3-O- | MassBank |
| 318.83, 267.08, 241, 160.75 | glycoside | arabinoside | PR306890 | |||||
| 19 | 5.94 | [M − H]− | 421.25 | 403.08, 384.92, 382.92, | 422.17293 | Flavonoid | Isoangustone A | HMDB |
| 346.92, 327.17, 293.08 | HMDB0038905 | |||||||
| 20 | 6.08 | [M − H]− | 423.08 | 405, 386.92, 349.08, 343.08, | 424.18858 | Flavonoid | Gancaonin E | HMDB |
| 328.17, 302.83, 279, 222.67 | HMDB0038868 | |||||||
| 21 | 6.44 | [M − H]− | 433.42 | 396.92, 365.08, 355.92, 317, | 434.397 | Flavonoid | Naringenin-7-O- | MassBank |
| 269.42, 214.08, 182.08 | glucoside | PR309316 | ||||||
| 22 | 6.53 | [M − H]− | 447.17 | 428.92, 411, 372.83, 349, | 448.10056 | Flavonoid | Isoorientin | MassBank |
| 301, 296.92, 201.17, 182 | FIO00716 | |||||||
| 23 | 6.94 | [M − H]− | 481 | 463, 444.83, 442.83, 423, | 482.12137 | Flavonoid | Isosilybin A | MassBank |
| 392.92, 351.25, 331, 301 | FIO01031 | |||||||
| 24 | 7.66 | [M − H]− | 529.92 | 511, 493.83, 469.25, 453.33, | 531.24683 | Alkaloid | Harringtonine | MassBank |
| 449.08, 431.33, 403.25, | BML82381 | |||||||
| 325.08, 299.92, 231.92, 182 | ||||||||
| 25 | 0.40 | [M + H]+ | 116.08 | 107, 98, 84, 74.08, 70, 58.08 | 115.06333 | Amino acid | L-Proline | MassBank |
| KO003672 | ||||||||
| 26 | 0.66 | [M + H]+ | 123 | 106.33, 105.08, 94.83, 81.83, | 122.03677 | Carboxylic | Benzoic acid | HMDB |
| 67.67, 52.08 | acid | HMDB0001870 | ||||||
| 27 | 0.81 | [M + H]+ | 130.25 | 114.17, 112.08, 98.08, 88.08, | 129.07897 | Amino acid | Cycloleucine | HMDB |
| 74.08, 70.08, 57 | HMDB0062225 | |||||||
| 28 | 1.00 | [M + H]+ | 138 | 133.50, 121, 118.08, 96.83, | 137.04768 | Benzoic acid | 4-Aminobenzoic | MassBank |
| 94, 78.92, 70, 57 | derivative | acid | KO002120 | |||||
| 29 | 1.31 | [M + H]+ | 148 | 139.08, 130.08, 116, 112.08, | 147.05316 | Amino acid | Glutamic acid | MassBank |
| 102.08, 100, 88.08, 79, 61 | PB000463 | |||||||
| 30 | 1.63 | [M + H]+ | 149 | 140, 131.08, 121, 111, 93, 79, | 148.161 | Cinnamaldehyde | p-Coumaraldehyde | MassBank |
| 58.83 | PR304040 | |||||||
| 31 | 1.80 | [M + H]+ | 156.83 | 142, 140.17, 134.92, 129.08, | 156.06874 | 3-Alkylindoles | 3-Indoleacetonitrile | HMDB |
| 111.08, 83.92, 79, 58.67 | HMDB0006524 | |||||||
| 32 | 1.89 | [M + H]+ | 163 | 145.08, 135, 133, 121.08, | 162.0317 | Coumarin | 4- | MassBank |
| 107, 95.08, 79, 70, 61 | derivative | Hydroxycoumarin | NA000184 | |||||
| 33 | 2.12 | [M + H]+ | 175.08 | 157.08, 153.08, 130.08, 116, | 174.01643 | Carboxylic | cis-Aconitic acid | HMDB |
| 112.08, 81.08, 72.08, 60.08 | acid | HMDB0000072 | ||||||
| 34 | 2.51 | [M + H]+ | 181.08 | 166.92, 163, 153.17, 149, | 180.0423 | Phenolic | Caffeic acid | MassBank |
| 135.08, 121, 112, 95, 69.08 | acid | RP016701 | ||||||
| 35 | 2.65 | [M + H]+ | 189.08 | 174.92, 171.08, 144.08, | 188.23 | Alkaloid | Vasicine | MassBank |
| 130.08, 114.92, 109, 95.08 | PR301208 | |||||||
| 36 | 2.94 | [M + H]+ | 193.08 | 184.08, 175.08, 168.92, 155, | 192.04226 | Coumarin | Scopoletin | MassBank |
| 148.92, 133, 120.08, 95.08 | PB002203 | |||||||
| 37 | 3.17 | [M + H]+ | 203.08 | 186.17, 185.08, 173.17, 161, | 202.213 | Alkaloid | Vasicinone | MassBank |
| 147, 143, 133.08, 103.08, 81 | PR301376 | |||||||
| 38 | 3.40 | [M + H]+ | 205.08 | 188.08, 177, 141 | 204.08988 | Amino acid | Tryptophan | MassBank |
| FIO00629 | ||||||||
| 39 | 3.48 | [M + H]+ | 215 | 200.08, 186.83, 174, 154.92, | 214.11061 | Alkaloid | Harmaline | MassBank |
| 131.08, 114, 110.08, 70.08 | KO003099 | |||||||
| 40 | 3.91 | [M + H]+ | 221.08 | 212.92, 203.08, 179, 177.08, | 220.07355 | Coumarin | 5,7-Dimethoxy-4- | MassBank |
| 161.08, 157, 121.17, 68.83 | methylcoumarin | BML01472 | ||||||
| 41 | 4.14 | [M + H]+ | 229.25 | 226.75, 211.17, 199.08, 193, | 228.247 | Stilbene | 3,4,5- | MassBank |
| 190.92, 169, 142.08, 86.17 | Trihydroxystilbene | PR302602 | ||||||
| 42 | 4.87 | [M + H]+ | 255.08 | 246.67, 237.08, 235, 227.17, | 254.0579 | Flavonoid | Chrysin | MassBank |
| 190.58, 183.08, 171, 101, 79 | LU080404 | |||||||
| 43 | 5.39 | [M + H]+ | 275.08 | 257.25, 247.08, 231.17, 197, | 274.0841 | Naphthopyra- | (R)-Semivioxanthin | MassBank |
| 170, 159.08, 102.08, 88.08 | none | HB003729 | ||||||
| 44 | 5.58 | [M + H]+ | 285 | 267.08, 249.17, 229.08, | 284.0685 | Flavonoid | Biochanin A | MassBank |
| 225.08, 206.92, 188.58, 173, | LU096003 | |||||||
| 45 | 5.74 | [M + H]+ | 303.08 | 285.17, 275.08, 247, 229.08, | 302.1991 | Tannin | Ellagic acid | MassBank |
| 199, 174.92, 137.17, 102.17 | CB000408 | |||||||
| 46 | 5.91 | [M + H]+ | 305.17 | 296.25, 268.75, 248.58, 245, | 304.2587 | Flavonoid | Taxifolin | MassBank |
| 192.50, 166.92, 135.33, 96 | CB000303 | |||||||
| 47 | 6.91 | [M + H]+ | 365.33 | 333.25, 308.83, 275.08, | 364.09468 | Lignan | Justicidin B | MassBank |
| 261.25, 184.42, 151.25 | BML01604 | |||||||
| 48 | 8.71 | [M + H]+ | 441.33 | 423.25, 418.08, 410.75, 399, | 440.162 | Lignan | Maruchantin E | MassBank |
| 381.08, 366.58, 339.25, | PB011901 | |||||||
| 313.17, 310.50, 250.75, | ||||||||
| 49 | 9.38 | [M + H]+ | 481.42 | 472.42, 449.25, 424.83, 393, | 480.29881 | Alkaloid | Emetine | MassBank |
| 408.83, 383.25, 367.08, 261 | CE000076 | |||||||
| 50 | 10.04 | [M + H]+ | 529.50 | 492.17, 483.08, 473.42, | 528.38148 | Triterpenoid | Pachymic acid | MassBank |
| 468.92, 464.92, 424.25, 417, | MSJ00299 | |||||||
| 51 | 12.30 | [M + H]+ | 819.75 | 801.58, 783.75, 771.08, | 818.687 | Flavonoid | Flavonol base + | MassBank |
| 752.58, 744.08, 735.75, | glycoside | 4O, 1MeO, O-Hex- | PR310938 | |||||
| 683, 668.33, 653.25, 632.58, | Hex, O-Hex | |||||||
| 52 | 13.31 | [M + H]+ | 839.75 | 790.17, 691.42, 665.33, | 838.941 | Triterpenoid | Licoricesaponin G2 | MassBank |
| 579.33, 526.42, 490.25, | saponin | PR310662 | ||||||
| 466.92, 448.92, 378.17 | ||||||||
| 53 | 13.75 | [M + H]+ | 867.21 | 849, 816, 701.42, 692.42, | 866.20581 | Tannin | Procyanidin C1 | HMDB |
| 611.50, 517, 509, 494, | HMDB0038370 | |||||||
| 477.25, 395.67, 284.08 |
| Name | Docking Score (XP- Glide) | Glide Energy | ΔG Binding | Log Ki (µMolar) | ΔG Coulomb | ΔG Covalent | ΔG Hbond | ΔG Lipo | ΔG Solv GB | ΔG vdW | Interactions of Residues and Their Ligands with Distance (Å) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hydrogen Bonds | Electrostatic/ Hydrophobic Bonds | |||||||||||
| HMG-CoA reductase (PBD ID: 2R4F) | ||||||||||||
| Isoorientin | −8.389 | −63.875 | −24.43 | −7.38 | −35.84 | 1.67 | −6.02 | −4.16 | 72.05 | −49.95 | Conventional Hydrogen | Electrostatic π-Cation; |
| Bond: Lys633 (2.58), | π-Donor Hydrogen | |||||||||||
| Lys633 (2.42), Glu700 | Bond: Lys633 (3.01) | |||||||||||
| (1.89), Glu700 (2.17), | π-Anion: Glu700 (4.49), | |||||||||||
| Glu610 (2.10), Gln648 | Glu700 (3.75), Glu700 | |||||||||||
| (1.95), Glu610 (2.04), | (3.47), | |||||||||||
| Glu610 (2.00), | π-Sigma: His635 (2.90), | |||||||||||
| Carbon Hydrogen | π-Alkyl: Lys633 (5.46), | |||||||||||
| Bond: His635 (2.64) | ||||||||||||
| Isosilybin A | −7.992 | −64.444 | −29.2 | −9.45 | −41.77 | 12.23 | −5.46 | −6.67 | 61.31 | −46.18 | Conventional Hydrogen | Electrostatic π-Cation: |
| Bond: Lys633 (2.72), | Lys633 (4.86), | |||||||||||
| Lys633 (2.43), Thr636 | π-Anion: Glu610 (4.75), | |||||||||||
| (1.88), Lys606 (2.60), | π-π Stacked: His635 | |||||||||||
| Thr636 (2.35), Glu610 | (4.40), | |||||||||||
| (2.21), Gln632 (1.63), | π-π T-shaped: His635 | |||||||||||
| Thr636 (1.74), | (4.88), | |||||||||||
| Carbon Hydrogen | π-Alkyl: Lys633 (4.98) | |||||||||||
| Bond: His635 (2.64), | ||||||||||||
| Lys633 (2.51), Glu700 | ||||||||||||
| (2.49), Leu634 (2.45), | ||||||||||||
| Leu634 (2.43), Glu610 | ||||||||||||
| (2.54) | ||||||||||||
| Epigallocatechin | −6.897 | −50.109 | −20.35 | −5.61 | −42.43 | 9.39 | −4.38 | −2.78 | 50.9 | −30.01 | Conventional Hydrogen | Electrostatic π-Cation: |
| Bond: Thr636 (2.08), | Lys633 (4.74), | |||||||||||
| Thr636 (2.41), Leu634 | π-Anion: Glu610 (4.97), | |||||||||||
| (2.19), Glu700 (1.80), | π-π T-shaped: His635 | |||||||||||
| Gln632 (1.76), Lys633 | (4.89), | |||||||||||
| (2.09), Glu700 (2.04), | π-Alkyl: Ala585 (4.44), | |||||||||||
| Glu700 (2.44), | Lys633 (4.84) | |||||||||||
| Carbon Hydrogen | ||||||||||||
| Bond: Glu610 (2.47) | ||||||||||||
| Fustin | −6.849 | −42.993 | −18.5 | −4.81 | −19.44 | 2.55 | −3.67 | −3.49 | 39.42 | −32 | Conventional Hydrogen | Electrostatic π-Cation: |
| Bond: Lys633 (2.27), | Lys633 (4.79), | |||||||||||
| Thr636 (1.93), Thr636 | π-Anion: Glu700 (4.20) | |||||||||||
| (2.31), Leu634 (1.77), | π-π Stacked: His635 | |||||||||||
| Gln648 (2.41), Thr636 | (4.42) | |||||||||||
| (1.98), | π-π T-shaped: His635 | |||||||||||
| Carbon Hydrogen | (5.64) | |||||||||||
| Bond: His635 (2.47), | ||||||||||||
| Glu700 (2.51) | ||||||||||||
| Verapamil | −3.182 | −61.455 | −14.78 | −3.19 | −2.4 | −1.22 | −3.16 | −4.43 | 43.98 | −46.89 | Conventional Hydrogen | Electrostatic Salt |
| Bond: Lys633 (2.20) | Bridge; Attractive | |||||||||||
| Carbon Hydrogen | Charge: Glu700 (2.35), | |||||||||||
| Bond: Lys633 (3.07), | π-Cation;π-Donor | |||||||||||
| Gln648 (2.74), Glu700 | Hydrogen Bond: Lys633 | |||||||||||
| (2.82), Gln648 (2.72), | (3.19), | |||||||||||
| Gln648 (2.45), Leu634 | π-Cation: Lys 606 | |||||||||||
| (2.58), Leu634 (2.95), | (4.09), | |||||||||||
| Glu610 (2.66), Ile699 | π-Anion: Glu610 (3.96), | |||||||||||
| (2.60), Thr636 (2.66), | Glu700 (4.01), | |||||||||||
| Ile699 (2.75), Glu700 | Alkyl: Ala585 (4.35), | |||||||||||
| (2.80), Gln648 (2.75), | π-Alkyl: His635 (4.66), | |||||||||||
| Lys633 (2.82) | His635 (4.07), Ala585 | |||||||||||
| (4.58), Lys633 (4.49) | ||||||||||||
| Nitric oxide synthase (PBD ID: 1M9R) | ||||||||||||
| Vanillic acid | −6.534 | −25.711 | −13.92 | −2.82 | −44.7 | 1.9 | −2.83 | −16.06 | 81 | −27.64 | Conventional Hydrogen | π-π Stacked: Trp445 |
| Bond: Ser102 (2.72), | (5.16), Trp447 (4.60), | |||||||||||
| Carbon Hydrogen | Trp447 (3.81), | |||||||||||
| Bond: Phe460 (2.68) | Alkyl: Val104 (5.13), | |||||||||||
| π-Alkyl: Val104 (5.37), | ||||||||||||
| Ala446 (5.17) | ||||||||||||
| Verapamil | −3.539 | −41.256 | −41.97 | −15.00 | 17.01 | 0.52 | −2.11 | −12.09 | −9.41 | −34.32 | Conventional Hydrogen | Electrostatic Salt |
| Bond: Thr80 (2.31), | Bridge;Attractive | |||||||||||
| Lys436 (2.40), Gln462 | Charge: Glu432 (2.29), | |||||||||||
| (2.15), | Alkyl: Leu431 (4.66), | |||||||||||
| Carbon Hydrogen | π-Alkyl: Leu431 (5.13) | |||||||||||
| Bond: Ser78 (2.72), | ||||||||||||
| Ser78 (2.42), Ile79 (2.46), | ||||||||||||
| Val71 (2.48), Ser78 | ||||||||||||
| Acetylcholinesterase (PBD ID: 4EY6) | ||||||||||||
| Isoorientin | −15.180 | −51.937 | −67.31 | −26.55 | −50.13 | 6.57 | −2.45 | −17.67 | 43.31 | −38.03 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (1.87), | (5.42), Trp286 (5.17), | |||||||||||
| Ser293 (1.74), Tyr337 | Trp286 (4.02), | |||||||||||
| (1.79), | π-π T-shaped: Tyr124 | |||||||||||
| Carbon Hydrogen | (5.49) | |||||||||||
| Bond: Gly121 (2.50), | ||||||||||||
| Val294 (2.85), | ||||||||||||
| π-Donor Hydrogen | ||||||||||||
| Bond: Lys633 (3.01), | ||||||||||||
| Tyr124 (2.99), Trp286 | ||||||||||||
| (2.10) | ||||||||||||
| Epigallocatechin | −11.787 | −42.294 | −43.31 | −15.58 | −39 | 5.49 | −3.61 | −18.41 | 40.5 | −23.62 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (2.20), | (4.76), | |||||||||||
| Tyr341 (1.61), Asp74 | π-π T-shaped: Tyr72 | |||||||||||
| (1.86) | (5.01) | |||||||||||
| π-Alkyl: Trp286 (5.17), | ||||||||||||
| Trp286 (3.82), Tyr341 | ||||||||||||
| (5.24), Leu76 (5.44) | ||||||||||||
| Gancaonin E | −11.706 | −55.142 | −31.99 | −10.66 | −16.94 | 7.62 | −1.54 | −28.71 | 64.06 | −50.53 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Gly121 (2.95), | (5.55), Trp286 (4.29), | |||||||||||
| Phe295 (2.45), Tyr341 | π-π T-shaped: Tyr124 | |||||||||||
| (3.03) | (5.73), Phe338 (5.38), | |||||||||||
| π-Alkyl: Trp86 (5.09), | ||||||||||||
| Trp86 (4.59), Trp86 | ||||||||||||
| (4.99), Trp86 (3.83), | ||||||||||||
| Tyr124 (4.70) | ||||||||||||
| Ellagic Acid | −11.403 | −38.936 | −68.58 | −26.00 | −32.35 | 4.93 | −2.45 | −20.64 | 24.98 | −30.32 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Tyr72 (2.71), | (5.75), Trp286 (4.59), | |||||||||||
| Phe295 (1.89), Ser293 | Trp286 (4.61), Trp286 | |||||||||||
| (1.79), Tyr124 (1.92), | (4.09), Trp286 (3.74), | |||||||||||
| Tyr337 (2.56), | Trp286 (3.76), Trp286 | |||||||||||
| Carbon Hydrogen | (4.64), | |||||||||||
| Bond: Val294 (2.53) | π-π T-shaped: Tyr124 | |||||||||||
| (5.71) | ||||||||||||
| Scaposin | −11.072 | −47.758 | −63.35 | −24.28 | −24.55 | 6.63 | −1.75 | −22.1 | 30.64 | −43.95 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (1.92), | (5.40), Trp286 (5.13), | |||||||||||
| Tyr341 (2.40), Tyr337 | Trp286 (4.01), | |||||||||||
| (1.82), | π-π T-shaped: Tyr124 | |||||||||||
| Carbon Hydrogen | (5.43), Tyr337 (5.26), | |||||||||||
| Bond: Gly121 (2.44), | π-Alkyl: Tyr72 (5.41), | |||||||||||
| Val294 (2.87), Asp74 | Trp286 (4.65), Trp286 | |||||||||||
| (2.68), | (3.96), Phe295 (5.19), | |||||||||||
| π-Donor Hydrogen | Phe297 (4.21), Phe338 | |||||||||||
| Bond: Tyr124 (2.88) | (5.19), Tyr341 (4.65), | |||||||||||
| His447 (5.24), His447 | ||||||||||||
| (4.78) | ||||||||||||
| Isosilybin A | −11.052 | −55.882 | −45.74 | −16.64 | −22.95 | 7.75 | −3.01 | −37.52 | 63.22 | −44.57 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (2.29), | (4.55), Trp286 (3.77), | |||||||||||
| Ser293 (2.63), Glu202 | π-π T-shaped: Tyr124 | |||||||||||
| (2.29), | (5.29), Tyr337 (5.34), | |||||||||||
| Carbon Hydrogen | Tyr341 (4.72), | |||||||||||
| Bond: Val294 (2.33), | Amide-π Stacked: | |||||||||||
| Trp86 (2.79), Gly120 | Gly120 (4.12), Gly121 | |||||||||||
| (2.93), | (4.12), | |||||||||||
| π-Donor Hydrogen | π-Alkyl: Trp86 (5.25), | |||||||||||
| Bond: Tyr124 (2.08) | Trp86 (5.37), Tyr124 | |||||||||||
| (5.48), Tyr337 (5.26), | ||||||||||||
| Phe338 (5.34), | ||||||||||||
| Quercetin | −10.967 | −49.91 | −23.66 | −6.61 | −40.59 | 3.59 | −3.17 | −10.54 | 67.32 | −29.99 | Conventional Hydrogen | Electrostatic π-Anion: |
| Bond: Tyr133 (2.38), | Asp74 (4.92), | |||||||||||
| Ser125 (1.80), Glu202 | π-π Stacked: Trp86 | |||||||||||
| (1.88), His447 (1.74), | (4.06), Trp86 (3.78), | |||||||||||
| Tyr72 (1.81), Tyr72 | Trp86 (4.48), Trp86 | |||||||||||
| (1.62), | (3.65), | |||||||||||
| Carbon Hydrogen | π-π T-shaped: Tyr124 | |||||||||||
| Bond: Gly126 (2.77) | (5.23) | |||||||||||
| 14,15-Dehydro- | −10.376 | −34.243 | −43.29 | −15.57 | −28.97 | 2.46 | −0.78 | −18.44 | 43.75 | −35.9 | Conventional Hydrogen | Electrostatic π-Cation: |
| 16-epi- | Bond: Tyr341 (1.81), | Trp286 (4.69), | ||||||||||
| vincamine | Carbon Hydrogen | π-π Stacked: Trp286 | ||||||||||
| Bond: Tyr72 (3.10), | (5.03), Trp286 (4.68), | |||||||||||
| Gln291 (2.74) | Trp286 (3.88), | |||||||||||
| Alkyl: Leu289 (4.58) | ||||||||||||
| Enterolactone | −9.057 | −34.759 | −49.58 | −18.30 | −27.93 | 7.71 | −2.24 | −30.84 | 34.56 | −26.12 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (1.78), | (4.80), Trp286 (4.77), | |||||||||||
| Asp74 (1.92), | π-π T-shaped: Tyr72 | |||||||||||
| Carbon Hydrogen | (4.85), Tyr124 (5.09), | |||||||||||
| Bond: Phe295 (2.64), | Tyr337 (5.29), Tyr341 | |||||||||||
| π-Donor Hydrogen | (5.88) | |||||||||||
| Bond: Tyr124 (2.19) | ||||||||||||
| Scopoletin | −8.202 | −26.348 | −36.51 | −12.63 | −19.46 | 2.29 | −1.13 | −18.24 | 24.79 | −20.74 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (1.75), | (4.92), Tyr341 (4.82), | |||||||||||
| Tyr337 (1.91), | Tyr341 (3.96), | |||||||||||
| Carbon Hydrogen | π-π T-shaped: Tyr124 | |||||||||||
| Bond: Val294 (2.54), | (5.61), | |||||||||||
| Tyr124 (2.46), Tyr124 | π-Alkyl: Tyr72 (4.61), | |||||||||||
| (2.65) | Tyr124 (4.98) | |||||||||||
| Biochanin A | −7.901 | −34.324 | −38.01 | −13.28 | −17.33 | 4.71 | −1.28 | −9.95 | 26.69 | −31.73 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (1.97), | (5.11), Trp286 (4.75), | |||||||||||
| Tyr337 (2.79), | Trp286 (3.74), Trp286 | |||||||||||
| Carbon Hydrogen | (4.92), Trp286 (4.33), | |||||||||||
| Bond: Trp286 (2.65) | Alkyl: Leu289 (5.05) | |||||||||||
| Vasicine | −7.848 | −30.824 | −41.04 | −14.59 | −21.56 | 0.44 | −0.59 | −8.98 | 20.65 | −26 | Conventional Hydrogen | π-π Stacked: Trp286 |
| Bond: Phe295 (1.80), | (4.39), Trp286 (5.65), | |||||||||||
| Carbon Hydrogen | π-π T-shaped: Tyr124 | |||||||||||
| Bond: Val294 (2.61), | (5.64), | |||||||||||
| Tyr341 (2.78), Ser293 | π-Alkyl: Trp286 (4.19) | |||||||||||
| (2.52), Ser293 (2.66), | ||||||||||||
| Tyr341 (2.90) | ||||||||||||
| Galanthamine | −9.742 | −43.067 | −23.61 | −7.02 | −62.45 | 3.84 | −1.62 | −30.51 | 88.88 | -20.93 | Conventional Hydrogen | Electrostatic Attractive |
| Bond: Ser203 (2.00), | Charge: Asp74 (4.50), | |||||||||||
| Glu202 (1.74), Tyr337 | π-π T-shaped: Tyr337 | |||||||||||
| (2.16), | (5.83), | |||||||||||
| Carbon Hydrogen | Amide-π Stacked: | |||||||||||
| Bond: His447 (2.41), | Gly121 (4.04), Gly122 | |||||||||||
| Tyr133 (2.86), Ser125 | (4.04), | |||||||||||
| (2.91), Asp74 (2.59) | π-Alkyl: Trp86 (4.19), | |||||||||||
| Trp86 (3.91), Phe295 | ||||||||||||
| (4.18), Phe297 (4.50), | ||||||||||||
| Phe338 (4.66), His447 | ||||||||||||
| (4.64) | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khurm, M.; Guo, Y.; Wu, Q.; Zhang, X.; Ghori, M.U.; Rasool, M.F.; Imran, I.; Saqib, F.; Wahid, M.; Guo, Z. Conocarpus lancifolius (Combretaceae): Pharmacological Effects, LC-ESI-MS/MS Profiling and In Silico Attributes. Metabolites 2023, 13, 794. https://doi.org/10.3390/metabo13070794
Khurm M, Guo Y, Wu Q, Zhang X, Ghori MU, Rasool MF, Imran I, Saqib F, Wahid M, Guo Z. Conocarpus lancifolius (Combretaceae): Pharmacological Effects, LC-ESI-MS/MS Profiling and In Silico Attributes. Metabolites. 2023; 13(7):794. https://doi.org/10.3390/metabo13070794
Chicago/Turabian StyleKhurm, Muhammad, Yuting Guo, Qingqing Wu, Xinxin Zhang, Muhammad Umer Ghori, Muhammad Fawad Rasool, Imran Imran, Fatima Saqib, Muqeet Wahid, and Zengjun Guo. 2023. "Conocarpus lancifolius (Combretaceae): Pharmacological Effects, LC-ESI-MS/MS Profiling and In Silico Attributes" Metabolites 13, no. 7: 794. https://doi.org/10.3390/metabo13070794