2. Materials and Methods
2.1. Chemicals
The following reagents were purchased from Sigma-Aldrich (Saint Quentin Fallavier, France): 2,2-diphenyl-1-picrylhydrazyl (DPPH), potassium persulfate (99.99%), 3-(2-pyridyl)-5,6-diphenyl-1,2,4-triazine-p,p′-disulfonic acid monosodium salt hydrate (97%), copper (II) chloride (99.99%), 2,4,6-tris(2-pyriodyl)-s-triazine (TPTZ) (≥98%), sodium acetate (≥99%), 2,2′-azobis(2-methylpropionamidine) dihydrochloride granular (97%), trolox (≥98%), ferric chloride (97%), neocuproine (≥98%), glacial acetic acid, formic acid, ammonium acetate and ferric chloride hexahydrate. The 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) (98%) reagent was purchased from ThermoFisher-Alfa Aesar, (Kandel, Germany).
The following enzymatic kits were purchased from Thermo Fischer Scientific (Waltham, MA, USA): EnzChek Gelatinase/Collagenase assay kit E-12055 and EnzChek Elastase assay kit E12056. Xanthine oxidase inhibition was measured using an assay kit 19160 purchased from Sigma Aldrich (Saint Quentin Fallavier, France).
The ultrapure water was produced with a PurelabFlex system from Veolia (Wissous, France). The acetonitrile used for the semi-preparative fractionation was HPLC grade, purchased from VWR, (Fontenay-sous-Bois, France). The acetonitrile from SDS Carlo Erba (Val de Reuil, France), used for the UHPLC-HRMS analysis, was a HPLC PLUS gradient grade.
The ethanol used for the oak wood extraction was HPLC grade and was purchased from VWR (Fontenay-sous-Bois, France). The formic acid used for the UHPLC-HRMS analysis was of an Optima LC-MS grade from Fisher Scientific (Illkirch-Graffenstaden, France).
3. Results and Discussion
The objective of this study was to assess the potential of oak heartwood extracts as anti-aging ingredients, to determine if the activity was dependent on the species of oak used, since the molecular composition can vary depending on the species, and to target the compound families involved in the activity. The literature reports that sessile oak is statistically richer in aromatic compounds while pedunculate oak is richer in tannins [
21]. In addition, pedunculate oak contains compounds such as bartogenic acid or terpenes in dimeric forms while sessile oak contains other types of terpenes such as quercotriterpenosids that are glycosylated terpenes [
22,
23,
24,
25]. The molecular composition of pubescent oak remains undescribed.
To target anti-aging molecules, crude extracts of the three oak species were fractionated, the fractionation was then followed by UHPLC-DAD-HRMS/MS, and the molecular composition of each fraction was determined and compared to that of the crude extract using molecular networks. The different clusters highlighted the molecular families detected in each fraction.
The antioxidant and anti-enzymatic potentials of the total extract and the different fractions of the three species were then evaluated. The results obtained for the total extracts of the three species were compared to determine if the activity depended on the species. The results within the same species were also compared to highlight which fraction and, therefore, which molecular family was involved in the activity of the total extract.
3.2. Comparison of the Chemical Composition of Fractions and Crude Extract
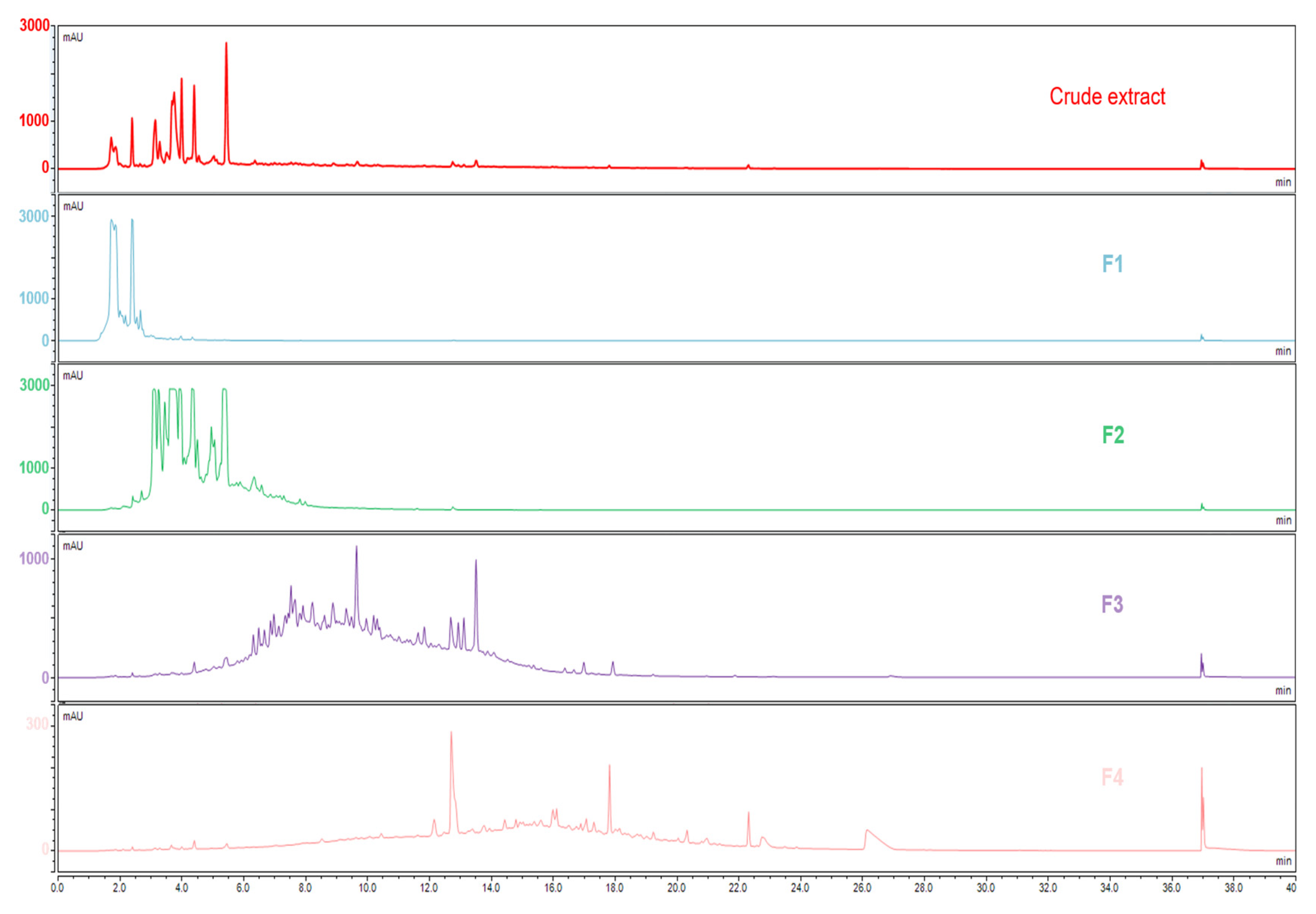
The generation of the molecular network was based on the mass spectrometry analysis of the crude extracts and fractions. The MS/MS spectra of the compounds were compared pairwise to find similarities in their fragmentation pathways, i.e., the same fragment ions or similar neutral losses. To obtain suitable MS/MS spectra with a convenient number of fragment ions, the collision energies were optimized to find the ones that enabled fragmentation of the rigid structures while obtaining good spectra for the molecules that fragmented more easily. The molecules presenting similar fragmentation patterns were grouped in the same cluster [
19,
26,
27]. As an example,
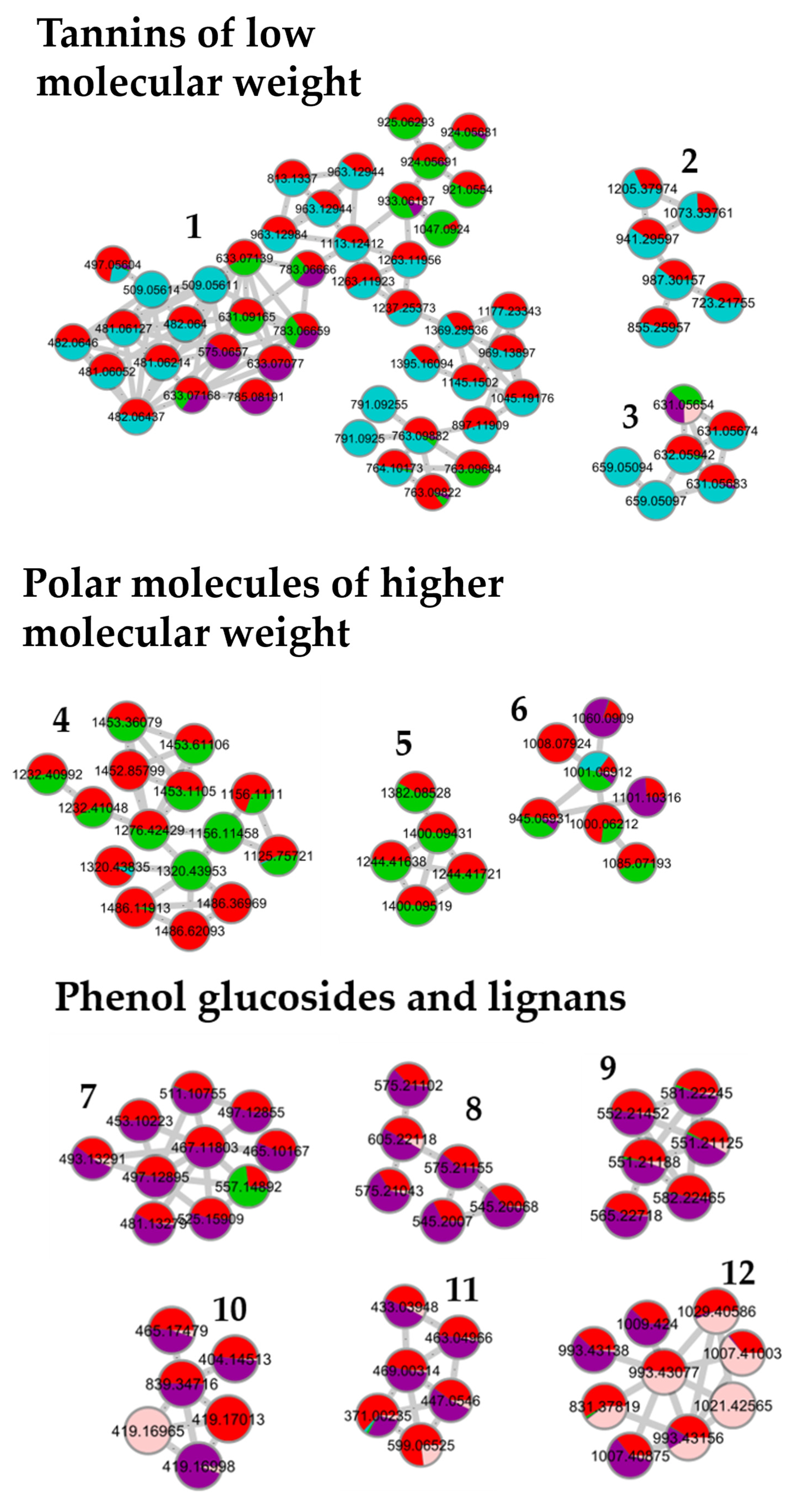
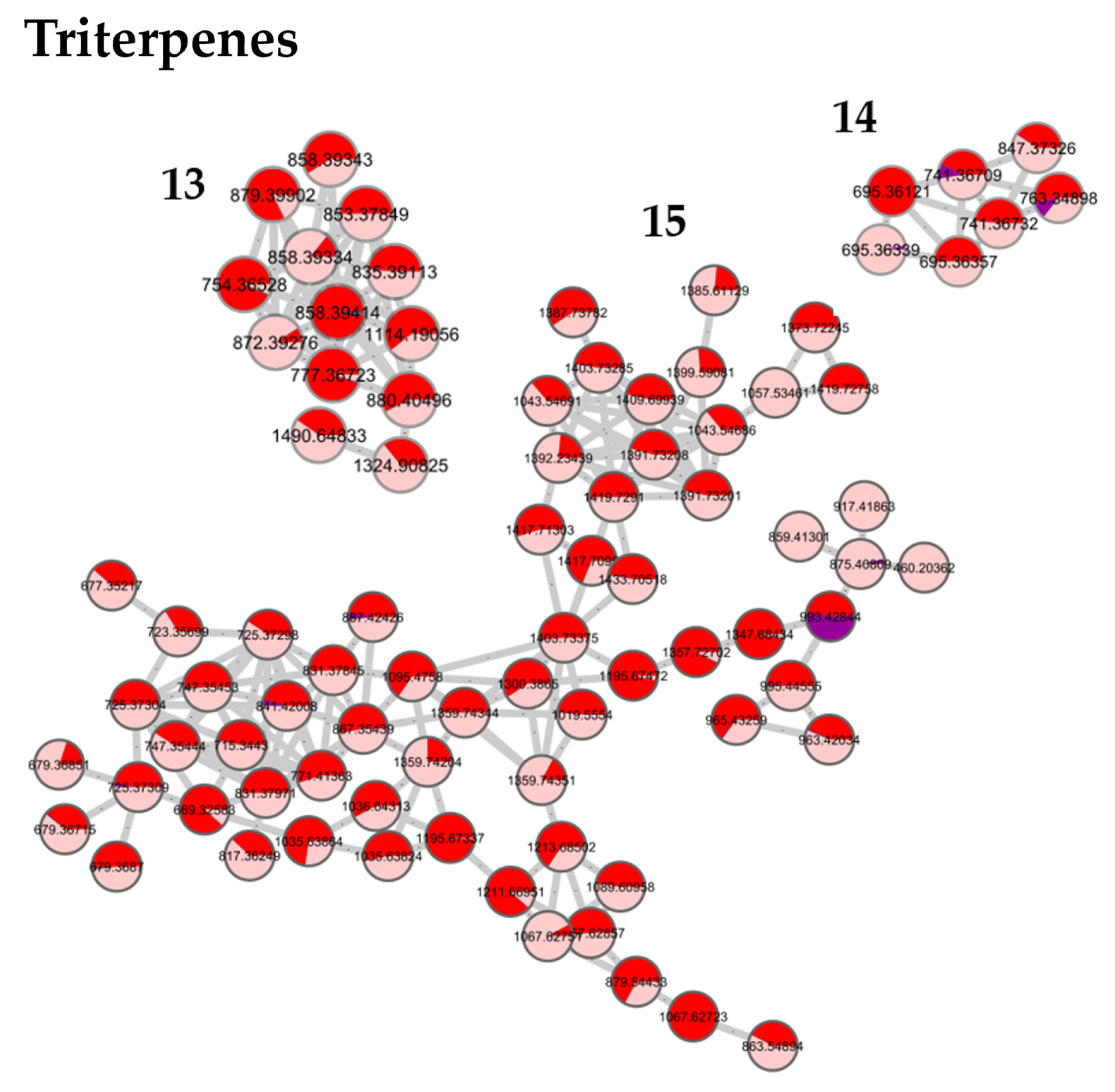
Figure 3 shows the results obtained for the pedunculate oak. A similar distribution of fractions was obtained for the sessile and pubescent oak. According to the selected parameters, 13 clusters containing at least five nodes were formed. Based on previous work, on literature data, and on the interpretation of mass spectra and databases, the network was annotated [
22,
23,
28]. Clusters 1 to 3 contained low molecular weight tannins and Clusters 4 to 6 polar molecules of a higher molecular weight. Clusters 7 to 12 were composed of phenols, glucosides, and lignans, while Clusters 13 to 15 contained triterpenes.
Clusters 1 to 12 were identical between the three species while clusters 13 to 15 were triterpenes derived from quercotriterpenosids for the sessile oak and derived from roburgenic acid for the pedunculate and pubescent oaks.
In
Figure 3, the network of the crude extract (red) is associated with the network of each fraction to evaluate the distribution of molecules in the different fractions of each species. It confirms that only a small proportion of the molecules observed in the network were shared between several fractions. For example, ion
m/
z 783.0066 in Cluster 1 was distributed between Fraction 2 and 3 in a proportion that was rather similar; however, some molecules of a different polarity but belonging to the same cluster and, therefore, to the same molecular family, may have been eluted in different fractions. For example, Cluster 1 contained the most molecules eluted in Fraction 1 but also in Fractions 2 and 3.
Fraction 1 in blue, the most polar fraction, was divided into three Clusters 1, 2, and 3. It was composed of low-molecular mass ellagitannins with common fragments of ellagic acid such as the well-known vescalin/castalin suggested in Cluster 3 or other small-polar molecules such as HHDP glucose suggested in Cluster 1 [
22].
Fraction 2 in green, was mainly composed of ellagitannins with a higher molecular weight than in Fraction 1, that were distributed in Clusters 1, 4, 5, and 6 with losses of vescalagin or ellagic acid [
29].
Fraction 3 in purple, mainly grouped the Clusters 7, 8, 9, 10, and 11. These clusters highlighted molecular families of polyphenols that differed from the first two fractions, namely, phenol glucosides such as 3,4,5-trimethoxyphenyl-(6′-O-galloyl)-O-
β-glucopyranoside in Cluster 6 or lignans such as lyoniresinol in Cluster 10 [
30]. Cluster 9 was composed of molecules that had lyoniresinol as a fragment [
31].
These three fractions were composed of phenolic compounds. In accordance with the literature, the high content of ellagitannins in oak wood was well identified in the heaviest Fractions 1 and 2. Other phenolic compounds were identified in lower concentrations in Fraction 3.
Fraction 4 in pink, was mainly composed of terpenes, which are a well-known component of pedunculate oak. They were distributed in Clusters 12, 13, 14, and 15, such as triterpenes in dimeric form of the roburosid family or derivatives from bartogenic acid.
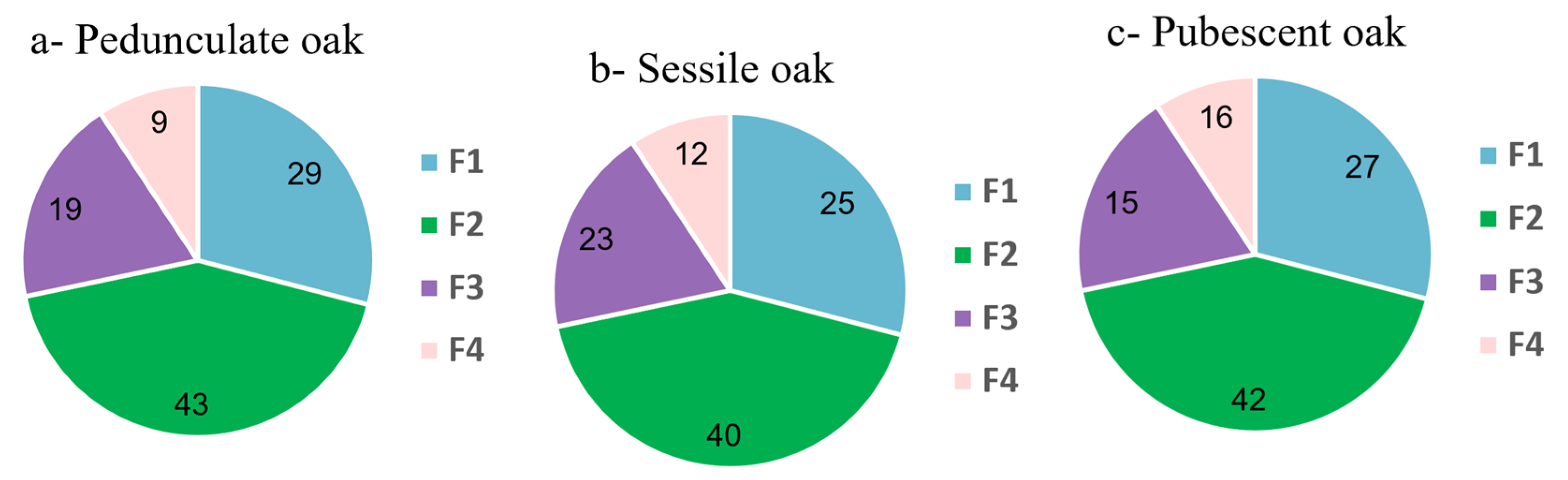
For the other two species the distribution was similar. The first three fractions were equivalent to those of the pedunculate oak. Fractions 1 and 2 were the heaviest for each of the three species and were composed of polyphenols, in particular ellagitannins. Fraction 3, which was heavier in the sessile and lighter in the pubescent oak, was also common to the three species. It was composed of lignan and phenol glucosides in the molecular network. Fraction 4, whose proportion varied between the species from 9% in the pedunculate to 16% for the pubescent and 12% for the sessile oaks, also differed in its molecular composition according to the species. For the pedunculate and pubescent oaks, the same type of molecule was found, namely, triterpenes such as bartogenic acid derivatives. For the sessile oak, other types of terpenes were found, mainly glycosylated terpenes including quercotriterpenosides [
22,
23,
25].
In summary, this strategy allowed the rapid identification of molecular families contained in the crude extract by clustering compounds according to their structures without interpreting all the MS spectra. Moreover, it made it possible to highlight the molecular distribution of the compounds in each fraction. The three oak extracts had many molecules in common, with all the common polar parts composed of sugars and polyphenols. The three extracts were composed of low- and high-molecular mass ellagitannins, lignans and phenol glucosides. Finally, it was the most apolar fraction containing terpenic compounds that differed the most between the species; however, this fraction only represented about 10–15% of the crude extract and even if the molecules differed, they were all derived from pentacyclic triterpenes of the oleanolic acid type.
3.3. In Vitro Antioxidant Capability
A complete evaluation of the antioxidant activity of the oak crude extracts and all the semi-preparative fractions was performed using five different types of assays to measure the antioxidant properties. The assays were based on hydrogen atom transfer (DPPH and ABTS), single electron transfer mechanisms (FRAP, CUPRAC) or iron (II) chelating assays.
For each test, three concentrations of extracts were tested to verify the dose/response effect of the test. The whole results of the five antioxidant assays expressed as an activity percentage for a final sample concentration in the well of 25 µg/mL are presented in
Table S1 in the supplementary data.
Crude extracts of the three species showed a good activity for each test. Some fractions also appeared to have an antioxidant activity.
For the crude extracts, the DPPH and ABTS tests showed a high percentage of inhibition for each of the three species. The results for the same test were similar, with about 72% of inhibition for the DPPH test and 96% for the ABTS test. While the reference standard trolox gave a 100% inhibition percentage at 40 µg/mL (i.e., the final concentration in well), the antioxidant activity for the ABTS test remained stronger than the DPPH test.
For the FRAP and CUPRAC tests, the activity results were more contrasted between the species with the highest activity for the pedunculate oak, then the pubescent and finally the sessile oak. For the FRAP test, there was very good activity between 80 and 90% while the activity of the CUPRAC test was less than this, ranging from 40 to 53%. Trolox gave similar inhibition percentages at 25 µg/mL for the FRAP and 50 µg/mL for the CUPRAC (i.e., the final concentration in well).
The activity for the iron chelation test was also very good and identical between the three species. The concentration in the reference standard trolox, which gave the same inhibition percentage, was around 50 µg/mL.
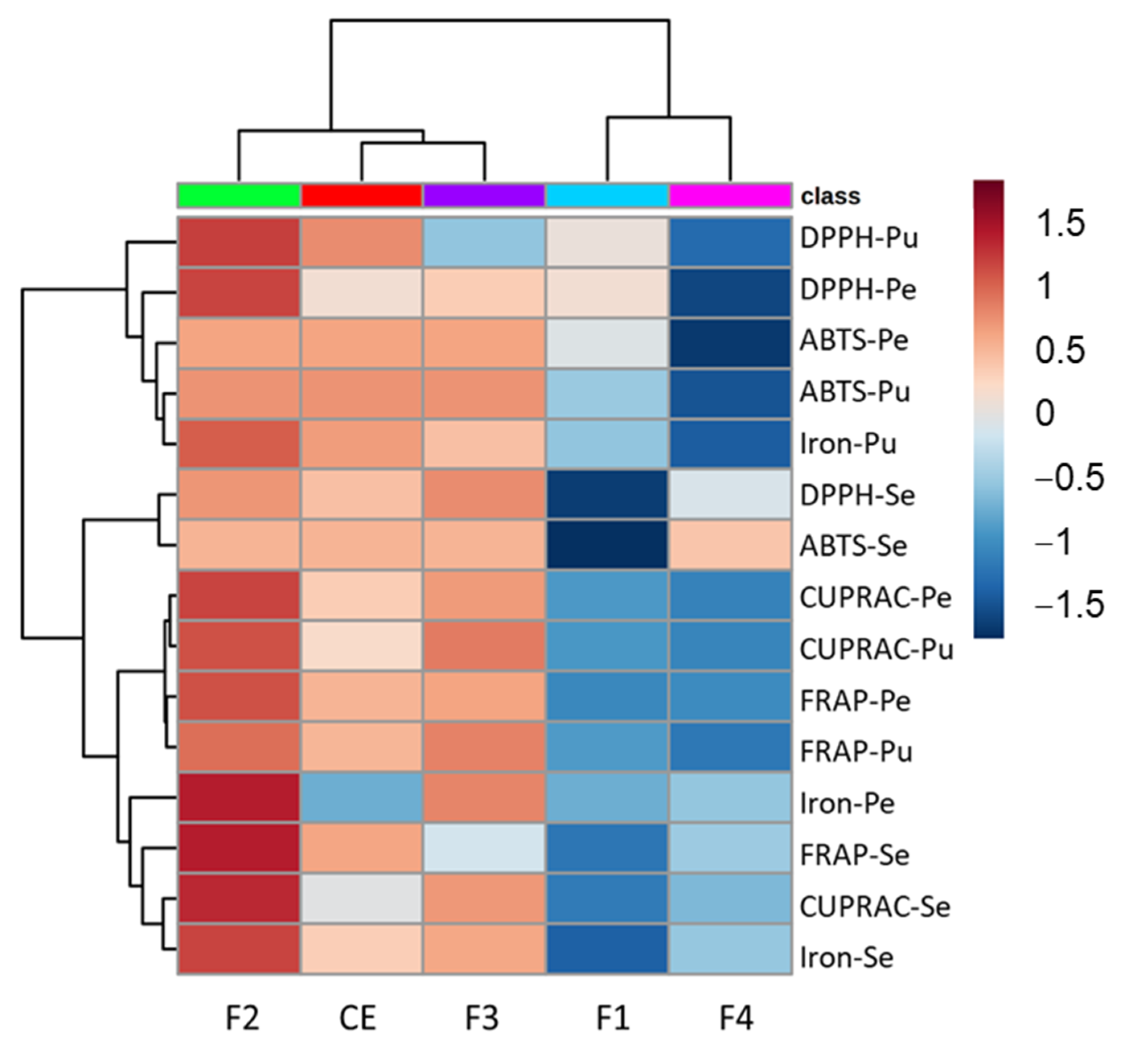
Figure 4 presents the heat map correlating the crude extract, fractions and antioxidant activity.
The heat map shows a more similar behavior of the pubescent and pedunculate oaks that were clustered together while the sessile oak was dissociated from these two species for each activity test. The DPPH and ABTS assays based on the hydrogen atom transfer are clustered in the upper part of the heat map with the iron chelation for the pubescent oak, while the FRAP and CUPRAC based on single electron transfer mechanisms and iron chelation for the pedunculate and sessile oaks are clustered together in the lower part of the heat map, highlighting the similar mechanism involved for these tests.
Fraction 2 showed the higher activity for most of the assays, in particular for the DPPH, CUPRAC and FRAP assays for the pedunculate and pubescent oak, as well as for the FRAP, CUPRAC and iron chelation for the sessile oak. Fraction 3 was also quite active with an intensity close to that of the crude extract. Fractions 1 and 4 were clustered together showing a lower activity. Fraction 1 showed a slight activity on the DPPH assay for the pubescent and pedunculate oaks, while Fraction 4 of the sessile was a little active on the ABTS assay.
In summary, the various antioxidant tests showed good activity of the total extract for the three species of oak. The transfer of protons involved in the DPPH and ABTS tests was equally effective for all species, and the same holds for the iron chelation. On the contrary, the reducing potential was lower, especially for the CUPRAC test, which had the lowest response compared to the other four tests. Moreover, for each assay, the oak extracts presented a higher or similar antioxidant capability than the reference standard trolox that demonstrated the good potential of these extracts.
The antioxidant activity has already been proven in the literature to be correlated with polyphenol oak richness [
32].
For the fractions, the activity was generally greater for Fractions 2 and 3, correlated with the molecular composition of these fractions, which were rich in tannins including vescalagin or castalagin derivatives for Fraction 2 and phenol glucosides for Fraction 3 [
29].
In all the tests, the activity of Fraction 1 of the sessile oak was generally lower than that of the other species and Fraction 3 was also found to be less active in the FRAP test.
Fraction 4 of the pubescent oak appeared to be less active than for the other two species. The pedunculate and pubescent oaks showed, for the crude extract and fraction, a similar behavior for the evaluated assays, while the sessile oak presented, for some of them, different results. In addition, each fraction participated in the overall activity of the total extract, which involved several different molecular families in the antioxidant response [
33].
3.4. Enzymatic Tests
To assess the anti-aging potential of the oak extract, the crude extracts and fractions were tested on three enzymes: xanthine oxidase, which is involved in the defense mechanism of ROS, and collagenase and elastase, which are involved in the loss of elasticity of the skin caused by the degradation of collagen and elastin.
Preliminary tests excluded tyrosinase which was not inhibited by the crude oak extract.
Table S2 in the supplementary data presents the results of the three enzymatic assays expressed as inhibition percentages. Several concentrations of extracts were tested to verify the dose/response effect of the tests. These concentrations were adapted to each test according to the response.
Good xanthine oxidase activity was measured with low tested concentrations. The three species were very active with an inhibition percentage of 99%, which saturated at 8.5 µg/mL. At this concentration all the fractions were highly active. The concentration had to be reduced to 4 µg/mL to observe a dose/response effect. Fraction 1 appeared to be less active compared to the crude extract for each species. These results complemented the previous antioxidant colorimetric tests showing good activity of the extracts.
The extracts as well as the fractions showed a weak elastase inhibition. The pedunculate and pubescent oaks were the most active species with 50% of inhibition at 250 µg/mL. At 125 µg/mL the activity was very low, presenting less than 15% of inhibition. In comparison, the reference inhibitor presented 73% of inhibition at 20 µg/mL. Fractions 2, 3 and 4 took part in the activity with an inhibition percentage between 25 and 30% while Fraction 1 was less active. The elastase inhibition of the sessile oak was half (20% of activity at 250 µg/mL) that of the other two species. It was less active on all fractions compared to the other two species.
The strongest activity was anti-collagenase inhibition which reached high percentages at very low tested concentrations of 0.4 and 0.04 µg/mL, while the reference inhibitor was around 80% of inhibition at a concentration of 47 µg/mL. The sessile oak appeared to be a little less active than the other two species. Fractions 2 and 3 were the most active. The least active fraction for the sessile oak was Fraction 4 while for the pedunculate and pubescent oak it was Fraction 1. Overall, each fraction participated in the activity of the crude extract even if Fractions 2 and 3 were more active [
29].
The differences in activity highlighted with Fraction 4 may be linked to the molecular composition of this fraction, which differed for the sessile oak compared to the other two species; the pedunculate and pubescent oak were richer in triterpenes derived from roburgenic acid.
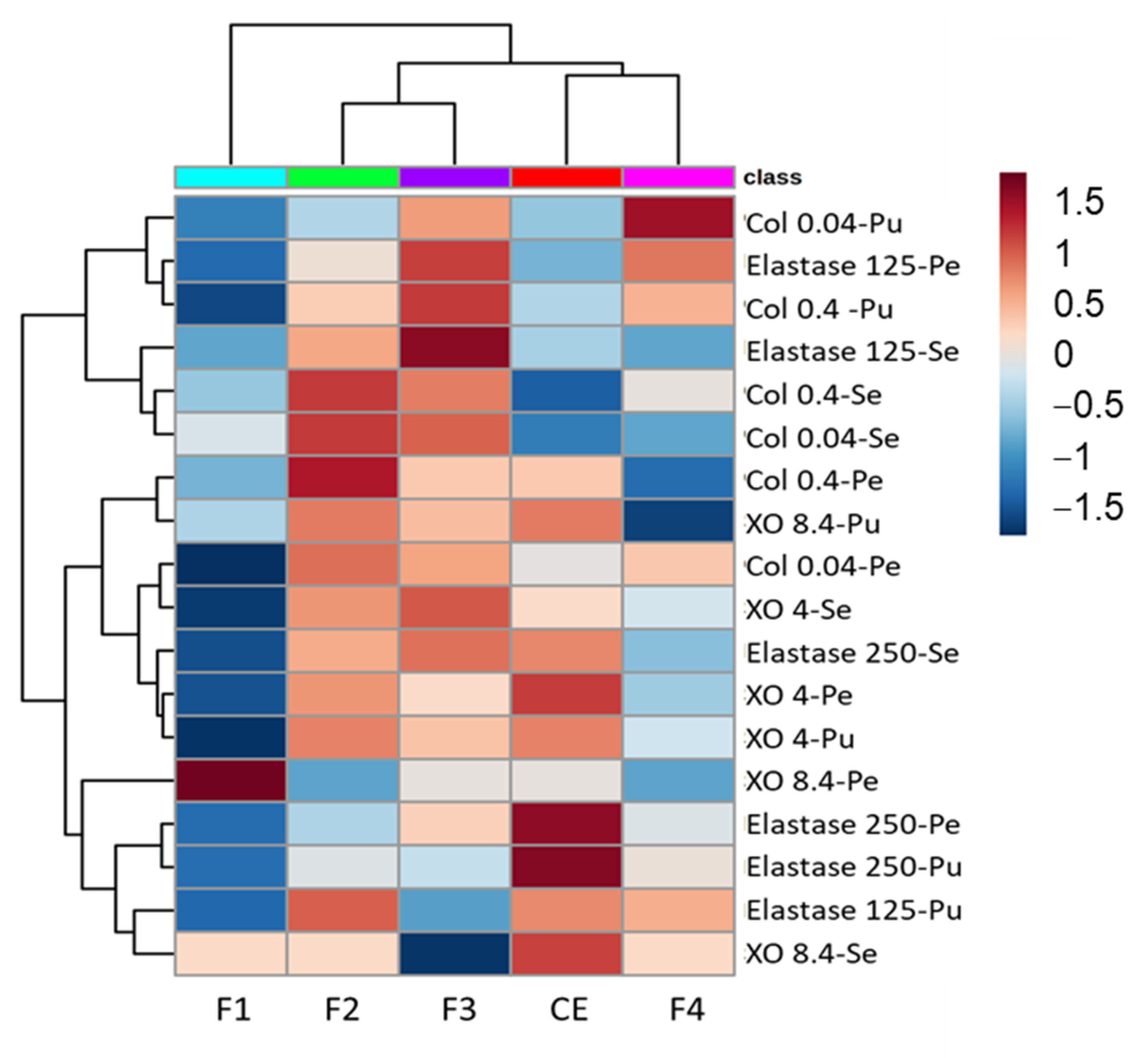
Figure 5 presents the heat map correlating the inhibition potential of the crude extracts and fraction to the enzymatic assays.
Unlike antioxidant activity, no clustering according to the oak species or enzyme tested showed any real similarity or difference in behavior; nevertheless, Fraction 1 showed the lower participation to the inhibitory effect for the main experiments. Fractions 2 and 3 were the most involved fractions, while the Fraction 4 contribution was enzyme- and oak species-dependent.
In conclusion, the colorimetric tests, as well as the xanthine oxidase enzymatic test, showed an overall good antioxidant activity for the three species of oaks. There was only a low difference in response depending on the species, with an activity for the sessile oak being slightly lower, showing that the antioxidant activity was not drastically species-dependent. There were few differences between the fractions, showing that each molecular family participated in the activity of the total extract, even if the compounds contained in Fraction 2 appeared to be mainly involved due to their polyphenolic structure and their abundance.
Whatever the species, the oak extracts presented a rather low inhibition of elastase while the inhibition of collagenase was more efficient at a lower concentration. More differences between the species’ behavior were observed for the enzymatic assays. Sessile oak responded less well in those tests compared to the other two species. This may be due to its most-polar and most- non-polar fractions that showed lower activity than the other fractions. The terpene composition of Fraction 4 was different from that of the pubescent and pedunculate oaks. The pubescent and pedunculate oaks showed more similar behaviors, with a slightly higher activity of the pubescent oak at low concentrations.
3.5. Correlation between Molecular Content and Activity
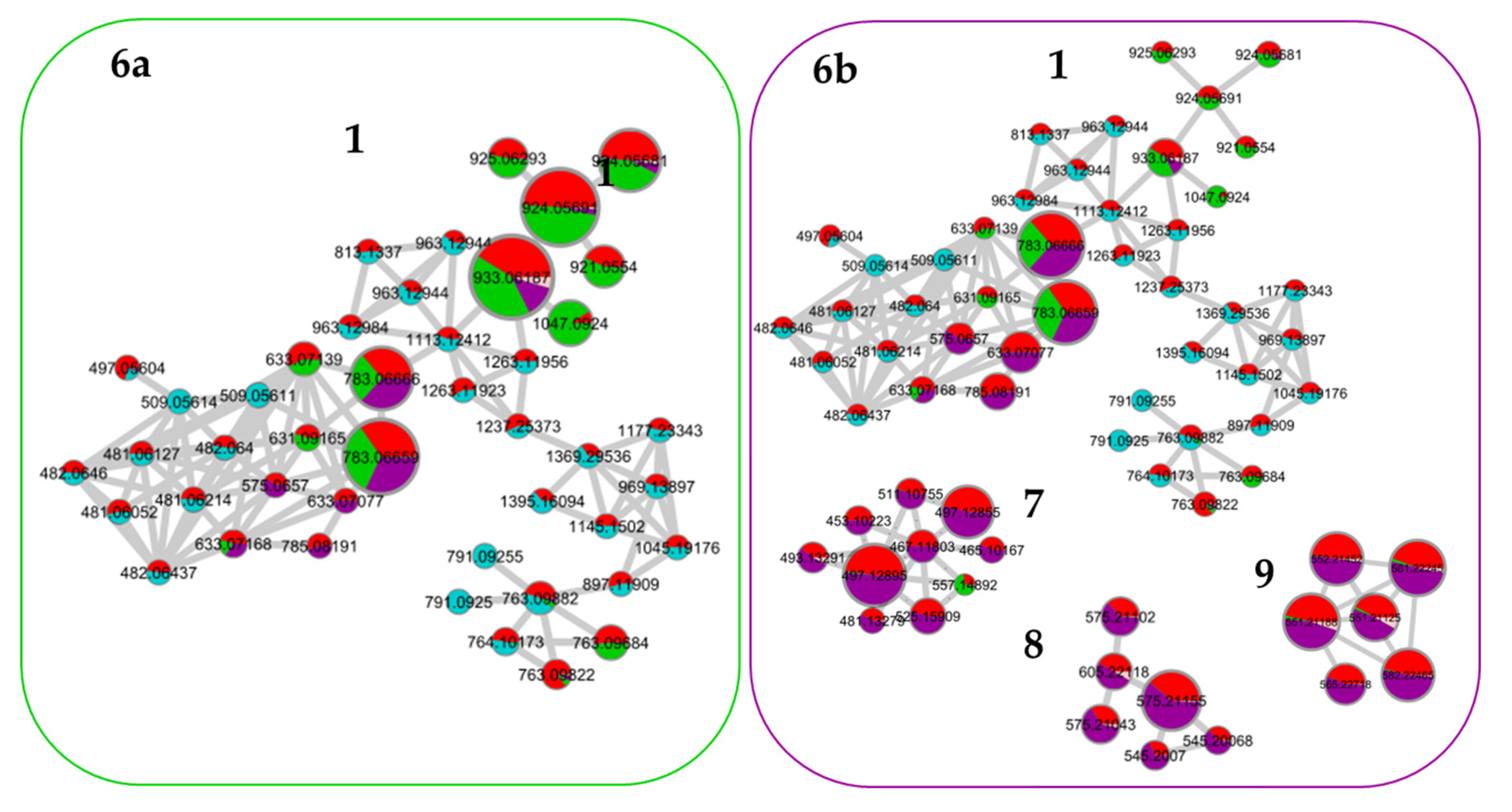
Based on the molecular network of the crude extract with its four fractions presented in
Figure 3, the most abundant molecules in Fractions 2 and 3 have been highlighted in
Figure 6. Indeed, it was previously shown that the fractionation of the extract was undertaken by the molecular family. Within the same molecular family, compounds have similar structures and, thus, similar ionization yields can be assumed. The molecules with the strongest ion intensity in Fractions 2 and 3 are represented by a larger node size in
Figure 6a,b, respectively.
Cluster 1 contained few major molecules contained in both Fraction 2 and 3 with a common molecule
m/
z 783.0665. Other abundant molecules were found for Fraction 2 in the single nodes. For Fraction 3, the majority of molecules were found in Clusters 7, 8, 9 and 10. Using the databases, the interpretation of the mass spectra and the literature, putative identifications were proposed for the most intense molecules in Fractions 2 and 3, and these are presented in the
supplementary data Table S3. The most abundant molecules of Fraction 2 were ellagitannins such as Roburin derivatives, Grandidin, Castalagin, Vescalagin and Pedunculagin. Fraction 3 also presented high amounts of lignane such as Lyoniside and Lyoniresinol derivatives [
29,
31,
34,
35,
36,
37].
To correlate the antioxidant activity to the molecular composition of the oak extract intensity of the ions obtained using UHPLC-HRMS, the percentages of antioxidant and enzymatic activities were examined using a principal component analysis.
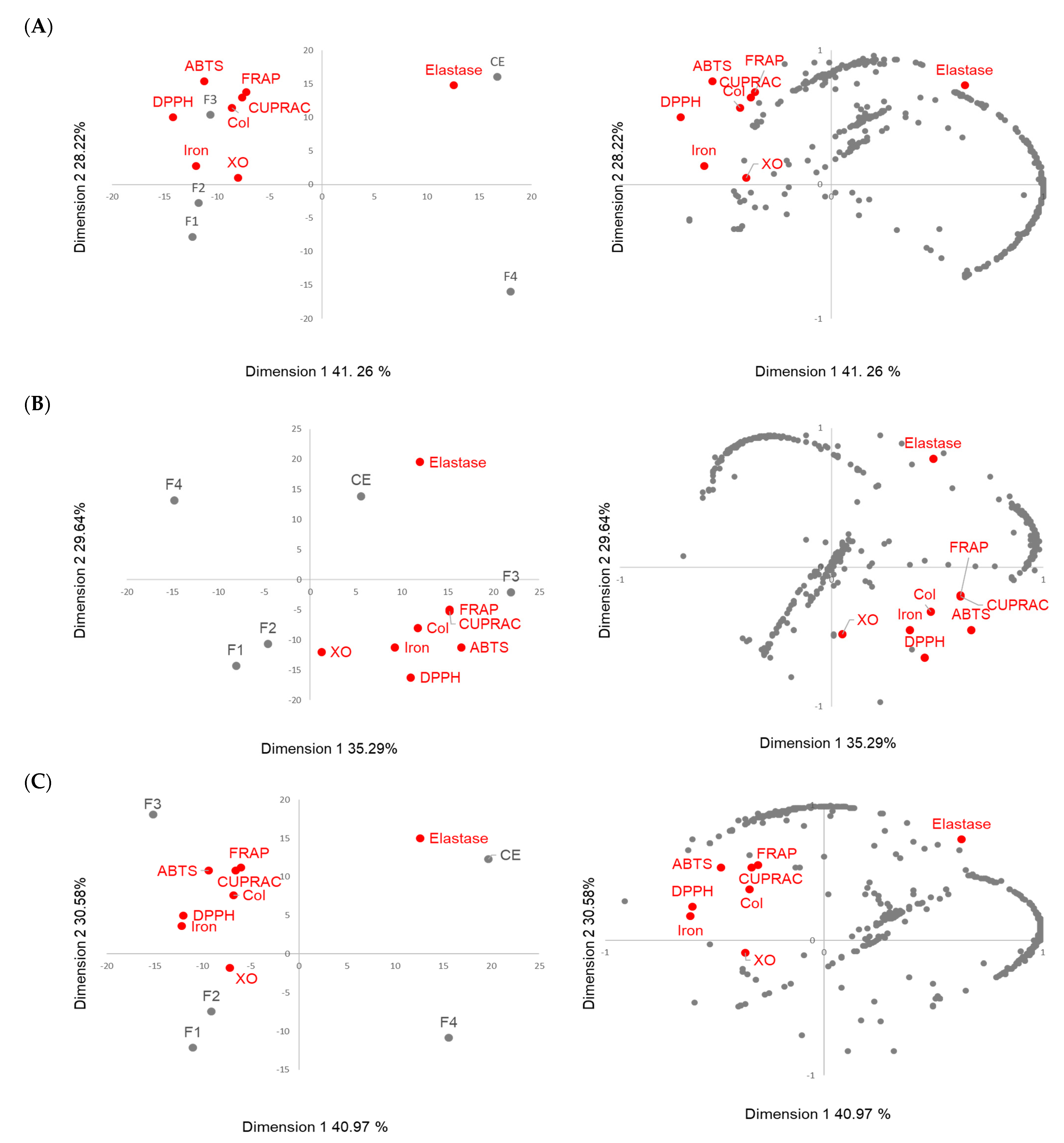
Figure 7 presents the biplot and the loading plot obtained for the three species.
For each of the three species, the first two principal components (PCs) explained around 70% of the initial variance. The pedunculate and pubescent oaks had more similar behavior, and the representations were close (
Figure 7A,C). The CE and F4 were projected in a PC1-positive score while F1, F2 and F3 were projected in a PC1-negative score that represented 41% of the variance for both species. The CE and F3 were projected in a PC2-positive score while F1, F2 and F4 were projected in a PC2-negative score, representing around 30% of the variance. The antioxidant and collagenase activities were mainly correlated with Fraction 3 and the XO activity was close to Fraction 2. Elastase was projected in the opposite side of the PC1 axe near to the crude extract. Fraction 1 was projected in the same side of the PC1 but was far from the activities on the PC2 axis, and Fraction 4 was projected completely away from the activities. Among the molecules projected near the antioxidant activities, the presence of lignanes such as Lyoniside belonging to Clusters 7 to 11 of the molecular network, and ellegitannin derivatives such as Pedunculagin found in Cluster 1, were observed and could contribute to the antioxidant activity of the oak extracts. Concerning the molecules projected close to the elastase, mainly compounds such as bartogenic acid derivatives belonging to the triterpene family found in Clusters 13 and 15 were observed. The pedunculate oak extract appeared to show a greater number of molecules contributing to elastase activity than the pubescent oak extract.
The sessile oak (
Figure 7B) differed from the pedunculate and pubescent oaks since elastase, but also the antioxidant activities, were projected in a PC1-positive score; however, the correlation of activities with the fractions remained the same since F3 was projected near the antioxidant activity, while CE was projected near the elastase one. Fractions 1 and 2 were also projected in a PC2-negative score, where F2 was quite close to the XO and Fraction 4 remained at the opposite side on the PC1 and PC2. On the loading plot, a fewer number of molecules were projected near to activities. Ions were found as a single node in the molecular network and these compounds had not been previously identified in oak extracts.
Gathering the results obtained with the heatmap and ACP, Fractions 2 and 3 were the most active and the molecules they contained appeared to be the main contributors to the extract’s antioxidant activity. Fraction 2 mainly contained ellagitannins, which are well-known for their antioxidant activity. Fraction 3 mainly contained lignans and phenol glucosides, which are also known for their antioxidant activity [
29,
38,
39]. These compounds may also have contributed to the strong collagenase inhibition observed for the oak extracts. The triterpenes of the pedunculate and pubescent oaks that are bartogenic acid derivatives appear to contribute to the elastase activity, unlike the quercotriterpenosids terpene derivatives of the sessile oak.
The comparison of the activities of the three main oak species, namely, sessile, pedunculate and pubescent oak, was carried out for the first time. Oak water extract proves to be an intriguing green ingredient demonstrating an interesting valuation of pubescent oak, which is still seldom exploited. It has both a strong antioxidant activity due to its richness in phenolic compounds such as ellagitannins or lignans and an average anti-elastase activity of 50–60% inhibition at 250 µg/mL, combined with a very strong anti-collagenase activity of around 80% inhibition at 0.4 µg/mL and 50 % at 0.04 µg/mL, that was newly evaluated here. For these two enzymes, the sessile oak species was less effective while the pubescent oak was a little more active than the pedunculate oak.
The oak extracts are rich in different molecular families such as small-polar ellagitannin derivatives, ellagitannins with heaviest molecular weight, that represented the main concentrated molecules in the crude extract, lignans, phenol glucosides and triterpene derivatives. Even if polyphenolic compounds appeared to contribute substantially to the overall activity, few differences in activity between the fractions were observed, indicating that each molecular family contributes to the overall activity. It is, therefore, more interesting to value the total extract to achieve an optimal ingredient effectiveness and to decrease production costs.
Thus, oak water extract could be used to neutralize the ROS produced during oxidative stress and to prevent the degradation of collagen and elastin during skin ageing. Its complementary properties could make oak extract a valued multi-functional ingredient to act against skin ageing. It would be interesting to delve further in the development of a cosmetic ingredient by confirming the in vitro tests with tests on cell cultures or skin models and investigating other types of activity.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







