

Antioxidant Enzyme Activity and Serum HSP70 Concentrations in Relation to Insulin Resistance and Lipid Profile in Lean and Overweight Young Men

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measures
2.2.1. Biochemical Parameters of Venous Blood
2.2.2. Statistical Analysis
3. Results
3.1. Anthropometrical and Biochemical Parameters in the Study Groups
3.2. Erythrocyte Antioxidant Enzyme Activity and Glutathione Concentration in Study Groups
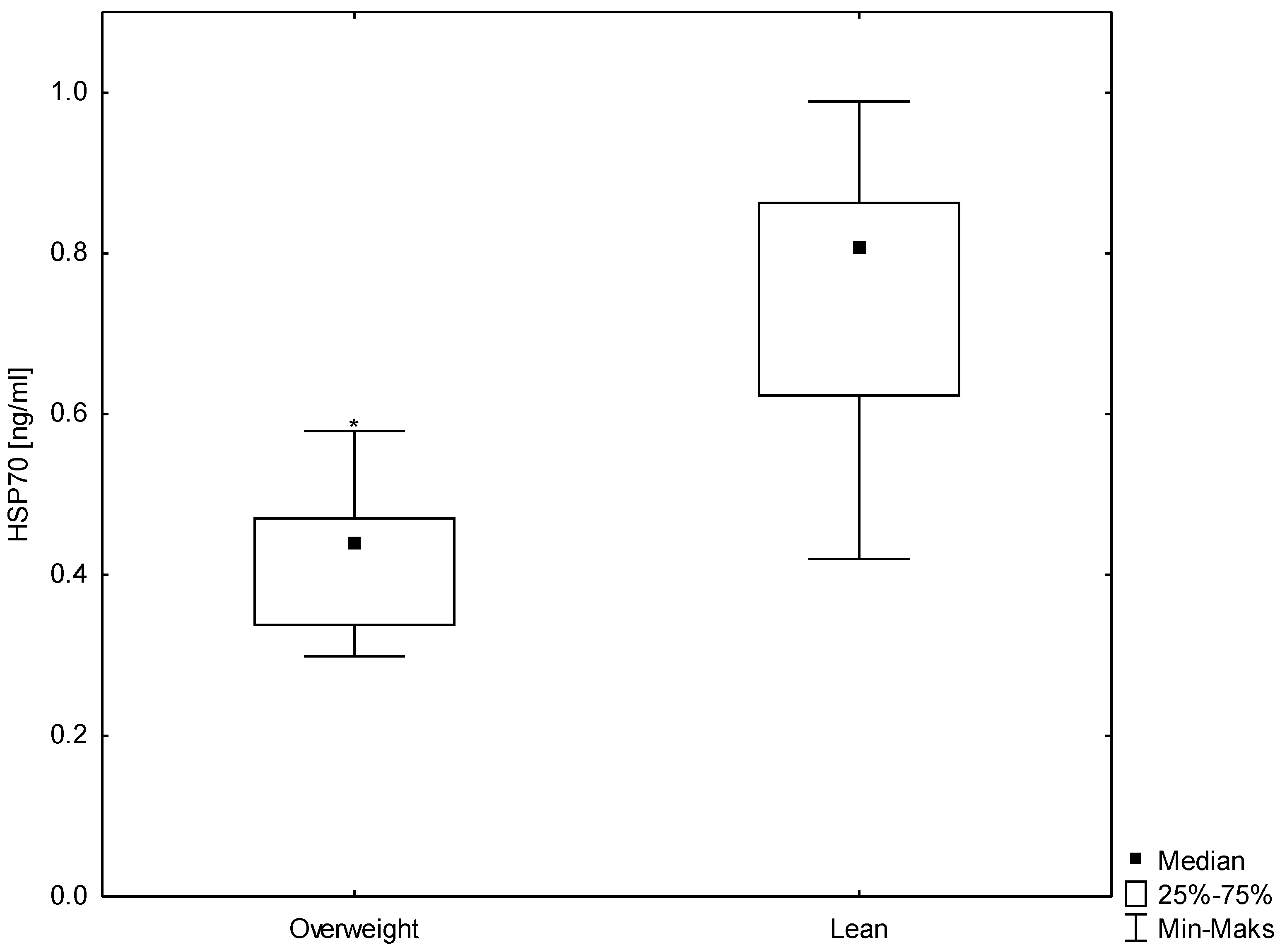
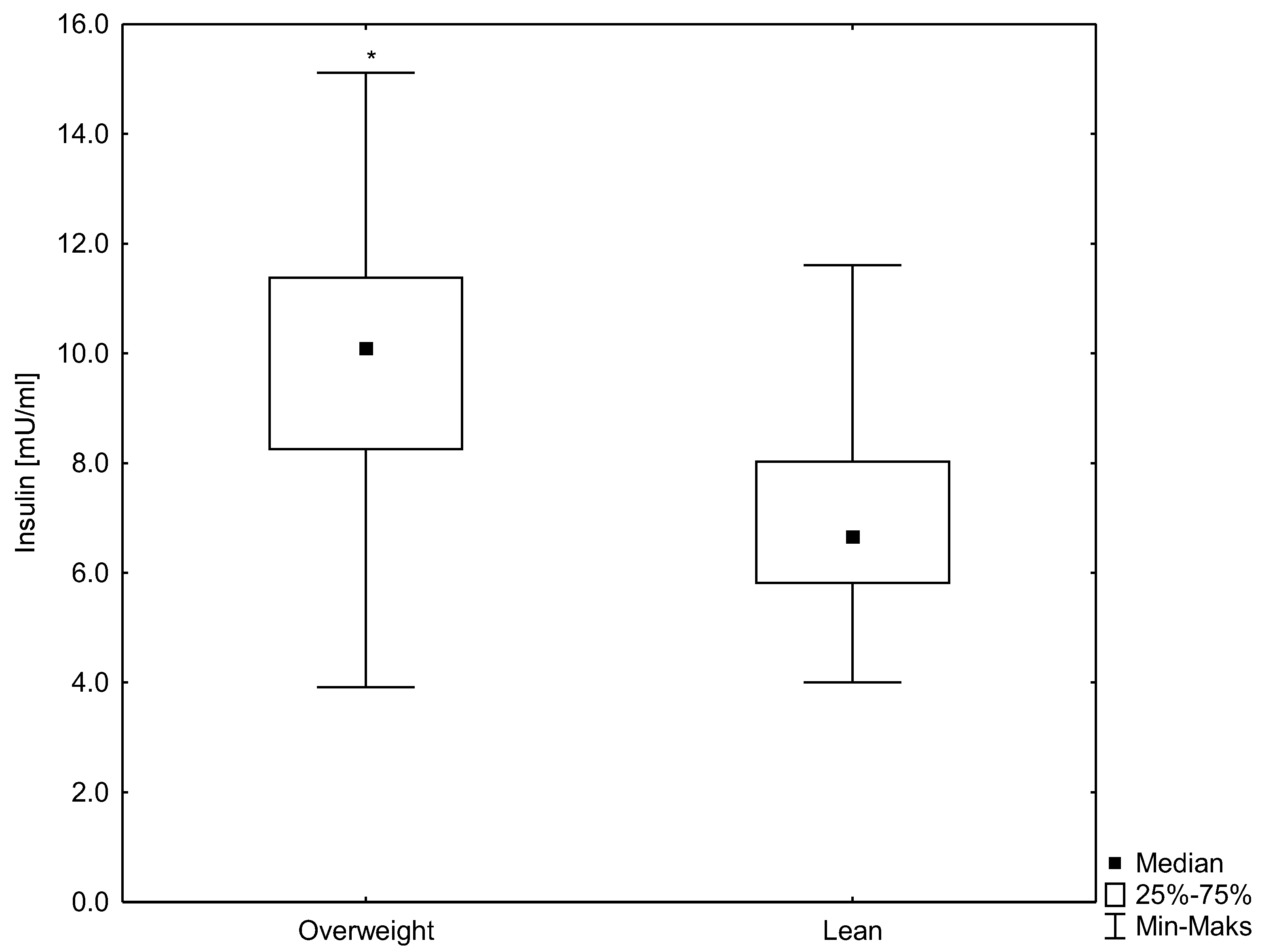
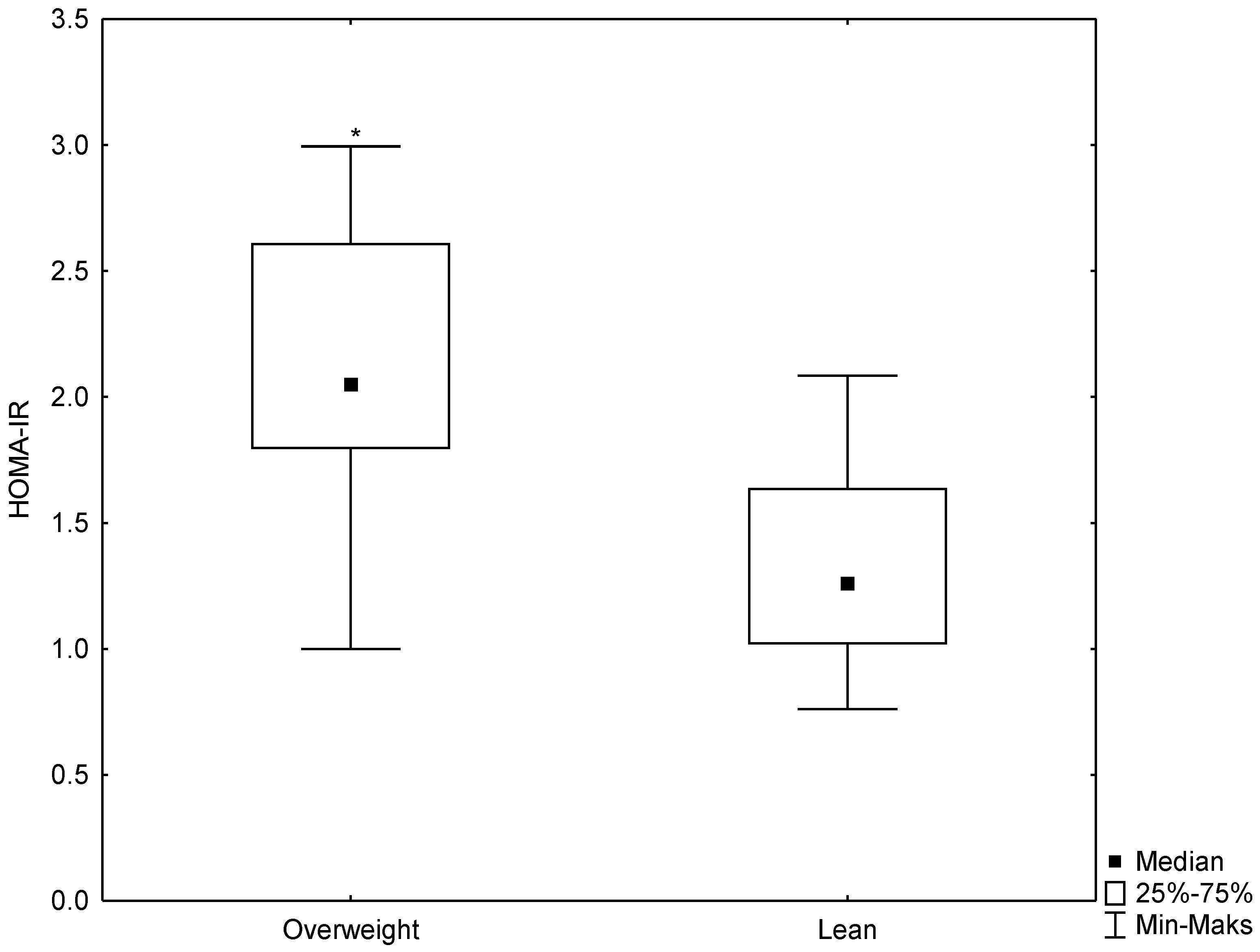
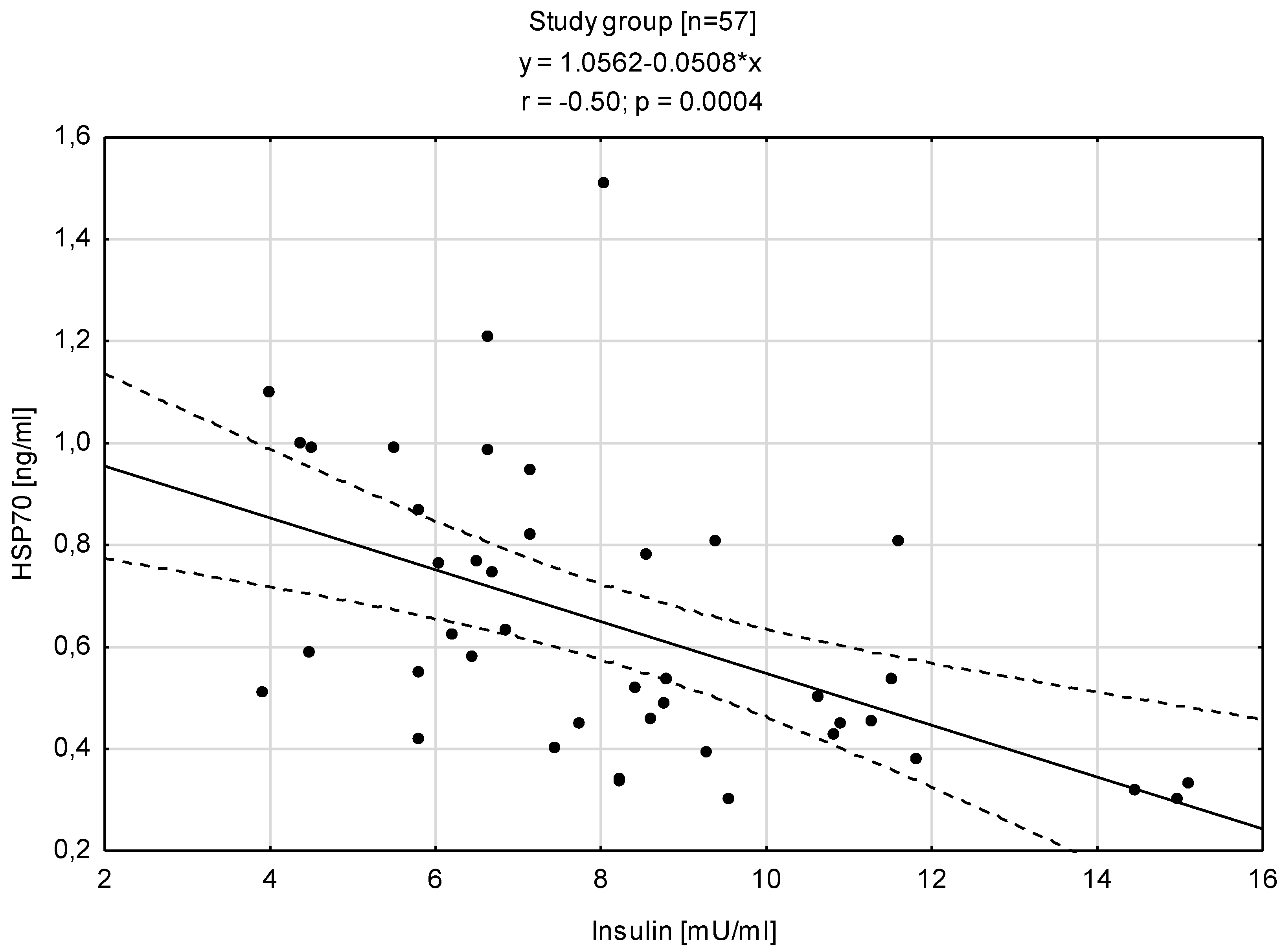
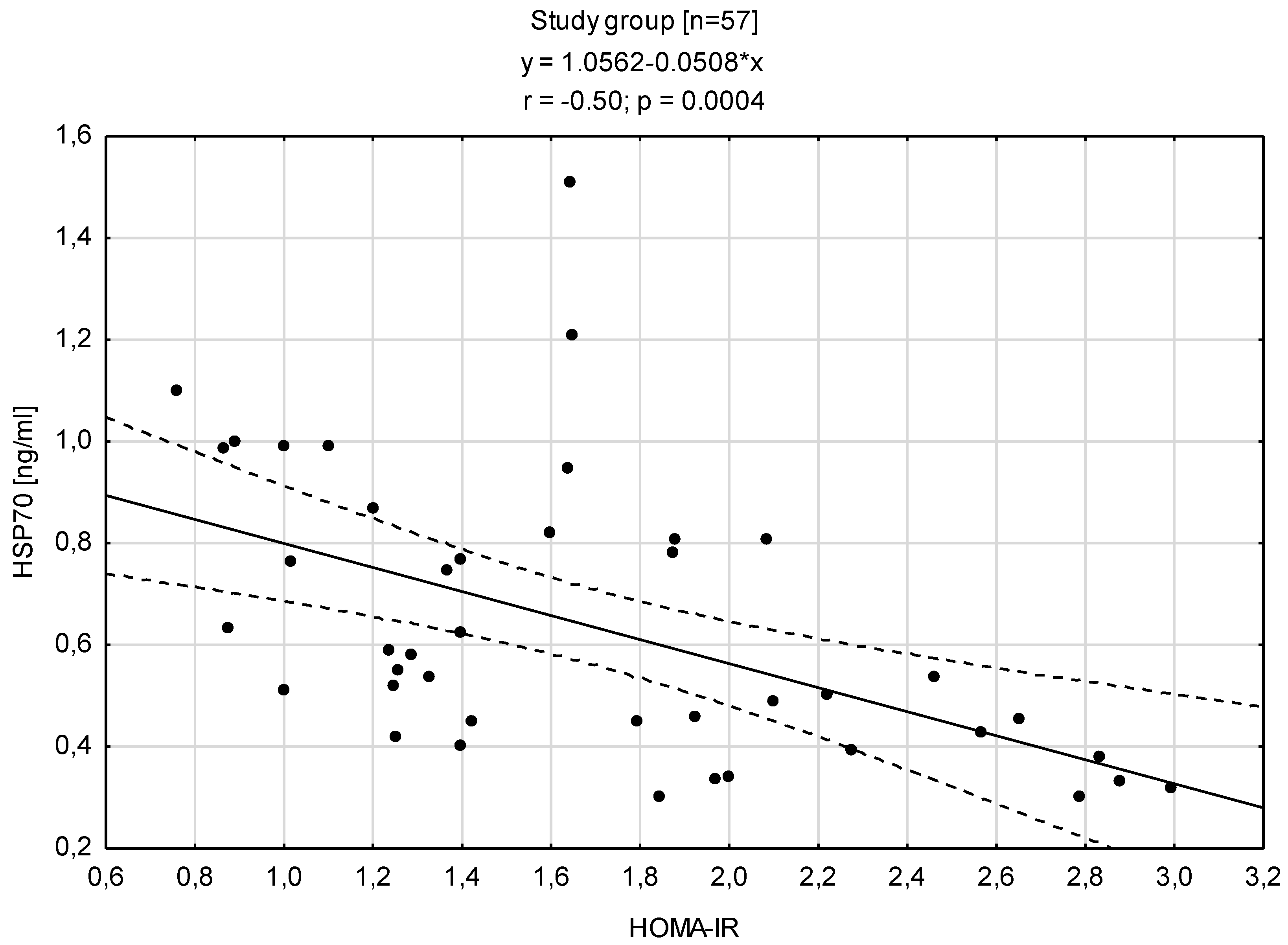
3.3. Relationships between HSP70 Level and Concentration of Insulin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jakubiak, G.K.; Osadnik, K.; Lejawa, M.; Osadnik, T.; Goławski, M.; Lewandowski, P.; Pawlas, N. “Obesity and Insulin Resistance” Is the Component of the Metabolic Syndrome Most Strongly Associated with Oxidative Stress. Antioxidants 2022, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative Stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardaweel, S.K.; Gul, M.; Alzweiri, M.; Ishaqat, A.; ALSalamat, H.A.; Bashatwah, R.M. Reactive Oxygen Species: The Dual Role in Physiological and Pathological Conditions of the Human Body. Eurasian J. Med. 2018, 50, 193–201. [Google Scholar] [CrossRef]
- Schuermann, D.; Mevissen, M. Manmade Electromagnetic Fields and Oxidative Stress—Biological Effects and Consequences for Health. Int. J. Mol. Sci. 2021, 22, 3772. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 Chaperone Network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Liu, Q.; Liang, C.; Zhou, L. Structural and Functional Analysis of the Hsp70/Hsp40 Chaperone System. Protein Sci. 2020, 29, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Lubkowska, A.; Pluta, W.; Strońska, A.; Lalko, A. Role of Heat Shock Proteins (Hsp70 and Hsp90) in Viral Infection. Int. J. Mol. Sci. 2021, 22, 9366. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Copani, A.; Testa, D.; Ravagna, A.; Spadaro, F.; Tendi, E.; Nicoletti, V.G.; Giuffrida Stella, A.M. Nitric Oxide Synthase Induction in Astroglial Cell Cultures: Effect on Heat Shock Protein 70 Synthesis and Oxidant/Antioxidant Balance. J. Neurosci. Res. 2000, 60, 613–622. [Google Scholar] [CrossRef]
- Calabrese, V.; Scapagnini, G.; Catalano, C.; Bates, T.E.; Dinotta, F.; Micali, G.; Giuffrida Stella, A.M. Induction of Heat Shock Protein Synthesis in Human Skin Fibroblasts in Response to Oxidative Stress: Regulation by a Natural Antioxidant from Rosemary Extract. Int. J. Tissue React. 2001, 23, 51–58. [Google Scholar]
- Yan, L.J.; Christians, E.S.; Liu, L.; Xiao, X.Z.; Sohal, R.S.; Benjamin, I.J. Mouse Heat Shock Transcription Factor 1 Deficiency Alters Cardiac Redox Homeostasis and Increases Mitochondrial Oxidative Damage. EMBO J. 2002, 21, 5164–5172. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Maruyama, S.; Sato, W.; Morita, Y.; Sato, F.; Miki, Y.; Kato, S.; Katsuno, M.; Sobue, G.; Yuzawa, Y.; et al. Geranylgeranylacetone Ameliorates Ischemic Acute Renal Failure via Induction of Hsp70. Kidney Int. 2005, 67, 2210–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Wharton, W.; Moseley, P.; Shi, H. Heat Shock Protein 70 Regulates Cellular Redox Status by Modulating Glutathione-Related Enzyme Activities. Cell Stress Chaperones 2007, 12, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M. Effects of Antioxidant Enzymes in the Molecular Control of Reactive Oxygen Species Toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; ISBN 9780198717478. [Google Scholar]
- Vávrová, L.; Kodydková, J.; Zeman, M.; Dušejovská, M.; Macášek, J.; Stanková, B.; Tvrzická, E.; Žák, A. Altered Activities of Antioxidant Enzymes in Patients with Metabolic Syndrome. Obes. Facts 2013, 6, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Karaouzene, N.; Merzouk, H.; Aribi, M.; Merzouk, S.A.; Yahia Berrouiguet, A.; Tessier, C.; Narce, M. Effects of the Association of Aging and Obesity on Lipids, Lipoproteins and Oxidative Stress Biomarkers: A Comparison of Older with Young Men. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 792–799. [Google Scholar] [CrossRef]
- Erdeve, O.; Siklar, Z.; Kocaturk, P.A.; Dallar, Y.; Kavas, G.O. Antioxidant Superoxide Dismutase Activity in Obese Children. Biol. Trace Elem. Res. 2004, 98, 219–228. [Google Scholar] [CrossRef] [PubMed]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of Insulin Resistance and Obesity in Mice Overexpressing Cellular Glutathione Peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar] [CrossRef] [Green Version]
- Rupérez, A.I.; Olza, J.; Gil-Campos, M.; Leis, R.; Mesa, M.D.; Tojo, R.; Cañete, R.; Gil, Á.; Aguilera, C.M. Association of Genetic Polymorphisms for Glutathione Peroxidase Genes with Obesity in Spanish Children. J. Nutr. Nutr. 2014, 7, 130–142. [Google Scholar] [CrossRef]
- Chen, X.; Scholl, T.O.; Leskiw, M.J.; Donaldson, M.R.; Stein, T.P. Association of Glutathione Peroxidase Activity with Insulin Resistance and Dietary Fat Intake during Normal Pregnancy. J. Clin. Endocrinol. Metab. 2003, 88, 5963–5968. [Google Scholar] [CrossRef] [Green Version]
- Ferro, F.E.D.; De Sousa Lima, V.B.; Soares, N.R.M.; De Sousa Almondes, K.G.; Pires, L.V.; Cozzolino, S.M.F.; Do Nascimento Marreiro, D. Parameters of Metabolic Syndrome and Its Relationship with Zincemia and Activities of Superoxide Dismutase and Glutathione Peroxidase in Obese Women. Biol. Trace Elem. Res. 2011, 143, 787–793. [Google Scholar] [CrossRef]
- Gunawardena, H.P.; Silva, K.D.R.R.; Sivakanesan, R.; Katulanda, P. Increased Lipid Peroxidation and Erythrocyte Glutathione Peroxidase Activity of Patients with Type 2 Diabetes Mellitus: Implications for Obesity and Central Obesity. Obes. Med. 2019, 15, 100118. [Google Scholar] [CrossRef]
- Baez-Duarte, B.G.; Zamora-Ginez, I.; Mendoza-Carrera, F.; Ruiz-Vivanco, G.; Torres-Rasgado, E.; Gonzalez-Mejia, M.E.; Garcia-Zapien, A.; Flores-Martinez, S.E.; Perez-Fuentes, R. Serum Levels of Glutathione Peroxidase 3 in Overweight and Obese Subjects from Central Mexico. Arch. Med. Res. 2012, 43, 541–547. [Google Scholar] [CrossRef]
- Nakao, C.; Ookawara, T.; Sato, Y.; Kizaki, T.; Imazeki, N.; Matsubara, O.; Haga, S.; Suzuki, K.; Taniguchi, N.; Ohno, H. Extracellular Superoxide Dismutase in Tissues from Obese (Ob/Ob) Mice. Free Radic. Res. 2000, 33, 229–241. [Google Scholar] [CrossRef]
- Asea, A.; Rehli, M.; Kabingu, E.; Boch, J.A.; Baré, O.; Auron, P.E.; Stevenson, M.A.; Calderwood, S.K. Novel Signal Transduction Pathway Utilized by Extracellular HSP70. J. Biol. Chem. 2002, 277, 15028–15034. [Google Scholar] [CrossRef] [Green Version]
- Lehner, T.; Wang, Y.; Whittall, T.; McGowan, E.; Kelly, C.G.; Singh, M. Functional Domains of HSP70 Stimulate Generation of Cytokines and Chemokines, Maturation of Dendritic Cells and Adjuvanticity. Biochem. Soc. Trans. 2004, 32, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Van Eden, W.; Van der Zee, R.; Prakken, B. Heat-Shock Proteins Induce T-Cell Regulation of Chronic Inflammation. Nat. Rev. Immunol. 2005, 5, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Toda, I.M.; De la Fuente, M. The Role of Hsp70 in Oxi-Inflamm-Aging and Its Use as a Potential Biomarker of Lifespan. Biogerontology 2015, 16, 709–721. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Mambula, S.S.; Gray, P.J.; Theriault, J.R. Extracellular Heat Shock Proteins in Cell Signaling. FEBS Lett. 2007, 581, 3689–3694. [Google Scholar] [CrossRef] [Green Version]
- Ortega, E.; Bote, M.E.; Besedovsky, H.O.; del Rey, A. Hsp72, Inflammation, and Aging: Causes, Consequences, and Perspectives. Ann. N. Y. Acad. Sci. 2012, 1261, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Fuente, M.; Miquel, J. An Update of the Oxidation-Inflammation Theory of Aging: The Involvement of the Immune System in Oxi-Inflamm-Aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef]
- Rea, S.L.; Wu, D.; Cypser, J.R.; Vaupel, J.W.; Johnson, T.E. A Stress-Sensitive Reporter Predicts Longevity in Isogenic Populations of Caenorhabditis Elegans. Nat. Genet. 2005, 37, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Senf, S.M. Skeletal Muscle Heat Shock Protein 70: Diverse Functions and Therapeutic Potential for Wasting Disorders. Front. Physiol. 2013, 4, 330. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.; Hait, S.; Andrews-Shigaki, B.; Carus, S.; Deuster, P. Plasma HSP70 Levels Correlate with Health Risk Factors and Insulin Resistance in African American Subjects. Exp. Clin. Endocrinol. Diabetes 2014, 122, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Cangeri Di Naso, F.; Rosa Porto, R.; Sarubbi Fillmann, H.; Maggioni, L.; Vontobel Padoin, A.; Jacques Ramos, R.; Corá Mottin, C.; Bittencourt, A.; Anair Possa Marroni, N.; Ivo Homem de Bittencourt, P. Obesity Depresses the Anti-Inflammatory HSP70 Pathway, Contributing to NAFLD Progression. Obesity 2015, 23, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Mambula, S.S.; Stevenson, M.A.; Ogawa, K.; Calderwood, S.K. Mechanisms for Hsp70 Secretion: Crossing Membranes without a Leader. Methods 2007, 43, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Hunter-Lavin, C.; Davies, E.L.; Bacelar, M.M.F.V.G.; Marshall, M.J.; Andrew, S.M.; Williams, J.H.H. Hsp70 Release from Peripheral Blood Mononuclear Cells. Biochem. Biophys. Res. Commun. 2004, 324, 511–517. [Google Scholar] [CrossRef]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory Mechanisms in Obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [Green Version]
- Kurucz, I.; Morva, A.; Vaag, A.; Eriksson, K.-F.; Huang, X.; Groop, L.; Koranyi, L. Decreased Expression of Heat Shock Protein 72 in Skeletal Muscle of Patients with Type 2 Diabetes Correlates with Insulin Resistance. Diabetes 2002, 51, 1102–1109. [Google Scholar] [CrossRef] [Green Version]
- Bruce, C.R.; Carey, A.L.; Hawley, J.A.; Febbraio, M.A. Intramuscular Heat Shock Protein 72 and Heme Oxygenase-1 MRNA Are Reduced in Patients with Type 2 Diabetes. Diabetes 2003, 52, 2338–2345. [Google Scholar] [CrossRef] [Green Version]
- Najemnikova, E.; Rodgers, C.D.; Locke, M. Altered Heat Stress Response Following Streptozotocin-Induced Diabetes. Cell Stress Chaperones 2007, 12, 342. [Google Scholar] [CrossRef]
- Chung, J.; Nguyen, A.-K.; Henstridge, D.C.; Holmes, A.G.; Chan, M.H.S.; Mesa, J.L.; Lancaster, G.I.; Southgate, R.J.; Bruce, C.R.; Duffy, S.J.; et al. HSP72 Protects against Obesity-Induced Insulin Resistance. Proc. Natl. Acad. Sci. USA 2008, 105, 1739–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sárközy, M.; Zvara, A.; Gyémánt, N.; Fekete, V.; Kocsis, G.F.; Pipis, J.; Szűcs, G.; Csonka, C.; Puskás, L.G.; Ferdinandy, P.; et al. Metabolic Syndrome Influences Cardiac Gene Expression Pattern at the Transcript Level in Male ZDF Rats. Cardiovasc. Diabetol. 2013, 12, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiss, A.; Khadir, A.; Abubaker, J.; Abu-Farha, M.; Al-Khairi, I.; Cherian, P.; John, J.; Kavalakatt, S.; Warsame, S.; Al-Ghimlas, F.; et al. Immunohistochemical Profiling of the Heat Shock Response in Obese Non-Diabetic Subjects Revealed Impaired Expression of Heat Shock Proteins in the Adipose Tissue. Lipids Health Dis. 2014, 13, 106. [Google Scholar] [CrossRef] [Green Version]
- Tarantino, G.; Finelli, C.; Colao, A.; Capone, D.; Tarantino, M.; Grimaldi, E.; Chianese, D.; Gioia, S.; Pasanisi, F.; Contaldo, F.; et al. Are Hepatic Steatosis and Carotid Intima Media Thickness Associated in Obese Patients with Normal or Slightly Elevated Gamma-Glutamyl-Transferase? J. Transl. Med. 2012, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Nakhjavani, M.; Morteza, A.; Meysamie, A.; Esteghamati, A.; Khalilzadeh, O.; Esfahanian, F.; Khajeali, L.; Feiz, F. Serum Heat Shock Protein 70 and Oxidized LDL in Patients with Type 2 Diabetes: Does Sex Matter? Cell Stress Chaperones 2011, 16, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oglesbee, M.J.; Herdman, A.V.; Passmore, G.G.; Hoffman, W.H. Diabetic Ketoacidosis Increases Extracellular Levels of the Major Inducible 70-KDa Heat Shock Protein. Clin. Biochem. 2005, 38, 900–904. [Google Scholar] [CrossRef]
- Rodrigues-Krause, J.; Krause, M.; O’Hagan, C.; De Vito, G.; Boreham, C.; Murphy, C.; Newsholme, P.; Colleran, G. Divergence of Intracellular and Extracellular HSP72 in Type 2 Diabetes: Does Fat Matter? Cell Stress Chaperones 2012, 17, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 Chaperones: Cellular Functions and Molecular Mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Njemini, R.; Demanet, C.; Mets, T. Inflammatory Status as an Important Determinant of Heat Shock Protein 70 Serum Concentrations during Aging. Biogerontology 2004, 5, 31–38. [Google Scholar] [CrossRef]
- Gruden, G.; Bruno, G.; Chaturvedi, N.; Burt, D.; Pinach, S.; Schalkwijk, C.; Stehouwer, C.D.; Witte, D.R.; Fuller, J.H.; Cavallo-Perin, P. ANTI-HSP60 and ANTI-HSP70 Antibody Levels and Micro/ Macrovascular Complications in Type 1 Diabetes: The EURODIAB Study. J. Intern. Med. 2009, 266, 527–536. [Google Scholar] [CrossRef]
- Hooper, P.L. Insulin Signaling, GSK-3, Heat Shock Proteins and the Natural History of Type 2 Diabetes Mellitus: A Hypothesis. Metab. Syndr. Relat. Disord. 2007, 5, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Hooper, P.L.; Hooper, P.L. Inflammation, Heat Shock Proteins, and Type 2 Diabetes. Cell Stress Chaperones 2009, 14, 113–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henstridge, D.C.; Forbes, J.M.; Penfold, S.A.; Formosa, M.F.; Dougherty, S.; Gasser, A.; de Courten, M.P.; Cooper, M.E.; Kingwell, B.A.; de Courten, B. The Relationship between Heat Shock Protein 72 Expression in Skeletal Muscle and Insulin Sensitivity Is Dependent on Adiposity. Metabolism 2010, 59, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Gupte, A.A.; Bomhoff, G.L.; Swerdlow, R.H.; Geiger, P.C. Heat Treatment Improves Glucose Tolerance and Prevents Skeletal Muscle Insulin Resistance in Rats Fed a High-Fat Diet. Diabetes 2009, 58, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simar, D.; Jacques, A.; Caillaud, C. Heat Shock Proteins Induction Reduces Stress Kinases Activation, Potentially Improving Insulin Signalling in Monocytes from Obese Subjects. Cell Stress Chaperones 2012, 17, 615–621. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lean n = 30 Median (Q25–Q75) | Overweight n = 27 Median (Q25–Q75) | p< | |
|---|---|---|---|
| Age (years) | 23.0 (23–23) | 23.5 (23–25) | NS |
| BMI (kg/m2) | 23.6 (22.83–24.25) | 26.9 (26.30–28.40) | 0.007 |
| Extracellular Hb (g/dL) | 30.0 (26.53–37.07) | 31.2 (28.48–39.99) | NS |
| Haptoglobin (g/L) | 0.3 (0.10–0.37) | 0.2 (0.14–0.32) | NS |
| Total bilirubin (mg/dL) | 1.5 (1.15–1.69) | 1.4 (1.27–1.44) | NS |
| Erythropoietin (ng/mL) | 263 (210–320) | 246 (200–278) | NS |
| Il-3 (pg/mL) | 18.1 (10.14–28.17) | 18.9 (13.78–35.29) | NS |
| Total protein (g/dL) | 7.1 (6.66–7.54) | 7.4 (6.87–7.43) | NS |
| Albumin (g/dL) | 51.2 (47.53–54.38) | 48.5 (42.90–50.86) | NS |
| Uric acid (mg/dL) | 5.0 (4.43–6.13) | 5.8 (4.71–7.01) | NS |
| Glucose (mM) | 4.8 (4.30–5.15) | 5.3 (4.33–5.52) | NS |
| Total cholesterol (mg/dL) | 196 (154–216) | 188 (163–211) | NS |
| HDL cholesterol (mg/dL) | 25.3 (24.25–28.14) | 23.9 (22.44–27.72) | NS |
| LDL cholesterol (mg/dL) | 142 (103–158) | 135 (112–162) | NS |
| Triglycerides (mg/dL) | 113 (82.60–149) | 117 (90.64–139) | NS |
| TG:TCh | 0.6 (0.46–0.80) | 0.6 (0-50–0.80) | NS |
| TCh:HDL | 7.0 (6.07–8.73) | 7.5 (6.78–8.42) | NS |
| LDL:HDL | 5.1 (4.05–6.55) | 5.4 (4.90–6.42) | NS |
| ApoA (ng/mL) | 2.5 (2.46–2.55) | 2.5 (2.49–2.51) | NS |
| ApoB (ng/mL) | 1.0 (0.98–1.04) | 1.0 (0.96–1.04) | NS |
| Lean n = 30 Median (Q25–Q75) | Overweight n = 27 Median (Q25–Q75) | p< | |
|---|---|---|---|
| SOD (U/gHb) | 762 (512–865) | 872 (862–871) | 0.02 |
| CAT (U/gHb) | 46 (29.66–57.80) | 54 (49.11–59.61) | NS |
| GPX (U/gHb) | 6.7 (5.67–7.68) | 8.3 (5.85–9.89) | 0.05 |
| GST (U/gHb) | 0.2 (0.10–0.25) | 0.2 (0.10–0.21) | NS |
| GSSG-R (U/gHb) | 0.1 (0.06–0.15) | 0.1 (0.06–0.11) | NS |
| GSHreduced (mM) | 722 (667–749) | 770 (716–787) | NS |
| GSSG (mM) | 40 (27.45–47.81) | 41 (36.40–47.65) | NS |
| GSH total (mM) | 762 (690–796) | 811 (757–845) | NS |
| GSH:GSSG | 17 (14.16–23.49) | 19 (14.68–20.88) | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubkowska, A.; Dudzińska, W.; Pluta, W. Antioxidant Enzyme Activity and Serum HSP70 Concentrations in Relation to Insulin Resistance and Lipid Profile in Lean and Overweight Young Men. Antioxidants 2023, 12, 655. https://doi.org/10.3390/antiox12030655
Lubkowska A, Dudzińska W, Pluta W. Antioxidant Enzyme Activity and Serum HSP70 Concentrations in Relation to Insulin Resistance and Lipid Profile in Lean and Overweight Young Men. Antioxidants. 2023; 12(3):655. https://doi.org/10.3390/antiox12030655
Chicago/Turabian StyleLubkowska, Anna, Wioleta Dudzińska, and Waldemar Pluta. 2023. "Antioxidant Enzyme Activity and Serum HSP70 Concentrations in Relation to Insulin Resistance and Lipid Profile in Lean and Overweight Young Men" Antioxidants 12, no. 3: 655. https://doi.org/10.3390/antiox12030655