
The Influence of Antioxidants on Oxidative Stress-Induced Vascular Aging in Obesity

 ,
,
Abstract
:1. Introduction
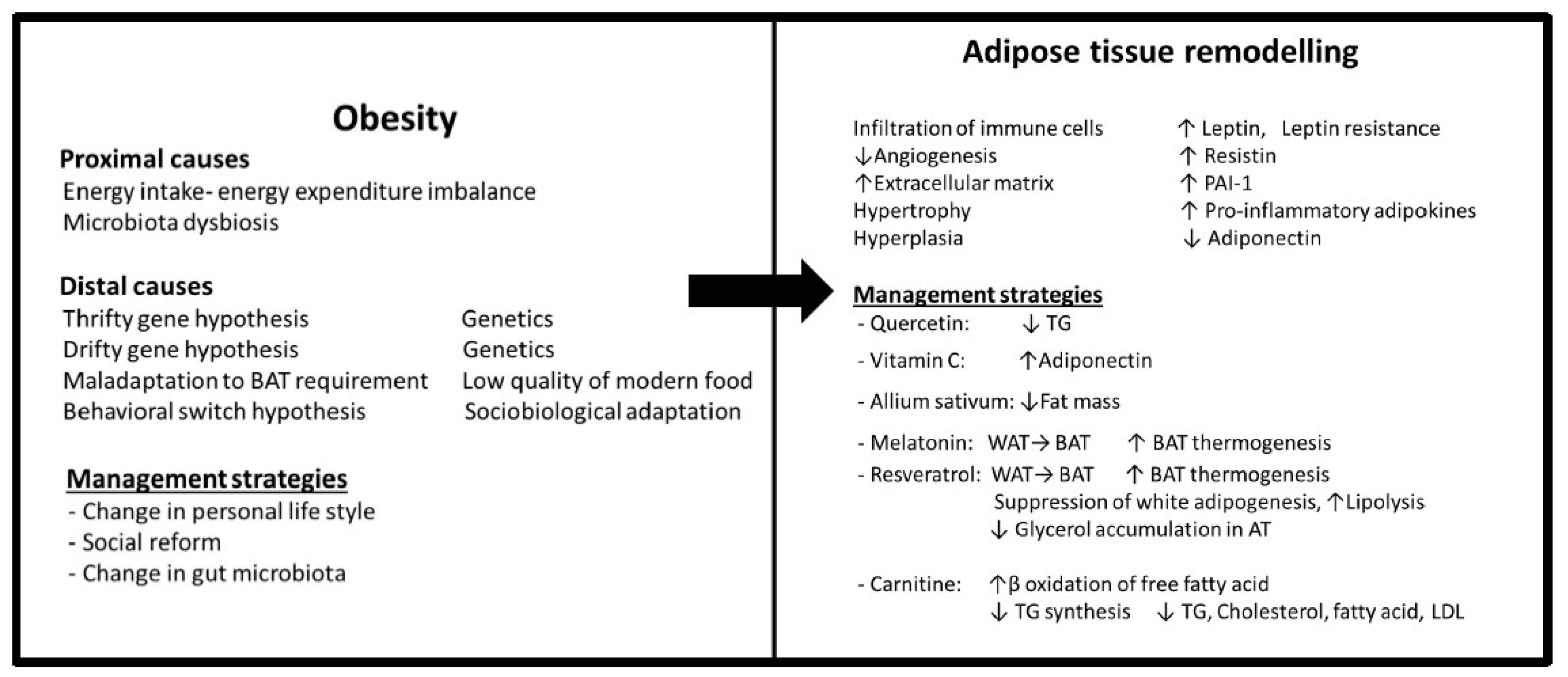
2. Obesity
2.1. Types and Functions of Adipose Tissue
2.2. Adipose Tissue Remodelling
3. Vascular Aging
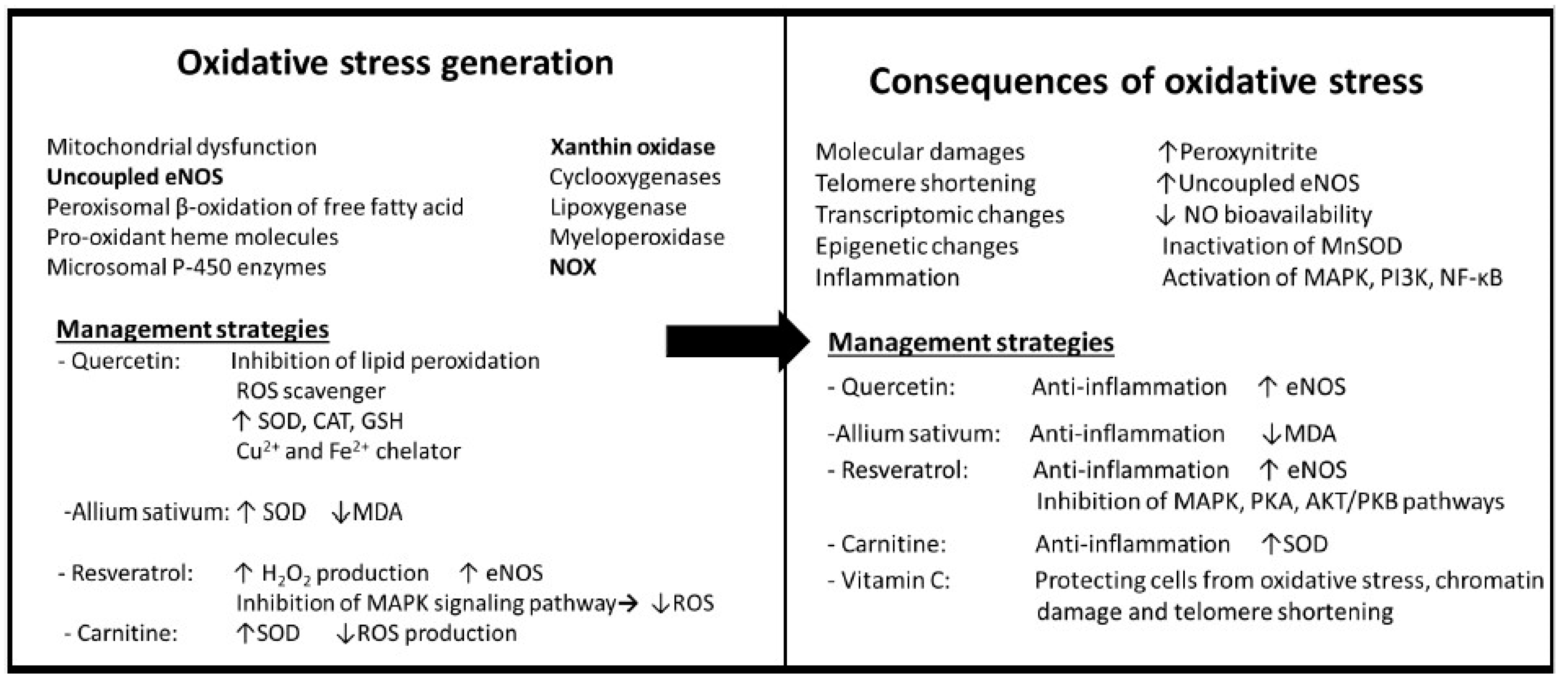
3.1. Oxidative Stress
Effects of Oxidative Stress
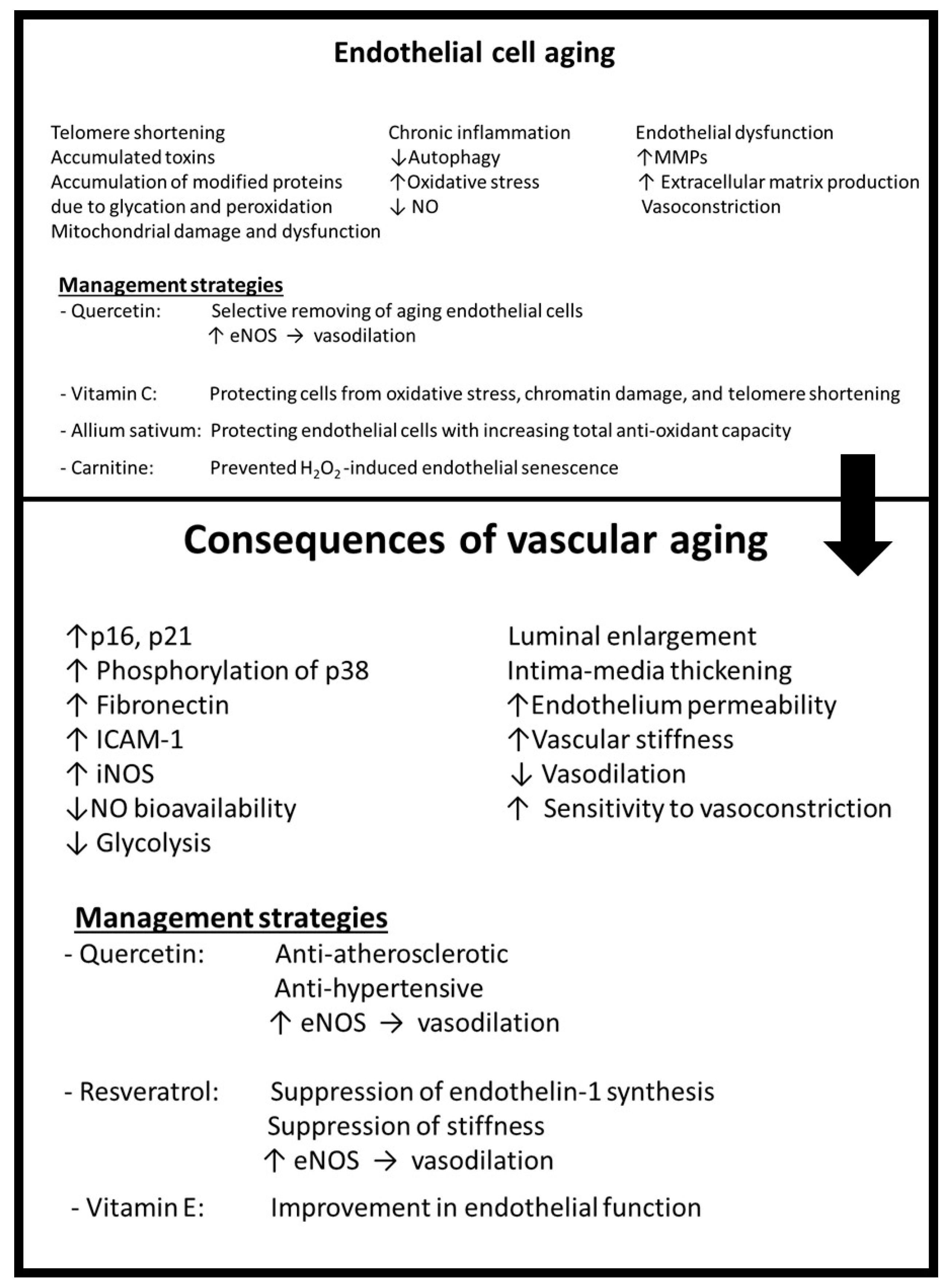
3.2. Mechanisms of Aging and Senescence
3.3. Consequences of Aging
4. Obesity and Microbiota Dysbiosis Role in Inflammation and Age-Related Vascular Dysfunction
5. Antioxidant Therapy
6. Possible Strategies—Are the Effects of Antioxidants or Other Strategies Greater as Prevention or Treatment?
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef] [PubMed]
- Stanek, A.; Grygiel-Górniak, B.; Brożyna-Tkaczyk, K.; Myśliński, W.; Cholewka, A.; Zolghadri, S. The Influence of Dietary Interventions on Arterial Stiffness in Overweight and Obese Subjects. Nutrients 2023, 15, 1440. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Jordan, J.; Nilsson, P.M.; Kotsis, V.; Olsen, M.H.; Grassi, G.; Yumuk, V.; Hauner, H.; Zahorska-Markiewicz, B.; Toplak, H.; Engeli, S.; et al. Joint scientific statement of the European Association for the Study of Obesity and the European Society of Hypertension: Obesity and early vascular ageing. J. Hypertens. 2015, 33, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hu, S.; Mao, Y. The mechanisms of vascular aging. Aging Med. 2021, 4, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Congdon, P.; Amugsi, D. Editorial: The obesity epidemic: Causes, context, prevention. Front. Public Health 2022, 10, 1030180. [Google Scholar] [CrossRef]
- Yang, M.; Liu, S.; Zhang, C. The Related Metabolic Diseases and Treatments of Obesity. Healthcare 2022, 10, 1616. [Google Scholar] [CrossRef]
- Jin, X.; Qiu, T.; Li, L.; Yu, R.; Chen, X. Pathophysiology of obesity and its associated diseases. Acta Pharm. Sin. B 2023, in press. [Google Scholar] [CrossRef]
- Xia, N.; Li, H. The role of perivascular adipose tissue in obesity-induced vascular dysfunction. Br. J. Pharmacol. 2017, 174, 3425–3442. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.W. Advances in Understanding of the Role of Lipid Metabolism in Aging. Cells 2021, 10, 880. [Google Scholar] [CrossRef]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Barton, M.; Baretella, O.; Meyer, M.R. Obesity and risk of vascular disease: Importance of endothelium-dependent vasoconstriction. Br. J. Pharmacol. 2012, 165, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Koenen, M.; Hill, M.A.; Cohen, P.; Sowers, J.R. Obesity, Adipose Tissue and Vascular Dysfunction. Circ. Res. 2021, 128, 951–968. [Google Scholar] [CrossRef] [PubMed]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüher, M. Adipose tissue dysfunction in obesity. Exp. Clin. Endocrinol. Diabetes 2009, 117, 241–250. [Google Scholar] [CrossRef]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.T.; Stanford, K.I. Batokines: Mediators of Inter-Tissue Communication (a Mini-Review). Curr. Obes. Rep. 2022, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, S.; Zolghadri, S.; Stanek, A. Beneficial Effects of Anti-Inflammatory Diet in Modulating Gut Microbiota and Controlling Obesity. Nutrients 2022, 14, 3985. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose tissue remodeling and obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, P.A.; James, M.E.; Goodwill, A.G.; Frisbee, J.C. Obesity and vascular dysfunction. Pathophysiology 2008, 15, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Bilgic Gazioglu, S.; Akan, G.; Atalar, F.; Erten, G. PAI-1 and TNF-α profiles of adipose tissue in obese cardiovascular disease patients. Int. J. Clin. Exp. Pathol. 2015, 8, 15919–15925. [Google Scholar]
- Alessi, M.C.; Poggi, M.; Juhan-Vague, I. Plasminogen activator inhibitor-1, adipose tissue and insulin resistance. Curr. Opin. Lipidol. 2007, 18, 240–245. [Google Scholar] [CrossRef]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, Obesity, and Leptin Resistance: Where Are We 25 Years Later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.S.; Qasim, A.; Reilly, M.P. Leptin resistance: A possible interface of inflammation and metabolism in obesity-related cardiovascular disease. J. Am. Coll. Cardiol. 2008, 52, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- Stanek, A.; Brożyna-Tkaczyk, K.; Myśliński, W. The Role of Obesity-Induced Perivascular Adipose Tissue (PVAT) Dysfunction in Vascular Homeostasis. Nutrients 2021, 13, 3843. [Google Scholar] [CrossRef]
- Dodig, S.; Čepelak, I.; Pavić, I. Hallmarks of senescence and aging. Biochem. Med. 2019, 29, 030501. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Liu, C.; Deng, C. Research progress on the mechanism of aging of vascular endothelial cells and the intervention of traditional Chinese medicine: A review. Medicine 2022, 101, e32248. [Google Scholar] [CrossRef] [PubMed]
- McDonald, R.B. Biology of Aging; Garland Science: New York, NY, USA, 2019. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Aroor, A.R.; Jia, C.; Sowers, J.R. Endothelial cell senescence in aging-related vascular dysfunction. Biochim. Biophys. Acta-Mol. Basis Dis. 2019, 1865, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y. Roles of Oxidative Stress and Inflammation in Vascular Endothelial Dysfunction-Related Disease. Antioxidants 2022, 11, 1958. [Google Scholar] [CrossRef] [PubMed]
- Sena, C.M.; Leandro, A.; Azul, L.; Seiça, R.; Perry, G. Vascular oxidative stress: Impact and therapeutic approaches. Front. Physiol. 2018, 9, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.J.; Kim, N.; Herman, A.B.; Gorospe, M.; Lee, J.S. Factors and Pathways Modulating Endothelial Cell Senescence in Vascular Aging. Int. J. Mol. Sci. 2022, 23, 10135. [Google Scholar] [CrossRef] [PubMed]
- Nedeljkovic, Z.S.; Gokce, N.; Loscalzo, J. Mechanisms of oxidative stress and vascular dysfunction. Postgrad. Med. J. 2003, 79, 195–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Kim, S.Y. Endothelial senescence in vascular diseases: Current understanding and future opportunities in senotherapeutics. Exp. Mol. Med. 2023, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Izzo, C.; Vitillo, P.; Di Pietro, P.; Visco, V.; Strianese, A.; Virtuoso, N.; Ciccarelli, M.; Galasso, G.; Carrizzo, A.; Vecchione, C. The Role of Oxidative Stress in Cardiovascular Aging and Cardiovascular Diseases. Life 2021, 11, 60. [Google Scholar] [CrossRef]
- Stanek, A.; Fazeli, B.; Bartuś, S.; Sutkowska, E. The Role of Endothelium in Physiological and Pathological States: New Data. Biomed. Res. Int. 2018, 2018, 1098039. [Google Scholar] [CrossRef] [Green Version]
- Brandes, R.P.; Fleming, I.; Busse, R. Endothelial aging. Cardiovasc. Res. 2005, 66, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Aman, Y.; Schmauck-medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N.; et al. Autophagy in healthy aging and disease. Nat. Aging 2021, 1, 634. [Google Scholar] [CrossRef]
- Donato, A.J.; Machin, D.R.; Lesniewski, L.A. Mechanisms of dysfunction in the aging vasculature and role in age-related disease. Circ. Res. 2018, 123, 825–848. [Google Scholar] [CrossRef]
- Starzak, M.; Stanek, A.; Jakubiak, G.K.; Cholewka, A.; Cieślar, G. Arterial Stiffness Assessment by Pulse Wave Velocity in Patients with Metabolic Syndrome and Its Components: Is It a Useful Tool in Clinical Practice? Int. J. Environ. Res. Public Health 2022, 19, 10368. [Google Scholar] [CrossRef]
- Keramat, S.; Karahan, O.; Patel, M.; Fazeli, B. Acute phase reactant proteins in Buerger’s disease: Is it a systemic disease? Vascular 2022, 31, 473–476. [Google Scholar] [CrossRef]
- Tabibzadeh, S. Role of autophagy in aging: The good, the bad, and the ugly. Aging Cell 2023, 22, e13753. [Google Scholar] [CrossRef]
- Fazeli, B.; Keramat, S.; Assadi, L.; Taheri, H. Angiogenesis induction in Buerger’s disease: A disease management double-edged sword? Orphanet. J. Rare Dis. 2019, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento-Andrade, Y.; Suárez, R.; Quintero, B.; Garrochamba, K.; Chapela, S.P. Gut microbiota and obesity: New insights. Front. Nutr. 2022, 9, 1018212. [Google Scholar] [CrossRef] [PubMed]
- Vetrani, C.; Di Nisio, A.; Paschou, S.A.; Barrea, L.; Muscogiuri, G.; Graziadio, C.; Savastano, S.; Colao, A.; On Behalf of the Obesity Programs of Nutrition Education Research and Assessment Opera Group. From Gut Microbiota through Low-Grade Inflammation to Obesity: Key Players and Potential Targets. Nutrients 2022, 14, 2103. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.N.; Liu, X.T.; Liang, Z.H.; Wang, J.H. Gut microbiota in obesity. World J. Gastroenterol. 2021, 27, 3837–3850. [Google Scholar] [CrossRef]
- Brunt, V.E.; Gioscia-Ryan, R.A.; Richey, J.J.; Zigler, M.C.; Cuevas, L.M.; Gonzalez, A.; Vázquez-Baeza, Y.; Battson, M.L.; Smithson, A.T.; Gilley, A.D.; et al. Suppression of the gut microbiome ameliorates age-related arterial dysfunction and oxidative stress in mice. J. Physiol. 2019, 597, 2361–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The links between gut microbiota and obesity and obesity related diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef] [PubMed]
- Agnoletti, D.; Piani, F.; Cicero, A.F.G.; Borghi, C. The Gut Microbiota and Vascular Aging: A State-of-the-Art and Systematic Review of the Literature. J. Clin. Med. 2022, 11, 3557. [Google Scholar] [CrossRef] [PubMed]
- Aoun, A.; Darwish, F.; Hamod, N. The Influence of the Gut Microbiome on Obesity in Adults and the Role of Probiotics, Prebiotics, and Synbiotics for Weight Loss. Prev. Nutr. Food Sci. 2020, 25, 113–123. [Google Scholar] [CrossRef]
- Van Hul, M.; Cani, P.D. The gut microbiota in obesity and weight management: Microbes as friends or foe? Nat. Rev. Endocrinol. 2023, 19, 258–271. [Google Scholar] [CrossRef]
- Keramat, S.; Sharebiani, H.; Patel, M.; Fazeli, B.; Stanek, A. The Potential Role of Antioxidants in the Treatment of Peripheral Arterial Disease: A Systematic Review. Antioxidants 2022, 11, 2126. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Yao, L.; Gu, W.; Zhao, S.; Shen, Z.; Lin, Z.; Liu, W.; Yan, T. Pharmacological Activity of Quercetin: An Updated Review. Evid.-Based Complement. Altern. Med. 2022, 2022, 3997190. [Google Scholar] [CrossRef]
- Dagher, O.; Mury, P.; Thorin-Trescases, N.; Noly, P.E.; Thorin, E.; Carrier, M. Therapeutic Potential of Quercetin to Alleviate Endothelial Dysfunction in Age-Related Cardiovascular Diseases. Front. Cardiovasc. Med. 2021, 8, 658400. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Jiang, L.Y.; Wang, Y.C.; Ma, D.F.; Li, X. Quercetin Attenuates Atherosclerosis via Modulating Oxidized LDL-Induced Endothelial Cellular Senescence. Front. Pharmacol. 2020, 11, 512. [Google Scholar] [CrossRef]
- Pei, Y.; Parks, J.S.; Kang, H.W. Quercetin alleviates high-fat diet-induced inflammation in brown adipose tissue. J. Funct. Foods 2021, 85, 104614. [Google Scholar] [CrossRef]
- Kábelová, A.; Malínská, H.; Marková, I.; Hűttl, M.; Chylíková, B.; Šeda, O. Quercetin supplementation alters adipose tissue and hepatic transcriptomes and ameliorates adiposity, dyslipidemia, and glucose intolerance in adult male rats. Front. Nutr. 2022, 9, 952065. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S.; Rahmat, A.; Patimah, I.; Khaza’ai, H.; Abed, Y. Effect of vitamin C on inflammation and metabolic markers in hypertensive and/or diabetic obese adults: A randomized controlled trial. Drug Des. Dev. Ther. 2015, 9, 3405–3412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Ng, C.; He, J.; Yang, M.; Luo, X.; Herbert, T.P.; Whitehead, J.P. Vitamin C protects against hypoxia, inflammation, and ER stress in primary human preadipocytes and adipocytes. Mol. Cell. Endocrinol. 2022, 556, 111740. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, S.; Ali, S.; Tahir, H.M.; Kazmi, S.A.R.; Shakir, H.A.; Mughal, T.A.; Mumtaz, S.; Summer, M.; Farooq, M.A. Aging and its treatment with vitamin C: A comprehensive mechanistic review. Mol. Biol. Rep. 2021, 48, 8141–8153. [Google Scholar] [CrossRef] [PubMed]
- Ashor, A.W.; Siervo, M.; Lara, J.; Oggioni, C.; Afshar, S.; Mathers, J.C. Effect of vitamin C and vitamin e supplementation on endothelial function: A systematic review and meta-analysis of randomised controlled trials. Br. J. Nutr. 2015, 113, 1182–1194. [Google Scholar] [CrossRef] [Green Version]
- Alcalá, M.; Sánchez-Vera, I.; Sevillano, J.; Herrero, L.; Serra, D.; Ramos, M.P.; Viana, M. Vitamin E reduces adipose tissue fibrosis, inflammation, and oxidative stress and improves metabolic profile in obesity. Obesity 2015, 23, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and anti-inflammatory effects of garlic compounds. J. Immunol. Res. 2015, 2015, 401630. [Google Scholar] [CrossRef] [Green Version]
- Ogunyemi, B.C.; Osaretin, A.T.E.; Oshanupin Adeola, C.; Ogunyemi, A.K. Effect of Allium sativum extract in combination-with orlistat on insulin resistance and disrupted metabolic hormones in high fat diet induced obese rats. Sci. Afr. 2021, 14, e00994. [Google Scholar] [CrossRef]
- Hou, C.Y.; Tain, Y.L.; Yu, H.R.; Huang, L.T. The Effects of Resveratrol in the Treatment of Metabolic Syndrome. Int. J. Mol. Sci. 2019, 20, 535. [Google Scholar] [CrossRef] [Green Version]
- Konyalioglu, S.; Armagan, G.; Yalcin, A.; Atalayin, C.; Dagci, T. Effects of resveratrol on hydrogen peroxide-induced oxidative stress in embryonic neural stem cells. Neural Regen. Res. 2013, 8, 485–495. [Google Scholar] [CrossRef]
- Shimojima, Y.; Kishida, D.; Ichikawa, T.; Takamatsu, R.; Nomura, S.; Sekijima, Y. Oxidative Stress Promotes Instability of Regulatory T Cells in Antineutrophil Cytoplasmic Antibody-Associated Vasculitis. Front. Immunol. 2021, 12, 789740. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, J.; Li, X.; Zhou, Q.; Bai, J.; Shi, Y.; Le, G. Resveratrol prevents suppression of regulatory T-cell production, oxidative stress, and inflammation of mice prone or resistant to high-fat diet-induced obesity. Nutr. Res. 2013, 33, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.A.; Franco, F.N.; Caldeira, C.A.; de Araújo, G.R.; Vieira, A.; Chaves, M.M.; Lara, R.C. Antioxidant effect of Resveratrol: Change in MAPK cell signaling pathway during the aging process. Arch. Gerontol. Geriatr. 2021, 92, 104266. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Shang, X.; Wu, H.; Gautam, S.C.; Al-Holou, S.; Li, C.; Kuo, J.; Zhang, L.; Chopp, M. Resveratrol downregulates PI3K/Akt/mTOR signaling pathways in human U251 glioma cells. J. Exp. Ther. Oncol. 2009, 8, 25–33. [Google Scholar] [PubMed]
- Andrade, J.M.; Frade, A.C.; Guimarães, J.B.; Freitas, K.M.; Lopes, M.T.; Guimarães, A.L.; de Paula, A.M.; Coimbra, C.C.; Santos, S.H. Resveratrol increases brown adipose tissue thermogenesis markers by increasing SIRT1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur. J. Nutr. 2014, 53, 1503–1510. [Google Scholar] [CrossRef]
- Kim, O.Y.; Chung, J.Y.; Song, J. Effect of resveratrol on adipokines and myokines involved in fat browning: Perspectives in healthy weight against obesity. Pharmacol. Res. 2019, 148, 104411. [Google Scholar] [CrossRef]
- Li, H.; Xia, N.; Hasselwander, S.; Daiber, A. Resveratrol and Vascular Function. Int. J. Mol. Sci. 2019, 20, 2155. [Google Scholar] [CrossRef] [Green Version]
- Rebouche, C.J. Carnitine. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Tucker, K.L., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 252–253. [Google Scholar]
- de Sotomayor, M.A.; Mingorance, C.; Rodriguez-Rodriguez, R.; Marhuenda, E.; Herrera, M.D. l-carnitine and its propionate: Improvement of endothelial function in SHR through superoxide dismutase-dependent mechanisms. Free Radic. Res. 2007, 41, 884–891. [Google Scholar] [CrossRef]
- Virmani, M.A.; Cirulli, M. The Role of l-Carnitine in Mitochondria, Prevention of Metabolic Inflexibility and Disease Initiation. Int. J. Mol. Sci. 2022, 23, 2717. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Chou, H.C.; Chen, Y.T.; Tung, S.Y.; Ko, T.L.; Buyandelger, B.; Wen, L.L.; Juan, S.H. L-Carnitine reduces reactive oxygen species/endoplasmic reticulum stress and maintains mitochondrial function during autophagy-mediated cell apoptosis in perfluorooctanesulfonate-treated renal tubular cells. Sci. Rep. 2022, 12, 4673. [Google Scholar] [CrossRef]
- Wang, Q.; Ju, X.; Chen, Y.; Dong, X.; Luo, S.; Liu, H.; Zhang, D. L-carnitine exerts a cytoprotective effect against H2O2 -induced oxidative stress in the fathead minnow muscle cell line. Aquac. Res. 2017, 48, 941–954. [Google Scholar] [CrossRef] [Green Version]
- Keshani, M.; Alikiaii, B.; Askari, G.; Yahyapoor, F.; Ferns, G.A.; Bagherniya, M. The effects of L-carnitine supplementation on inflammatory factors, oxidative stress, and clinical outcomes in patients with sepsis admitted to the intensive care unit (ICU): Study protocol for a double blind, randomized, placebo-controlled clinical trial. Trials 2022, 23, 170. [Google Scholar] [CrossRef]
- Abbasnezhad, A.; Hasanavand, A.; Falahi, E.; Kashkooli, S.; Asbaghi, O.; Choghakhori, R. Effect of L-Carnitine Supplementation on Lipid Profiles of Patients with Liver Disease: A Systematic Review and Meta-Analysis. Prev. Nutr. Food Sci. 2020, 25, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R. Evolutionary perspectives on the obesity epidemic: Adaptive, maladaptive, and neutral viewpoints. Annu. Rev. Nutr. 2013, 33, 289–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genné-Bacon, E.A. Thinking evolutionarily about obesity. Yale J. Biol. Med. 2014, 87, 99–112. [Google Scholar]
- Benzie, I.F.F. Evolution of dietary antioxidants. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 136, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Taherkhani, S.; Suzuki, K.; Ruhee, R.T. A brief overview of oxidative stress in adipose tissue with a therapeutic approach to taking antioxidant supplements. Antioxidants 2021, 10, 594. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Choudhury, S.T.; Seidel, V.; Rahman, A.B.; Aziz, M.A.; Richi, A.E.; Rahman, A.; Jafrin, U.H.; Hannan, J.M.A.; Abdel-Wahab, Y.H.A. Therapeutic Potential of Quercetin in the Management of Type-2 Diabetes Mellitus. Life 2022, 12, 1146. [Google Scholar] [CrossRef]
- Pei, Y.; Otieno, D.; Gu, I.; Lee, S.O.; Parks, J.S.; Schimmel, K.; Kang, H.W. Effect of quercetin on nonshivering thermogenesis of brown adipose tissue in high-fat diet-induced obese mice. J. Nutr. Biochem. 2021, 88, 108532. [Google Scholar] [CrossRef]
- Forney, L.A.; Lenard, N.R.; Stewart, L.K.; Henagan, T.M. Dietary quercetin attenuates adipose tissue expansion and inflammation and alters adipocyte morphology in a tissue-specific manner. Int. J. Mol. Sci. 2018, 19, 895. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Mukai, Y. Modulation of Chronic Inflammation by Quercetin: The Beneficial Effects on Obesity. J. Inflamm. Res. 2020, 13, 421–431. [Google Scholar] [CrossRef]
- Kuipers, E.N.; van Dam, A.D.; Held, N.M.; Mol, I.M.; Houtkooper, R.H.; Rensen, P.C.N.; Boon, M.R. Quercetin lowers plasma triglycerides accompanied by white adipose tissue browning in diet-induced obese mice. Int. J. Mol. Sci. 2018, 19, 1786. [Google Scholar] [CrossRef] [Green Version]
- Rajizadeh, M.A.; Bejeshk, M.A.; Doustimotlagh, A.H.; Najafipour, H.; Eftekhari, M.; Mahmoodi, M.; Azizi, M.; Rostamabadi, F.; Pourghadamyari, H. The Alleviating Impacts of Quercetin on Inflammation and Oxidant-antioxidant Imbalance in Rats with Allergic Asthma. Iran. J. Allergy Asthma Immunol. 2023, 22, 138–149. [Google Scholar] [CrossRef]
- Aouichat, S.; Raya, E.; Molina-Carballo, A.; Munoz-Hoyos, A.; Aloweidi, A.S.; Elmahallawy, E.K.; Agil, A. Dose-Dependent Effect of Melatonin on BAT Thermogenesis in Zücker Diabetic Fatty Rat: Future Clinical Implications for Obesity. Antioxidants 2022, 11, 1646. [Google Scholar] [CrossRef] [PubMed]
- Man, A.W.C.; Xia, N.; Li, H. Circadian rhythm in adipose tissue: Novel antioxidant target for metabolic and cardiovascular diseases. Antioxidants 2020, 9, 968. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Bal, N.B.; Sadi, G.; Usanmaz, S.E.; Uludag, M.O.; Demirel-Yilmaz, E. The effects of resveratrol and exercise on age and gender-dependent alterations of vascular functions and biomarkers. Exp. Gerontol. 2018, 110, 191–201. [Google Scholar] [CrossRef]
- Liu, D.; Zeng, X.; Li, L.; Ou, Z.L. Carnitine promotes recovery from oxidative stress and extends lifespan in C. elegans. Aging 2020, 13, 813–830. [Google Scholar] [CrossRef]
- Plantinga, Y.; Ghiadoni, L.; Magagna, A.; Giannarelli, C.; Franzoni, F.; Taddei, S.; Salvetti, A. Supplementation with vitamins C and E improves arterial stiffness and endothelial function in essential hypertensive patients. Am. J. Hypertens. 2007, 20, 392–397. [Google Scholar] [CrossRef]
- Lee, C.Y. Effects of dietary vitamins on obesity-related metabolic parameters. J. Nutr. Sci. 2023, 12, e47. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lim, S.M.; Yoo, J.A.; Woo, M.J.; Cho, K.H. Consumption of high-dose vitamin C (1250 mg per day) enhances functional and structural properties of serum lipoprotein to improve anti-oxidant, anti-atherosclerotic, and anti-aging effects via regulation of anti-inflammatory microRNA. Food Funct. 2015, 6, 3604–3612. [Google Scholar] [CrossRef]
- Costa, R.M.; Neves, K.B.; Tostes, R.C.; Lobato, N.S. Perivascular adipose tissue as a relevant fat depot for cardiovascular risk in obesity. Front. Physiol. 2018, 9, 253. [Google Scholar] [CrossRef]
- Man, A.W.C.; Zhou, Y.; Xia, N.; Li, H. Perivascular adipose tissue as a target for antioxidant therapy for cardiovascular complications. Antioxidants 2020, 9, 574. [Google Scholar] [CrossRef]
- Siauciunaite, R.; Foulkes, N.S.; Calabrò, V.; Vallone, D. Evolution Shapes the Gene Expression Response to Oxidative Stress. Int. J. Mol. Sci. 2019, 20, 3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neba Ambe, G.N.N.; Breda, C.; Bhambra, A.S.; Arroo, R.R.J. Effect of the Citrus Flavone Nobiletin on Circadian Rhythms and Metabolic Syndrome. Molecules 2022, 27, 7727. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, J.; Yao, K.; Su, W.; Tan, B.; Wu, X.; Huang, X.; Li, T.; Yin, Y.; Tosini, G.; et al. Circadian rhythms and obesity: Timekeeping governs lipid metabolism. J. Pineal Res. 2020, 69, e12682. [Google Scholar] [CrossRef] [PubMed]
- Wilking, M.; Ndiaye, M.; Mukhtar, H.; Ahmad, N. Circadian rhythm connections to oxidative stress: Implications for human health. Antioxid. Redox Signal. 2013, 19, 192–208. [Google Scholar] [CrossRef] [Green Version]
- Colombini, B.; Dinu, M.; Murgo, E.; Lotti, S.; Tarquini, R.; Sofi, F.; Mazzoccoli, G. Ageing and Low-Level Chronic Inflammation: The Role of the Biological Clock. Antioxidants 2022, 11, 2228. [Google Scholar] [CrossRef]
- Chen, Z.; Xiong, Z.F.; Liu, X. Research progress on the interaction between circadian clock and early vascular aging. Exp. Gerontol. 2021, 146, 111241. [Google Scholar] [CrossRef]
- Ribas-Latre, A.; Eckel-Mahan, K. Nutrients and the Circadian Clock: A Partnership Controlling Adipose Tissue Function and Health. Nutrients 2022, 14, 2084. [Google Scholar] [CrossRef]
- Maury, E.; Navez, B.; Brichard, S.M. Circadian clock dysfunction in human omental fat links obesity to metabolic inflammation. Nat. Commun. 2021, 12, 2388. [Google Scholar] [CrossRef]
- Nernpermpisooth, N.; Qiu, S.; Mintz, J.D.; Suvitayavat, W.; Thirawarapan, S.; Rudic, D.R.; Fulton, D.J.; Stepp, D.W. Obesity Alters the Peripheral Circadian Clock in the Aorta and, Microcirculation. Microcirculation 2015, 22, 257–266. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Antioxidants | Sources | Functions | |||
|---|---|---|---|---|---|
| Antioxidants | Anti-Vascular Aging | Anti-Inflammatory | Anti-Obesity | ||
| Quercetin | Onions, grapes, cherries, apples, mangoes, citrus fruits, buckwheat, plums, tomatoes, and tea | ROS scavenger, protecting macromolecules, inhibition of lipid peroxidation, inhibiting ROS production, up-regulating the expression of SOD, CAT, and GSH | Selectively removing the aging endothelial cells, vasodilatory effect due to up-regulation of eNOS, displaying anti-hypertensive and anti-atherosclerosis effects | Inhibiting neutrophil infiltration and NLRP3, NF-κB, ROS/AMPK pathways; reducing TNF-α | Reducing inflammation in BAT, promoting thermogenesis and browning of WAT |
| Vitamins E and C | Vitamin C: fruits and fruiting vegetables; vitamin E: vegetable oils, olives, non-citrus fruits, and nuts | Reducing markers of hypoxia andER stress promoting the secretion of adiponectin, protecting endothelial cells from oxidative stress | Improve endothelial function, protecting endothelial cells from chromatin damages andtelomere shortening | Reducing levels of IL-6 and hs-CRP, leptin, resistin, TNF-α, and PAI-1 | Reducing collagen deposition in visceral adipose tissue |
| Allium sativum | Garlic | Improving serum levels of MDA and SOD | Protecting endothelial cells and the endothelial function due to increasing total antioxidant capacity | Iinhibiting NF-κB activation | Reducing total fat mass |
| Resveratrol | Grapes, apples, blueberries, plums, wine, and peanuts | Reducing H2O2 production, increasing the level of regulatory T cells; reducing the ROS level by inhibiting the MAPK pathways | Increasing NO production, up-regulating the expression of eNOS; suppressing the synthesis of endothelin-1 | Inhibiting the PKA and Akt/PBB pathway | WAT remodelling to BAT, reducing the accumulation of glycerol in adipose tissue; promoting thermogenesis by activation of SIRT1 and suppressing white adipogenesis |
| Carnitine | Animal products such as the richest red meat, poultry, fish, and dairy foods synthesized endogenously from lysine and methionine in kidneys, liver and brain | Inducing the production of SOD, reducing ROS production, control of ketogenesis and glucogenesis, and stabilization of cell membranes; improving fatty acid oxidation processes | Prevents H2O2-induced endothelial senescence; anti-atherosclerotic agent by reducing serum cholesterol levels | Suppressing NF-ĸB signaling pathway, reducing TNF-α, IL-1, and IL-6 in adipose tissue | Increasing mitochondrial β-oxidation of fatty acids, reducing triglyceride synthesis and serum cholesterol, LDL-C, triglycerides, and free fatty acids |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharebiani, H.; Keramat, S.; Chavoshan, A.; Fazeli, B.; Stanek, A. The Influence of Antioxidants on Oxidative Stress-Induced Vascular Aging in Obesity. Antioxidants 2023, 12, 1295. https://doi.org/10.3390/antiox12061295
Sharebiani H, Keramat S, Chavoshan A, Fazeli B, Stanek A. The Influence of Antioxidants on Oxidative Stress-Induced Vascular Aging in Obesity. Antioxidants. 2023; 12(6):1295. https://doi.org/10.3390/antiox12061295
Chicago/Turabian StyleSharebiani, Hiva, Shayan Keramat, Abdolali Chavoshan, Bahar Fazeli, and Agata Stanek. 2023. "The Influence of Antioxidants on Oxidative Stress-Induced Vascular Aging in Obesity" Antioxidants 12, no. 6: 1295. https://doi.org/10.3390/antiox12061295