


Açaí (Euterpe oleracea Mart.) Seed Oil Exerts a Cytotoxic Role over Colorectal Cancer Cells: Insights of Annexin A2 Regulation and Molecular Modeling
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Lyophilized Hydroalcoholic Seed Extract and Oil of Euterpe Oleracea Mart
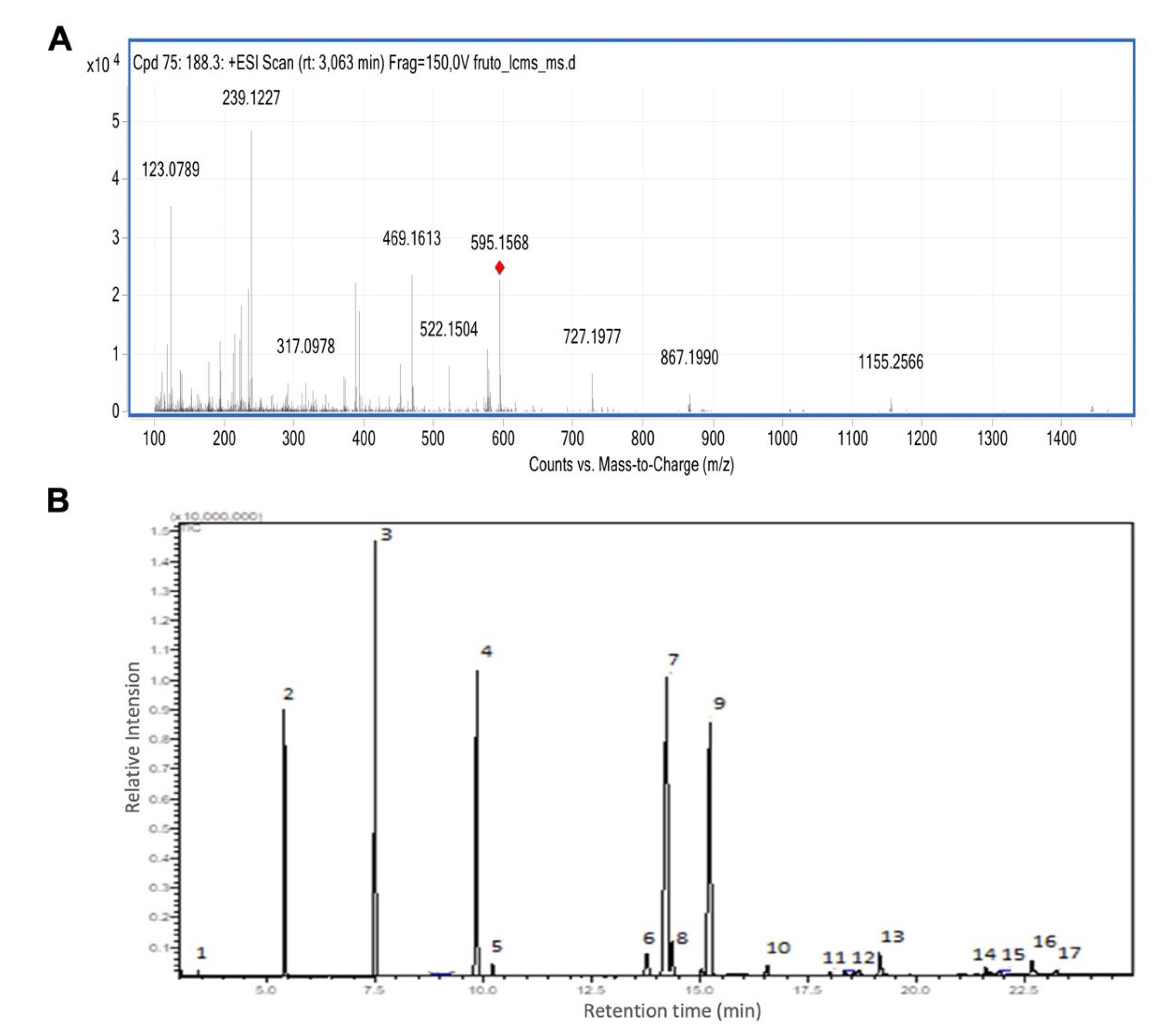
2.3. MS/MS Analysis
2.4. Oil Esterification
2.5. GC-MS Analysis
2.6. Cell Culture
2.7. Treatments with Euterpe Oleracea Mart Seed Extract and Oil
2.8. Cell Viability Test—MTT
2.9. Morphological Analysis via Inverted Light Microscopy
2.10. Annexin—V Assay
2.11. Western Blotting
2.12. Molecular Modeling Studies
2.12.1. Obtention of the 3D Structure of Annexin A2 and Construction of the Ligands
2.12.2. Docking Studies
2.12.3. Molecular Dynamics Simulations
2.13. Statistical Analysis
3. Results
3.1. Açaí Seed Extract Is Rich in Flavonoids
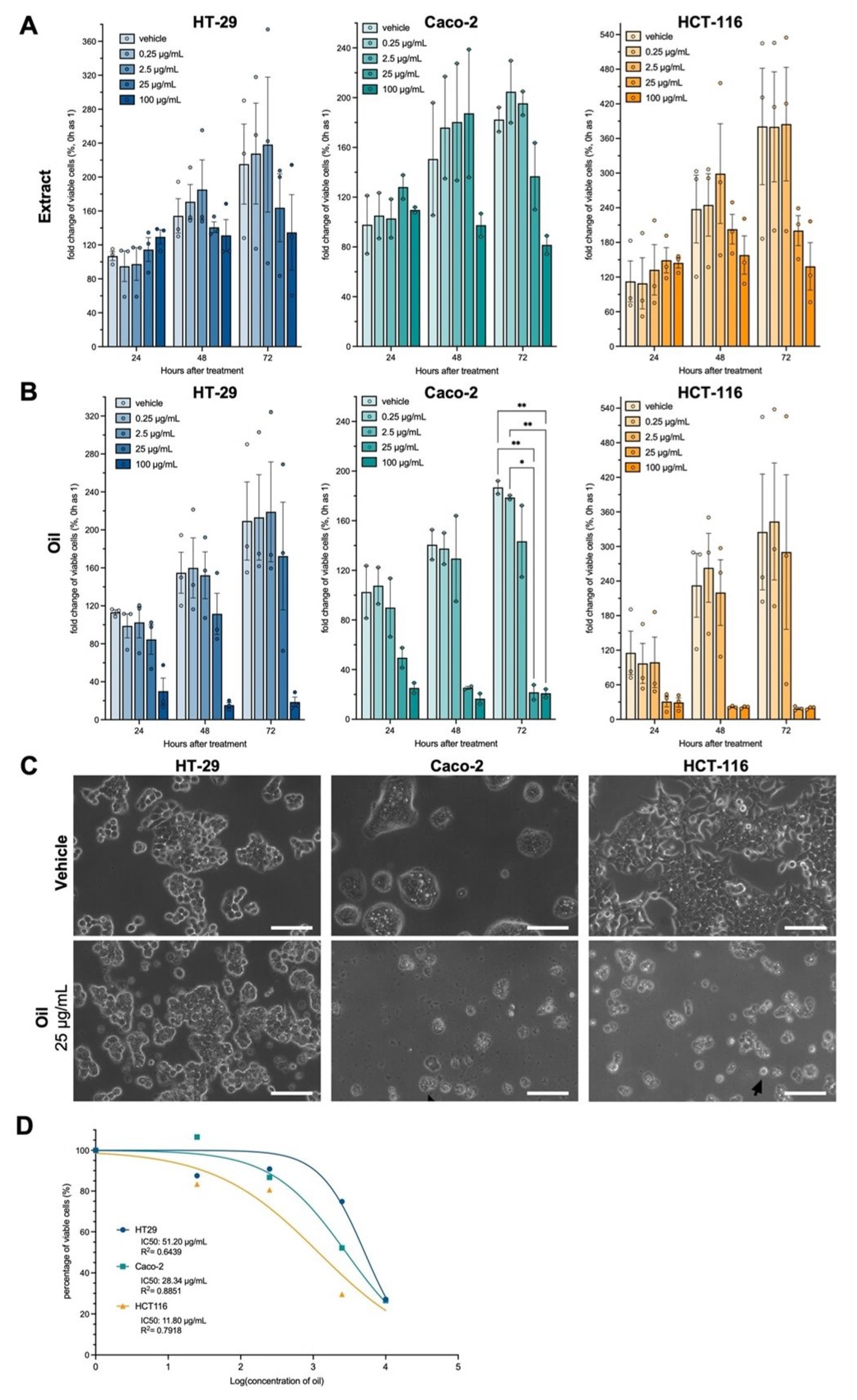
3.2. Açaí Seed Oil but Not Extract Exerts Cytotoxic Effect on Colorectal Cancer Cell Lines
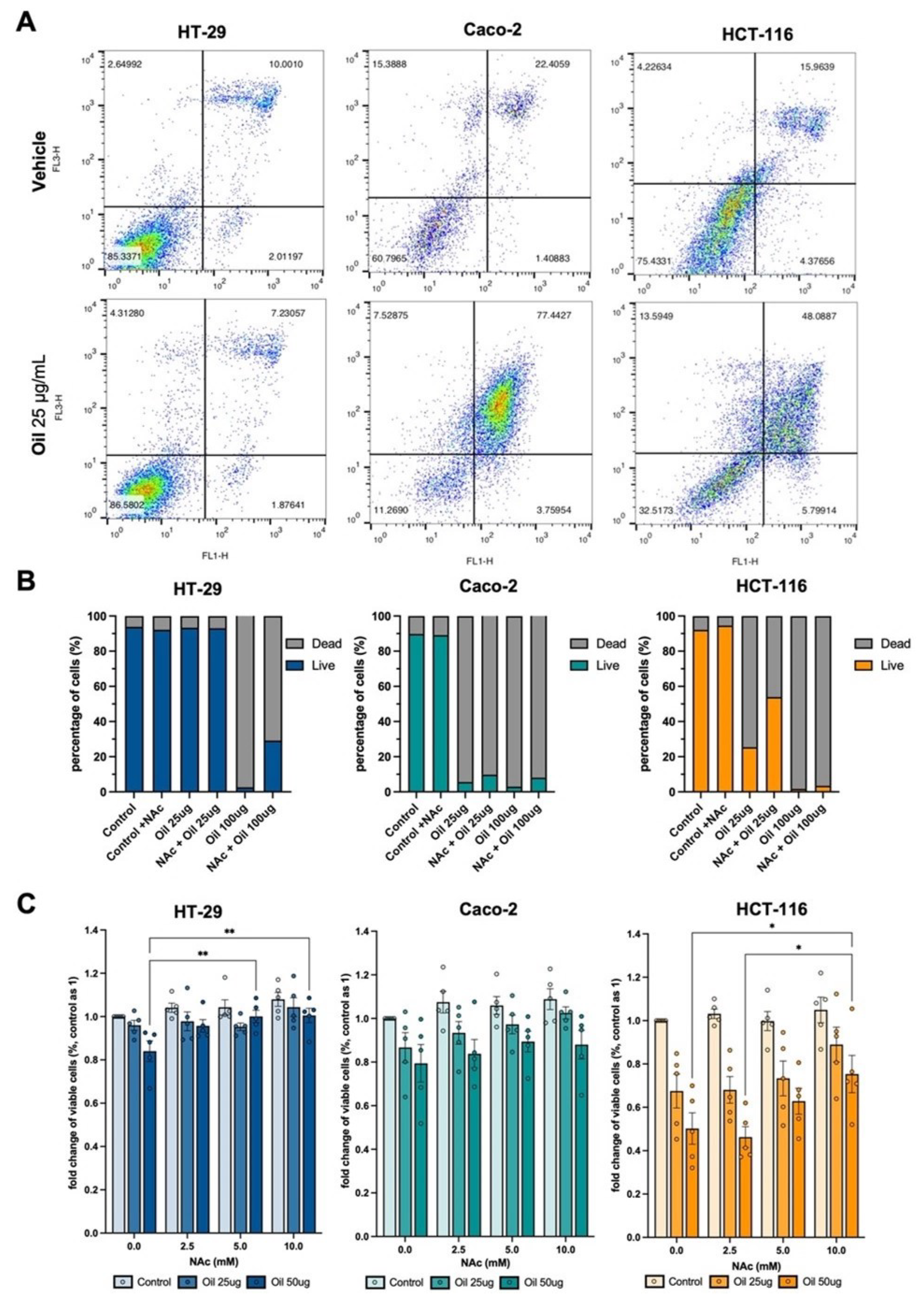
3.3. Açaí Seed Oil Induces Cellular Death through Increased ROS
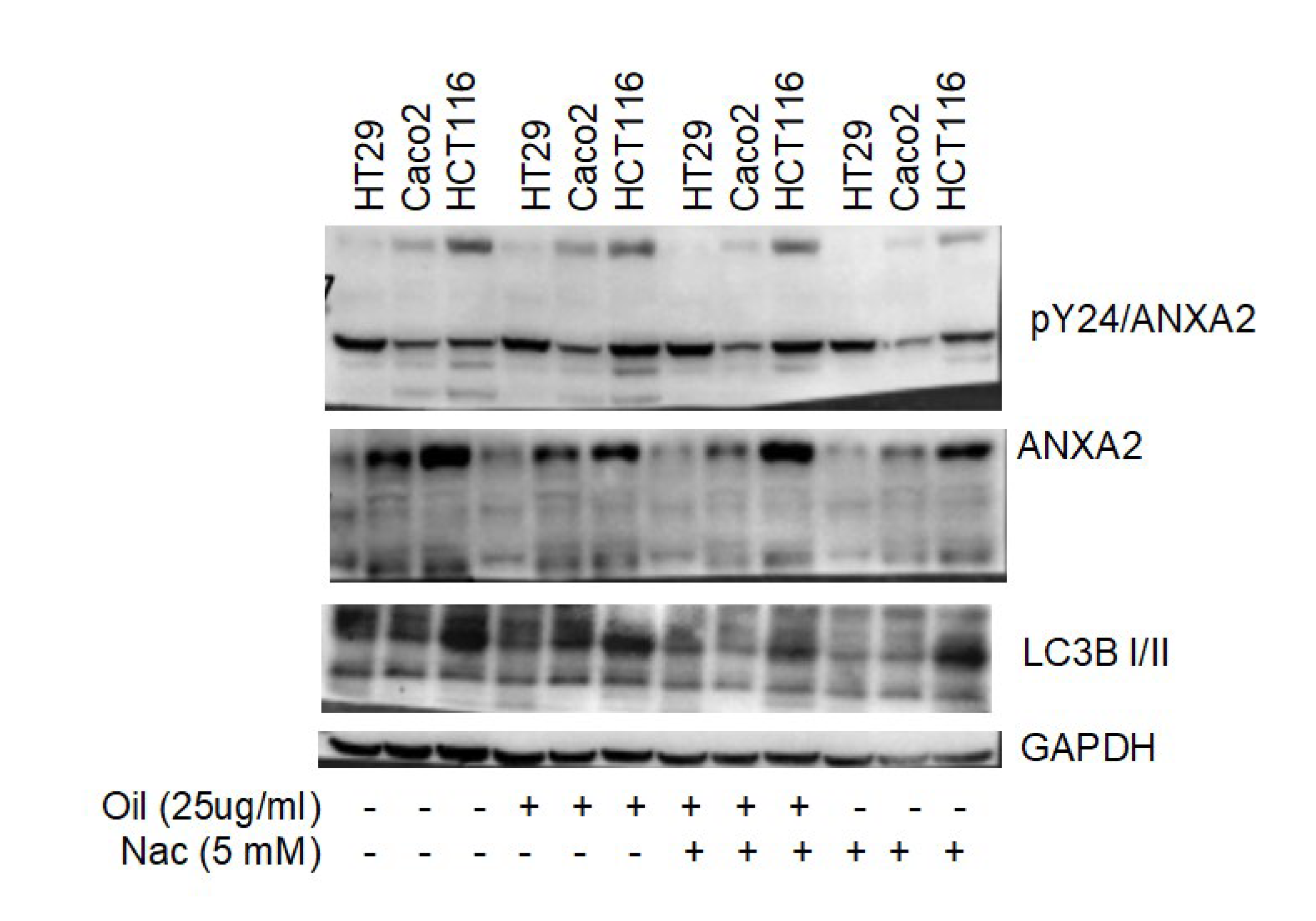
3.4. Autophagy and Annexin A2 Seem to Participate in Cellular Response to Açaí Seed Oil
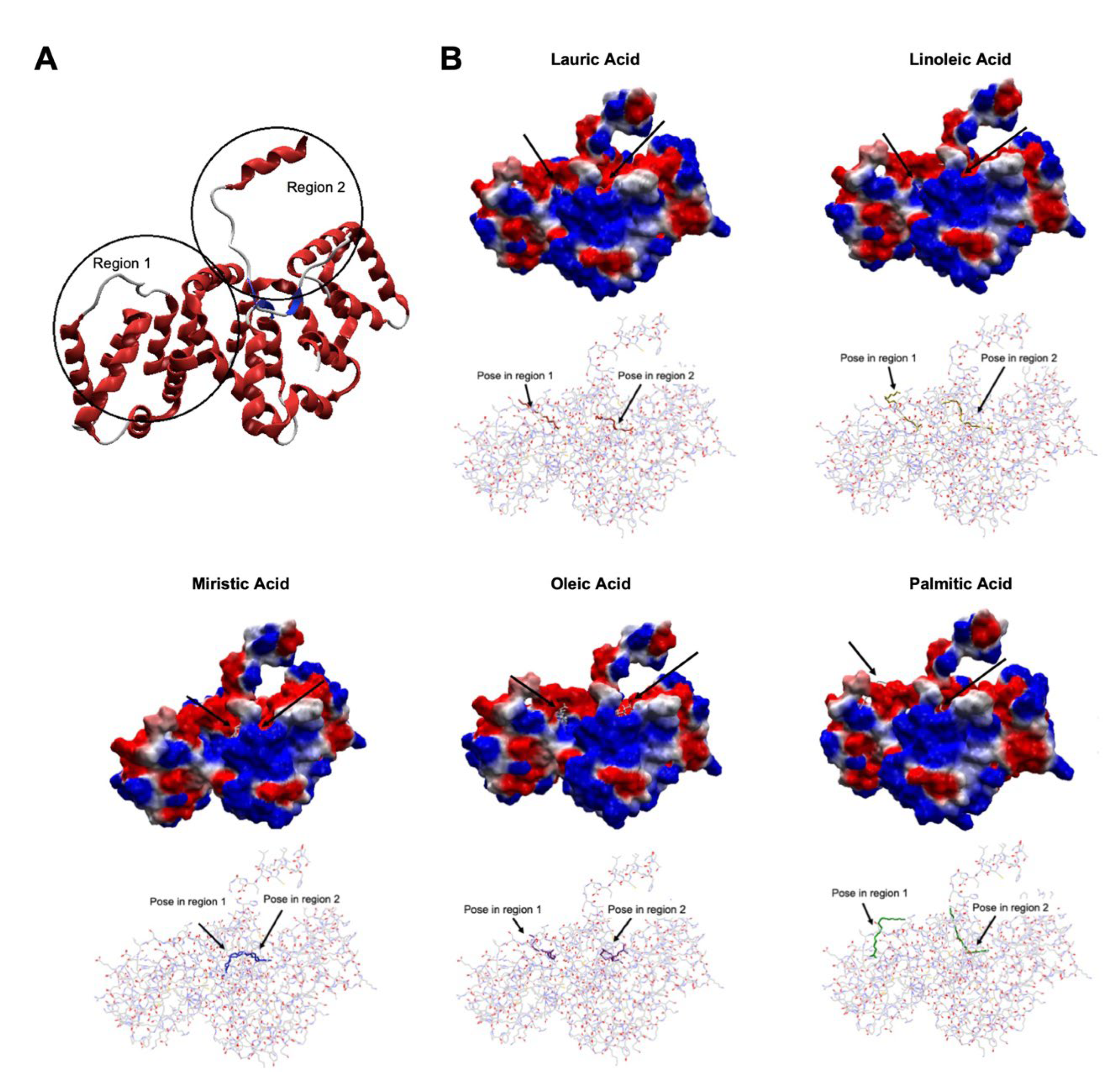
3.5. Docking
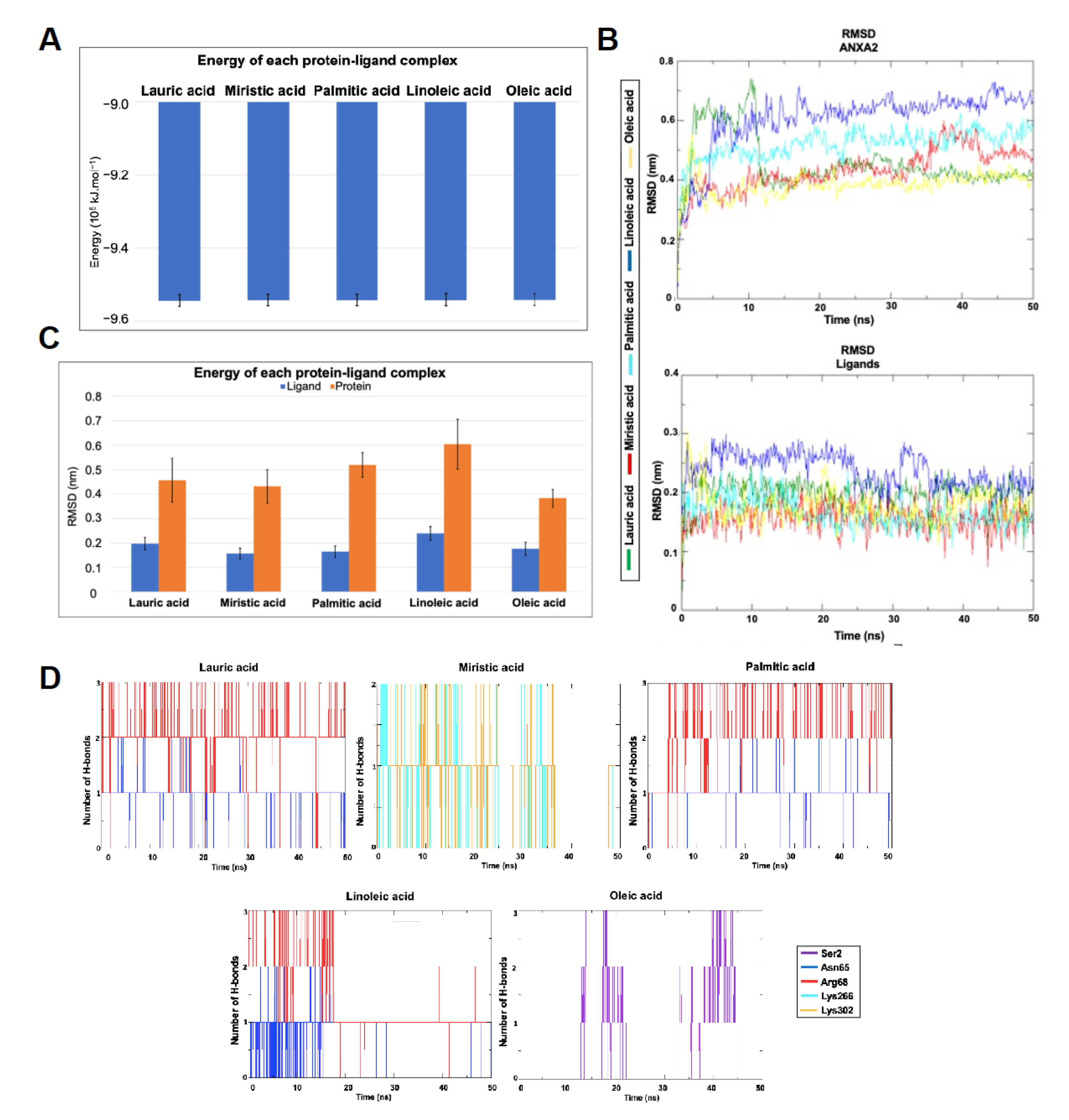
3.6. Molecular Dynamics Simulations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Brasil, Instituto Nacional de Câncer José Alencar Gomes da Silva. Estimativa 2023: Incidência de Câncer no Brasil; Instituto Nacional de Câncer Rio de Janeiro: Rio de Janeiro, Brazil, 2022; 160p, ISBN 978-65-88517-09-3. Available online: http://www.inca.gov.br (accessed on 10 January 2023).
- Empresa Brasileira de Pesquisa Agropecuária (Embrapa). Sistema de Produção do Açaí. 2006. Available online: http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Acai/SistemaProducaoAcai_2ed/index.htm (accessed on 8 August 2018).
- Gallori, S.; Bilia, A.R.; Bergonzi, M.C.; Barbosa, W.L.R.; Vincieri, F.F. Polyphenolic constituents of anthocyanins from the açaí fruit (Euterpe oleracea) Mart. Cienc. Tecnol. Aliment. 2004, 20, 388–390. [Google Scholar] [CrossRef]
- Rodrigues, R.B.; Lichtenthäler, R.; Zimmermann, B.F.; Papagiannopoulos, M.; Fabricius, H.; Marx, F.; Maia, J.G.S.; Almeida, O. Total Oxidant Scavenging Capacity of Euterpe oleracea Mart. (Açaí) Seeds and Identification of Their Polyphenolic Compounds. J. Agric. Food Chem. 2006, 54, 4162–4167. [Google Scholar] [CrossRef] [PubMed]
- Schauss, A.G.; Wu, X.; Prior, R.L.; Ou, B.; Patel, D.; Huang, D.; Kababick, J.P. Phytochemical and Nutrient Composition of the Freeze-Dried Amazonian Palm Berry, Euterpe oleraceae Mart. (Acai). J. Agric. Food Chem. 2006, 54, 8598–8603. [Google Scholar] [CrossRef]
- Chin, Y.-W.; Chai, H.-B.; Keller, W.J.; Kinghorn, A.D. Lignans and Other Constituents of the Fruits of Euterpe oleracea (Açai) with Antioxidant and Cytoprotective Activities. J. Agric. Food Chem. 2008, 56, 7759–7764. [Google Scholar] [CrossRef]
- Mantovani, I.S.B.; Fernandes, S.B.O.; Menezes, F.S. Constituintes apolares do fruto do açaí (Euterpe oleracea M.-Arecaceae). Rev. Bras. Farm. 2003, 13, 41–42. [Google Scholar] [CrossRef]
- Oliveira, M.S.P.; Carvalho, J.E.U.; Nascimento, W.M.O.; Müller, C.H. Cultivo do Açaizeiro Para Produção de Frutos; Ministério Da Agriculyura Pecuária E Abast: Brasilia, Brazil, 2002; ISSN 1517-211X. [Google Scholar]
- Mourão, L. História e natureza: Do açaí ao palmito. Rev. Territ. E Front. 2010, 3, 74–96. [Google Scholar] [CrossRef]
- Lee, R.; Balick, M.J. Palms, People, and Health. Explore 2008, 4, 59–62. [Google Scholar] [CrossRef]
- Goulding, M.; Smith, N. Palms: Sentinels for Amazon Conservation; Amazon Conservation Association & Missouri Botanical Garden: St. Louis, MO, USA, 2007; p. 356. [Google Scholar]
- Henderson, A. The Palms of the Amazon; University Press: Oxford, UK, 1995. [Google Scholar]
- Brian, M.B. Etnobotany of the Chacobo Indians and Their Palms Advanced in Economic Botany; The New York Botanic Garden: New York, NY, USA, 1988. [Google Scholar]
- Prance, G.H.; Silva, M.F. Árvores de Manaus; INPA: Manaus, Brazil, 1975; 312p. [Google Scholar]
- Kahn, F.; Granville, J. Palms in Forest Ecosystems of Amazonia, Ecological Studies No. 95; Springer: New York, NY, USA, 1992. [Google Scholar]
- Marinho, B.G.; Herdy, S.A.; Sá, A.C.; Santos, G.B.; Matheus, M.E.; Menezes, F.S.; Fernandes, P.D. Atividade antinociceptiva de extratos de açaí (Euterpe oleraceae Mart.). Rev. Bras. Farm. 2002, 12, 52–53. [Google Scholar] [CrossRef] [Green Version]
- Heitor, R.D.S.; Daniele, D.C.D.A.; Ariadna, L.P.; Hady, K.; Jesus, R.R.A.; José, C.T.C.; Da Silva, H.R.; Assis, D.D.C.D.; Prada, A.L.; Keita, H.; et al. Euterpe oleracea Mart. (aai): An old known plant with a new perspective. Afr. J. Pharm. Pharmacol. 2016, 10, 995–1006. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, H.; Roxo, M.; Krstin, S.; Wang, X.; Wink, M. Anthocyanin-rich extract of Acai (Euterpe precatoria Mart.) mediates neuroprotective activities in Caenorhabditis elegans. J. Funct. Foods 2016, 26, 385–393. [Google Scholar] [CrossRef]
- Romualdo, G.R.; Fragoso, M.F.; Borguini, R.G.; de Araújo Santiago, M.C.P.; Fernandes, A.A.H.; Barbisan, L.F. Protective effects of spray-dried açaí (Euterpe oleracea Mart.) fruit pulp against initiation step of colon carcinogenesis. Food Res. Int. 2015, 77, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Torma, P.D.C.M.R.; Brasil, A.V.S.; Carvalho, A.V.; Jablonski, A.; Rabelo, T.K.; Moreira, J.C.F.; Gelain, D.P.; Flôres, S.H.; Augusti, P.R.; Rios, A.D.O. Hydroethanolic extracts from different genotypes of açaí (Euterpe oleracea) presented antioxidant potential and protected human neuron-like cells (SH-SY5Y). Food Chem. 2017, 222, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.K.D.L.; Pereira, L.F.R.; Lamarão, C.V.; Lima, E.S.; da Veiga-Junior, V.F. Amazon acai: Chemistry and biological activities: A review. Food Chem. 2015, 179, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Melhorança Filho, A.L.; Pereira, M.R.R. Atividade antimicrobiana de óleos extraídos de açaí e de pupunha sobre o desenvolvimento de Pseudomonas aeruginosa e Staphylococcus aureus. Biosci. J. 2012, 28, 598–603. [Google Scholar]
- Magalhães, T.S.S.d.A.; Macedo, P.C.d.O.; Pacheco, S.Y.K.; da Silva, S.S.; Barbosa, E.G.; Pereira, R.R.; Costa, R.M.R.; Junior, J.O.C.S.; Ferreira, M.A.d.S.; de Almeida, J.C.; et al. Development and Evaluation of Antimicrobial and Modulatory Activity of Inclusion Complex of Euterpe oleracea Mart. Oil and β-Cyclodextrin or HP-β-Cyclodextrin. Int. J. Mol. Sci. 2020, 21, 942. [Google Scholar] [CrossRef] [Green Version]
- Favacho, H.A.S.; Oliveira, B.R.; Santos, K.C.; Medeiros, B.J.L.; Sousa, P.J.C.; Perazzo, F.F.; Carvalho, J.C.T. Anti-inflammatory and antinociceptive activities of Euterpe oleracea Mart., Arecaceae, oil. Rev. Bras. Farm. 2011, 21, 105–114. [Google Scholar] [CrossRef]
- Souza, B.S.F.; Carvalho, H.O.; Ferreira, I.M.; da Cunha, E.L.; Barros, A.S.; Taglialegna, T.; Carvalho, J.C. Effect of the treatment with Euterpe oleracea Mart. oil in rats with Triton-induced dyslipidemia. Biomed. Pharmacother. 2017, 90, 542–547. [Google Scholar] [CrossRef]
- Marques, E.S.; Tsuboy, M.S.F.; Carvalho, J.C.T.; Rosa, P.C.P.; Perazzo, F.F.; Gaivão, I.O.M.; Maistro, E.L. Research Article First cytotoxic, genotoxic, and antigenotoxic assessment of Euterpe oleracea fruit oil (açaí) in cultured human cells. Genet. Mol. Res. 2017, 16, gmr16039700. [Google Scholar] [CrossRef]
- Stoner, G.D.; Wang, L.-S.; Seguin, C.; Rocha, C.; Stoner, K.; Chiu, S.; Kinghorn, A.D. Multiple Berry Types Prevent N-Nitrosomethylbenzylamine-Induced Esophageal Cancer in Rats. Pharm. Res. 2010, 27, 1138–1145. [Google Scholar] [CrossRef] [Green Version]
- Fragoso, M.F.; Prado, M.G.; Barbosa, L.; Rocha, N.S.; Barbisan, L.F. Inhibition of Mouse Urinary Bladder Carcinogenesis by Açai Fruit (Euterpe oleraceae Martius) Intake. Plant Foods Hum. Nutr. 2012, 67, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, M.F.; Romualdo, G.R.; Ribeiro, D.A.; Barbisan, L.F. Açai (Euterpe oleracea Mart.) feeding attenuates dimethylhydrazine-induced rat colon carcinogenesis. Food Chem. Toxicol. 2013, 58, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragoso, M.F.; Romualo, G.R.; Vanderveer, L.A.; Franc-Barraza, J.; Cukierman, E.; Clapper, M.L.; Carvalho, R.F.; Barbisan, L.F. Lyophilized açaí pulp (Euterpe oleracea Mart.) attenuates colitis-associated colon carcinogenesis while its main an-thocyanin has the potential to affect the motility of colon cancer cells. Food Chem. Toxicol. 2018, 121, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Kim, N.; Nam, R.H.; Lee, S.; Lee, H.S.; Lee, H.-N.; Surh, Y.-J.; Lee, D.H. Açaí Berries Inhibit Colon Tumorigenesis in Azoxymethane/Dextran Sulfate Sodium-Treated Mice. Gut Liver 2017, 11, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.M.D.S.; Noratto, G.; Martino, H.S.D.; Arbizu, S.; Peluzio, M.D.C.G.; Talcott, S.; Ramos, A.M.; Mertens-Talcott, S.U. Pro-Apoptotic Activities of Polyphenolics from Açai (Euterpe oleracea Martius) in Human SW-480 Colon Cancer Cells. Nutr. Cancer 2014, 66, 1394–1405. [Google Scholar] [CrossRef]
- Monge-Fuentes, V.; Muehlmann, L.A.; Longo, J.P.F.; Silva, J.R.; Fascineli, M.L.; de Souza, P.E.N.; Faria, F.; Degterev, I.A.; Rodriguez, A.; Carneiro, F.P.; et al. Photodynamic therapy mediated by acai oil (Euterpe oleracea Martius) in nanoemulsion: A potential treatment for melanoma. J. Photochem. Photobiol. B Biol. 2017, 166, 301–310. [Google Scholar] [CrossRef]
- Alessandra-Perini, J.; Perini, J.A.; Rodrigues-Baptista, K.C.; de Moura, R.S.; Junior, A.P.; dos Santos, T.A.; Souza, P.J.C.; Nasciutti, L.E.; Machado, D.E. Euterpe oleracea extract inhibits tumorigenesis effect of the chemical carcinogen DMBA in breast experimental cancer. BMC Complement. Altern. Med. 2018, 18, 116. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.F.; Vidal, F.C.B.; Santos, D.; Costa, M.C.P.; Morgado-Díaz, J.A.; Nascimento, M.D.D.S.B.; De Moura, R.S. Cytotoxic effects of Euterpe oleracea Martius in malignant cell lines. BMC Complement. Altern. Med. 2014, 14, 175. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.A.C.N.; Costa, J.H.; Pacheco-Fill, T.; Ruiz, A.L.T.G.; Vidal, F.C.B.; Borges, K.R.A.; Guimarães, S.J.A.; de Azevedo-Santos, A.P.S.; Buglio, K.E.; Foglio, M.A.; et al. Açai (Euterpe oleracea Martius) Seed Extract Induces ROS Production and Cell Death in MCF-7 Breast Cancer Cell Line. Molecules 2021, 26, 3546. [Google Scholar] [CrossRef]
- Freitas, D.d.S.; Morgado-Díaz, J.; Gehren, A.S.; Vidal, F.C.B.; Fernandes, R.M.T.; Romão, W.; Tose, L.V.; Frazão, F.N.S.; Costa, M.C.P.; Silva, D.F.; et al. Cytotoxic analysis and chemical characterization of fractions of the hydroalcoholic extract of the Euterpe oleracea Martius seed in the MCF-7 cell line. J. Pharm. Pharmacol. 2017, 69, 714–721. [Google Scholar] [CrossRef]
- Chiang, Y.; Rizzino, A.; Sibenaller, Z.A.; Wold, M.S.; Vishwanatha, J.K. Specific down-regulation of Annexin II expres-sion in human cells interferes with cell proliferation. Mol. Cell. Biochem. 1999, 199, 139–147. [Google Scholar] [CrossRef]
- de Moura, R.S.; da Costa, G.F.; Moreira, A.S.B.; Queiroz, E.F.; Moreira, D.D.C.; Garcia-Souza, E.P.; Resende, C.; Moura, A.S.; Teixeira, M.T. Vitis vinifera L. grape skin extract activates the insulin-signalling cascade and reduces hyperglycaemia in alloxan-induced diabetic mice. J. Pharm. Pharmacol. 2011, 64, 268–276. [Google Scholar] [CrossRef]
- Hartman, L.; Lago, R.C. Rapid preparation of fatty acid methyl esters from lipids. Lab. Pract. 1973, 22, 475–476. [Google Scholar]
- De Albuquerque-Xavier, A.C.; Bastos, L.G.R.; De Freitas, J.C.M.; Leve, F.; De Souza, W.F.; De Araujo, W.M.; Wanderley, J.L.M.; Tanaka, M.N.; De Souza, W.; Morgado-Díaz, J.A. Blockade of irradiation-induced autophagosome formation impairs proliferation but does not enhance cell death in HCT-116 human colorectal carcinoma cells. Int. J. Oncol. 2012, 40, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.; Zhang, F.; Kemp, M.M.; Linhardt, R.J.; Waisman, D.M.; Head, J.H.; Seaton, B.A. Crystallographic Analysis of Calcium-dependent Heparin Binding to Annexin A2. J. Biol. Chem. 2006, 281, 31689–31695. [Google Scholar] [CrossRef] [PubMed]
- Hehre, W.J.; Deppmeier, B.J.; Klunzinger, P.E. PC Spartan Pro molecular modeling for desktop. Chem. Eng. News 1999, 77, 2. [Google Scholar]
- Reed, A.E.; Weinstock, R.B.; Weinhold, F. Natural population analysis. J. Chem. Phys. 1985, 83, 735–746. [Google Scholar] [CrossRef]
- Stewart, J.J.P. Optimization of parameters for semiempirical methods IV: Extension of MNDO, AM1, and PM3 to more main group elements. J. Mol. Model. 2004, 10, 155–164. [Google Scholar] [CrossRef]
- de Almeida, J.S.; Cavalcante, S.F.A.; Dolezal, R.; Kuca, K.; Musilek, K.; Jun, D.; França, T.C. Surface screening, molecular modeling and in vitro studies on the interactions of aflatoxin M1 and human enzymes acetyl- and butyrylcholinesterase. Chem. Interact. 2019, 308, 113–119. [Google Scholar] [CrossRef]
- Botelho, F.D.; dos Santos, M.C.; Gonçalves, A.d.S.; Kuca, K.; Valis, M.; LaPlante, S.R.; França, T.C.C.; de Almeida, J.S.F.D. Ligand-Based Virtual Screening, Molecular Docking, Molecular Dynamics, and MM-PBSA Calculations towards the Identification of Potential Novel Ricin Inhibitors. Toxins 2020, 12, 746. [Google Scholar] [CrossRef]
- Botelho, F.D.; Santos, M.C.; Gonçalves, A.S.; França, T.C.C.; LaPlante, S.R.; de Almeida, J.S.F.D. Identification of novel potential ricin inhibitors by virtual screening, molecular docking, molecular dynamics and MM-PBSA calculations: A drug repurposing approach. J. Biomol. Struct. Dyn. 2022, 40, 5309–5319. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; Van Der Spoel, D.; et al. GROMACS 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and Testing of the OPLS All-Atom Force Field on Conformational Energetics and Properties of Organic Liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Kaminski, G.A.; Friesner, R.A.; Tirado-Rives, J.; Jorgensen, W.L. Evaluation and Reparametrization of the OPLS-AA Force Field for Proteins via Comparison with Accurate Quantum Chemical Calculations on Peptides. J. Phys. Chem. B 2001, 105, 6474–6487. [Google Scholar] [CrossRef]
- da Silva, A.W.S.; Vranken, W.F. ACPYPE—AnteChamber PYthon Parser interface. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Turner, P.J. XMGRACE; Version 5.1.25; Center for Coastal and Land-Margin Research, Oregon Graduate Institute of Science and Technology: Beaverton, OR, USA, 2005. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Barth, S.; Glick, D.; Macleod, K.F. Autophagy: Assays and artifacts. J. Pathol. 2010, 221, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharadwaj, A.; Bydoun, M.; Holloway, R.; Waisman, D. Annexin A2 Heterotetramer: Structure and Function. Int. J. Mol. Sci. 2013, 14, 6259–6305. [Google Scholar] [CrossRef] [Green Version]
- Christensen, M.V.; Høgdall, C.K.; Jochumsen, K.M.; Høgdall, E.V. Annexin A2 and cancer: A systematic review. Int. J. Oncol. 2018, 52, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Madureira, P.A.; O’Connell, P.A.; Surette, A.P.; Miller, V.A.; Waisman, D.M. The Biochemistry and Regulation of S100A10: A Multifunctional Plasminogen Receptor Involved in Oncogenesis. J. Biomed. Biotechnol. 2012, 2012, 353687. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthäler, R.; Rodrigues, R.B.; Maia, J.G.S.; Papagiannopoulos, M.; Fabricius, H.; Marx, F. Total oxidant scavenging capacities of Euterpe oleracea Martius (Açaí) fruits. Int. J. Food Sci. Nutr. 2005, 56, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Palencia, L.A.; Duncan, C.E.; Talcott, S.T. Phytochemical composition and thermal stability of two commercial açai species, Euterpe oleracea and Euterpe precatoria. Food Chem. 2009, 115, 1199–1205. [Google Scholar] [CrossRef]
- Heinrich, M.; Dhanji, T.; Casselman, I. Açai (Euterpe oleracea Martius)—A phytochemical and pharmacological assessment of the species’ health claims. Phytochem. Lett. 2011, 4, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Mulabagal, V.; Calderón, A.I. Liquid chromatography/mass spectrometry based fingerprinting analysis and mass profiling of Euterpe oleracea (açaí) dietary supplement raw materials. Food Chem. 2012, 134, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, L.K.O.; Aguiar, J.P.L.; Filho, D.F.S.; Yuyama, K.; Varejão, M.D.J.; Favaro, D.I.T.; Vasconcellos, M.B.A.; Pimentel, S.A.; Caruso, M.S.F. Physicochemical characterization of acai juice of Euterpe precatoria Mart. from different Amazonian ecosystems. Acta Amaz. 2011, 41, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, R.J.S.D.; Couri, S.; Antoniassi, R.; Freitas, S.P. Composição em ácidos graxos do óleo da polpa de açaí extraído com enzimas e com hexano. Rev. Bras. De Frutic. 2008, 30, 498–502. [Google Scholar] [CrossRef]
- Zengin, G.; Mahomoodally, M.F.; Aktumsek, A.; Jekő, J.; Cziáky, Z.; Rodrigues, M.J.; Custodio, L.; Polat, R.; Cakilcioglu, U.; Ayna, A.; et al. Chemical Profiling and Biological Evaluation of Nepeta baytopii Extracts and Essential Oil: An Endemic Plant from Turkey. Plants 2021, 10, 1176. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylvestre, M.; Legault, J.; Dufour, D.; Pichette, A. Chemical composition and anticancer activity of leaf essential oil of Myrica gale L. Phytomedicine 2005, 12, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, J.; Dhumtanom, P.; Manosroi, A. Anti-proliferative activity of essential oil extracted from Thai medicinal plants on KB and P388 cell lines. Cancer Lett. 2006, 235, 114–120. [Google Scholar] [CrossRef]
- Wei, F.-X.; Li, M.-Y.; Song, Y.-H.; Li, H.-Z. Apoptosis and activity changes of telomerase induced by essential oil from pine needles in HepG2 cell line. J. Chin. Med. Mater. 2008, 31, 1197–1200. [Google Scholar]
- Jo, J.-R.; Park, J.S.; Park, Y.-K.; Chae, Y.Z.; Lee, G.-H.; Park, G.-Y.; Jang, B.-C. Pinus densiflora leaf essential oil induces apoptosis via ROS generation and activation of caspases in YD-8 human oral cancer cells. Int. J. Oncol. 2012, 40, 1238–1245. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Palencia, L.A.; Mertens-Talcott, S.; Talcott, S.T. Chemical Composition, Antioxidant Properties, and Thermal Stability of a Phytochemical Enriched Oil from Açai (Euterpe oleracea Martius). J. Agric. Food Chem. 2008, 56, 4631–4636. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Tan, J.; Miao, Y.; Lei, P.; Zhang, Q. ROS and Autophagy: Interactions and Molecular Regulatory Mechanisms. Cell. Mol. Neurobiol. 2015, 35, 615–621. [Google Scholar] [CrossRef]
- Maruyama, T.; Noda, N.N. Autophagy-regulating protease Atg4: Structure, function, regulation and inhibition. J. Antibiot. 2017, 71, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherz-Shouval, R.; Shvets, E.; Fass, E.; Shorer, H.; Gil, L.; Elazar, Z. Reactive oxygen species are essential for autophagy and specifically regulate the activity of Atg4. EMBO J. 2007, 26, 1749–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poillet-Perez, L.; Despouy, G.; Delage-Mourroux, R.; Boyer-Guittaut, M. Interplay between ROS and autophagy in cancer cells, from tumor initiation to cancer therapy. Redox Biol. 2015, 4, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerling, B.M.; Weinberg, F.; Snyder, C.; Burgess, Z.; Mutlu, G.M.; Viollet, B.; Budinger, G.S.; Chandel, N.S. Hypoxic activation of AMPK is dependent on mitochondrial ROS but independent of an increase in AMP/ATP ratio. Free. Radic. Biol. Med. 2009, 46, 1386–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spijkers-Hagelstein, J.A.P.; Pinhanços, S.M.; Schneider, P.; Pieters, R.; Stam, R.W. Src kinase-induced phosphorylation of Annexin A2 mediates glucocorticoid resistance in MLL-rearranged infant acute lymphoblastic leukemia. Leukemia 2012, 27, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnapillai, A.; Kolb, E.A.; Dhanan, P.; Mason, R.W.; Napper, A.; Barwe, S.P. Disruption of Annexin II/p11 Interaction Suppresses Leukemia Cell Binding, Homing and Engraftment, and Sensitizes the Leukemia Cells to Chemotherapy. PLoS ONE 2015, 10, e0140564. [Google Scholar] [CrossRef]
- Yang, J.; Cheng, T.-T.; Wang, L.-N.; Chen, W.; Li, X. The expression and clinical significance of Annexin II in clear-cell renal cell carcinoma. Chin. J. Cell. Mol. Immunol. 2012, 28, 59–62. [Google Scholar]
- Sharma, M.R.; Koltowski, L.; Ownbey, R.T.; Tuszynski, G.P.; Sharma, M.C. Angiogenesis-associated protein Annexin II in breast cancer: Selective expression in invasive breast cancer and contribution to tumor invasion and progression. Exp. Mol. Pathol. 2006, 81, 146–156. [Google Scholar] [CrossRef]
- Choi, C.H.; Chung, J.-Y.; Chung, E.J.; Sears, J.D.; Lee, J.-W.; Bae, D.-S.; Hewitt, S.M. Prognostic significance of Annexin A2 and Annexin A4 expression in patients with cervical cancer. BMC Cancer 2016, 16, 448. [Google Scholar] [CrossRef] [Green Version]
- Xiu, D.; Liu, L.; Qiao, F.; Yang, H.; Cui, L.; Liu, G. Annexin A2 Coordinates STAT3 to Regulate the Invasion and Migration of Colorectal Cancer Cells In Vitro. Gastroenterol. Res. Pract. 2016, 2016, 3521453. [Google Scholar] [CrossRef] [Green Version]
- Tristante, E.; Martínez, C.M.; Jiménez, S.; Mora, L.; Carballo, F.; Martínez-Lacaci, I.; de Torre-Minguela, C. Association of a characteristic membrane pattern of Annexin A2 with high invasiveness and nodal status in colon adenocarcinoma. Transl. Res. 2015, 166, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Gao, Y.; Li, X.; Cai, M.; Wang, H.; Zhuang, H.; Tan, M.; Liu, S.; Hao, Y.; Lin, B. Expression and clinical significance of Annexin A2 and human epididymis protein 4 in endometrial carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Yu, B.; Yan, Y.; Shen, J.; Zhao, S.; Zhu, J.; Qin, W.; Gao, Y. Correlation of expression levels of ANXA2, PGAM1, and CALR with glioma grade and prognosis. J. Neurosurg. 2013, 118, 846–853. [Google Scholar] [CrossRef]
- Masaki, T.; Mohammad, H.S.; Kurokohchi, K.; Yoneyama, H.; Tokuda, M.; Morishita, A.; Jian, G.; Shi, L.; Murota, M.; Tani, J.; et al. Annexin A2 expression and phosphorylation are up-regulated in hepatocellular carcinoma. Int. J. Oncol. 2008, 33, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-X.; Lv, H.; Li, Z.-X.; Li, C.; Wu, X.-Y. Effect of shRNA Mediated Down-Regulation of Annexin A2 on Biological Behavior of Human Lung Adencarcinoma Cells A549. Pathol. Oncol. Res. 2012, 18, 183–190. [Google Scholar] [CrossRef]
- Rocha, M.R.; Barcellos-De-Souza, P.; Sousa-Squiavinato, A.C.M.; Fernandes, P.V.; de Oliveira, I.M.; Boroni, M.; Morgado-Diaz, J.A. Annexin A2 overexpression associates with colorectal cancer invasiveness and TGF-ß induced epithelial mesenchymal transition via Src/ANXA2/STAT3. Sci. Rep. 2018, 8, 11285. [Google Scholar] [CrossRef] [Green Version]
- Emoto, K.; Yamada, Y.; Sawada, H.; Fujimoto, H.; Ueno, M.; Takayama, T.; Kamada, K.; Naito, A.; Hirao, S.; Nakajima, Y. Annexin II overexpression correlates with stromal tenascin-C overexpression: A prognostic marker in colorectal carcinoma. Cancer 2001, 92, 1419–1426. [Google Scholar] [CrossRef]
- Singh, P.; Wu, H.; Clark, C.; Owlia, A. Annexin II binds progastrin and gastrin-like peptides, and mediates growth factor effects of autocrine and exogenous gastrins on colon cancer and intestinal epithelial cells. Oncogene 2007, 26, 425–440. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Peaks | Carbon Chain | % Area— Retention Time (min) | Retention Time (min) |
|---|---|---|---|---|
| Capric acid | 1 | C 10:0 | 0.13 | 3.429 |
| Lauric acid | 2 | C 12:0 | 8.9 | 5.407 |
| Myristic acid | 3 | C 14:0 | 18.03 | 7.508 |
| Palmitic acid | 4 | C 16:0 | 16.61 | 9.845 |
| Palmitoleic acid | 5 | C 16:1 | 0.58 | 10.206 |
| Stearic acid | 6 | C 18:0 | 1.81 | 13.765 |
| Oleic acid | 7 | C 18:1 | 26.94 | 14.228 |
| Acid (isomer) | 8 | C 18:1 | 2.21 | 14.438 |
| Linoleic acid | 9 | C 18:2 | 19.92 | 15.223 |
| Linlenic acid | 10 | C 18:3 | 0.62 | 16.538 |
| Nanodecenoic acid | 11 | C 19:0 | 0.21 | 17.985 |
| Eicosenoic acid | 12 | C 20:0 | 0.35 | 18.326 |
| Non-esterified myristic acid | 13 | C 14:0 | 1.75 | 19.134 |
| Behenic acid | 14 | C 22:0 | 0.43 | 21.586 |
| Eicosatrienoic acid | 15 | C 23:0 | 0.22 | 21,904 |
| Non-esterified palmitic acid | 16 | C 16:0 | 0.88 | 22.649 |
| Tricosanotrienoic acid | 17 | C 23:0 | 0.22 | 23.212 |
| Ligand | Region | MolDock Score (kcal/mol) | Interacting Residues |
|---|---|---|---|
| Lauric acid | 1 | −99.42 | Arg179 |
| 2 | −84.21 | Asn65, Arg68, Ser296 | |
| Myristic acid | 1 | −94.63 | Asn65, Arg68 |
| 2 | −88.14 | Ser22, Lys302 | |
| Palmitic acid | 1 | −93.35 | Asp187, His224, Lys227 |
| 2 | −99.54 | Asn65 | |
| Oleic acid | 1 | −111.42 | Arg178 |
| 2 | −97.39 | Ala29 | |
| Linoleic acid | 1 | −105.94 | Arg179 |
| 2 | −112.26 | Asn65, Arg65, Ser296 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, M.A.C.N.; Tessmann, J.W.; Borges, K.R.A.; Wolff, L.A.S.; Botelho, F.D.; Vieira, L.A.; Morgado-Diaz, J.A.; Franca, T.C.C.; Barbosa, M.d.C.L.; Nascimento, M.d.D.S.B.; et al. Açaí (Euterpe oleracea Mart.) Seed Oil Exerts a Cytotoxic Role over Colorectal Cancer Cells: Insights of Annexin A2 Regulation and Molecular Modeling. Metabolites 2023, 13, 789. https://doi.org/10.3390/metabo13070789
da Silva MACN, Tessmann JW, Borges KRA, Wolff LAS, Botelho FD, Vieira LA, Morgado-Diaz JA, Franca TCC, Barbosa MdCL, Nascimento MdDSB, et al. Açaí (Euterpe oleracea Mart.) Seed Oil Exerts a Cytotoxic Role over Colorectal Cancer Cells: Insights of Annexin A2 Regulation and Molecular Modeling. Metabolites. 2023; 13(7):789. https://doi.org/10.3390/metabo13070789
Chicago/Turabian Styleda Silva, Marcos Antonio Custódio Neto, Josiane Weber Tessmann, Kátia Regina Assunção Borges, Laís Araújo Souza Wolff, Fernanda Diniz Botelho, Leandro Alegria Vieira, Jose Andres Morgado-Diaz, Tanos Celmar Costa Franca, Maria do Carmo Lacerda Barbosa, Maria do Desterro Soares Brandão Nascimento, and et al. 2023. "Açaí (Euterpe oleracea Mart.) Seed Oil Exerts a Cytotoxic Role over Colorectal Cancer Cells: Insights of Annexin A2 Regulation and Molecular Modeling" Metabolites 13, no. 7: 789. https://doi.org/10.3390/metabo13070789