
Effect of Exercise Repetitions on Arylesterase Activity of PON1 in Plasma of Average-Trained Men—The Dissociation between Activity and Concentration
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
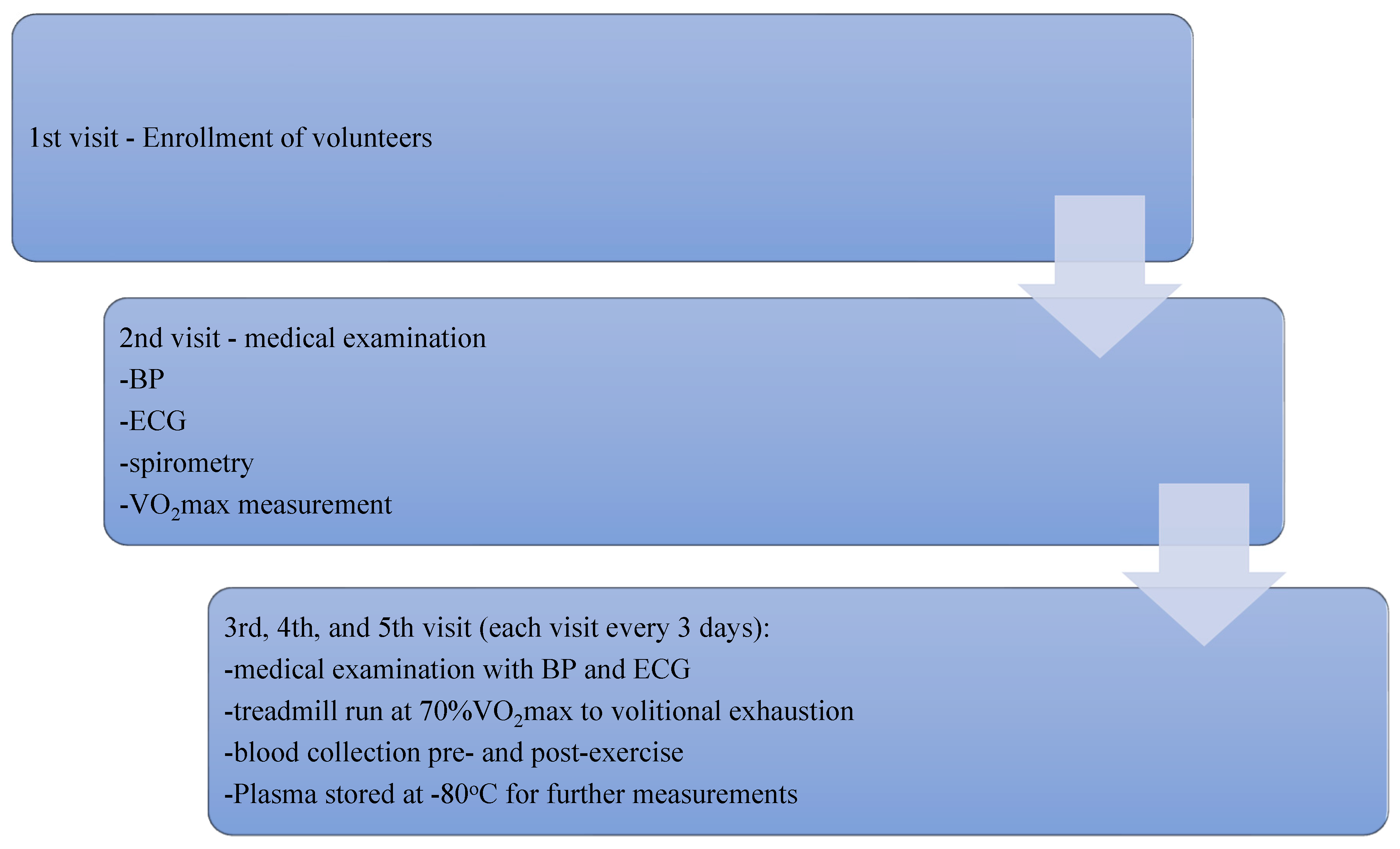
2.2. Study Protocol
2.3. Determinations in Plasma
2.4. Genotyping of the PON1 Gene
2.5. Chemicals
2.6. Statistical Analysis
3. Results
3.1. The Effect of Three Repeated Bouts of Exercise on ARE Activity and Other Biochemical Variables in Plasma
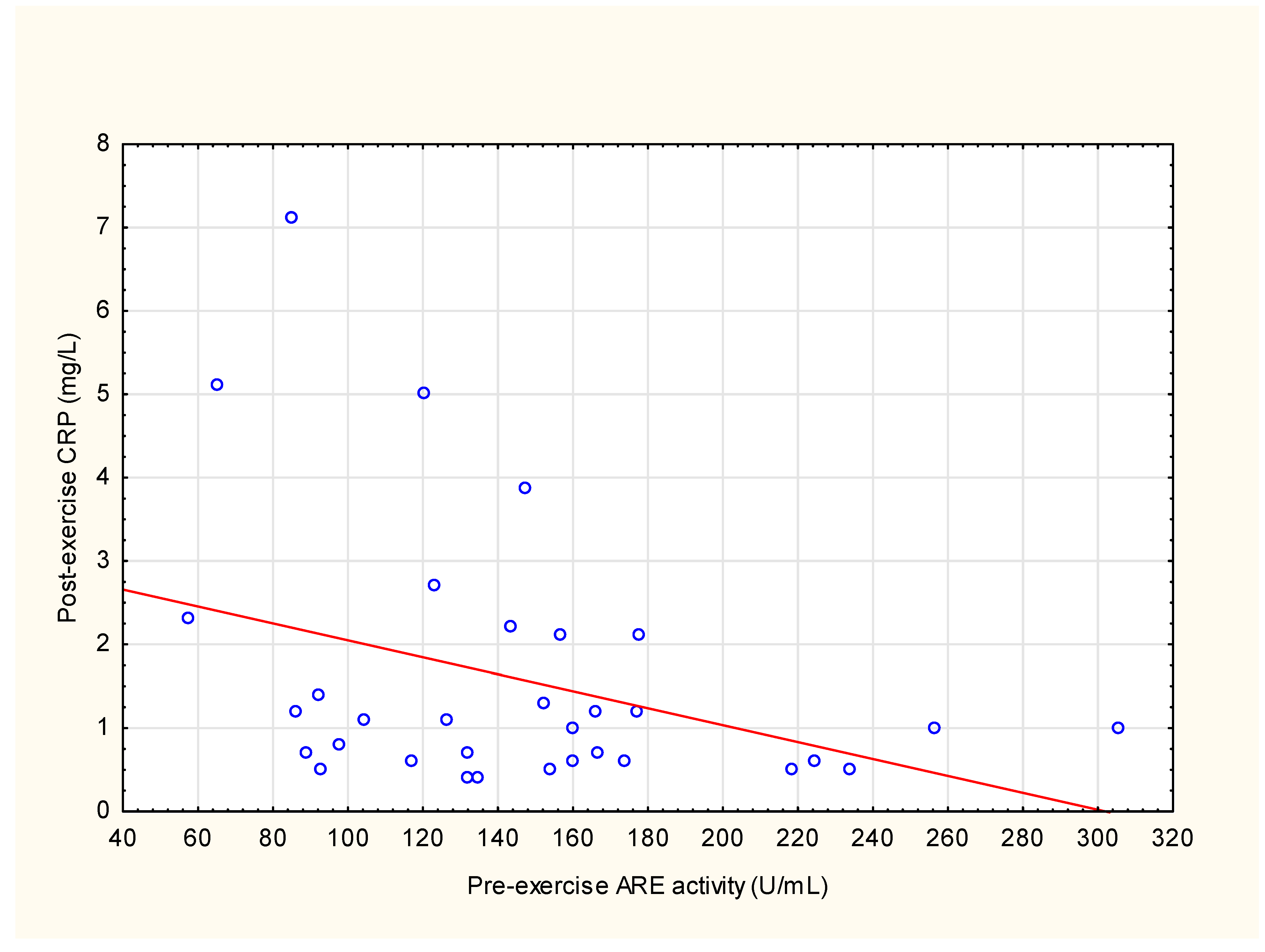
3.2. Correlations between ARE Activity and Other Parameters
4. Discussion
4.1. The Effect of Three Repeated Bouts of Exercise on ARE Activity and Other Biochemical Variables in the Plasma
4.2. Correlations between ARE Activity and Other Parameters
4.3. The Study Limitations
5. Conclusions
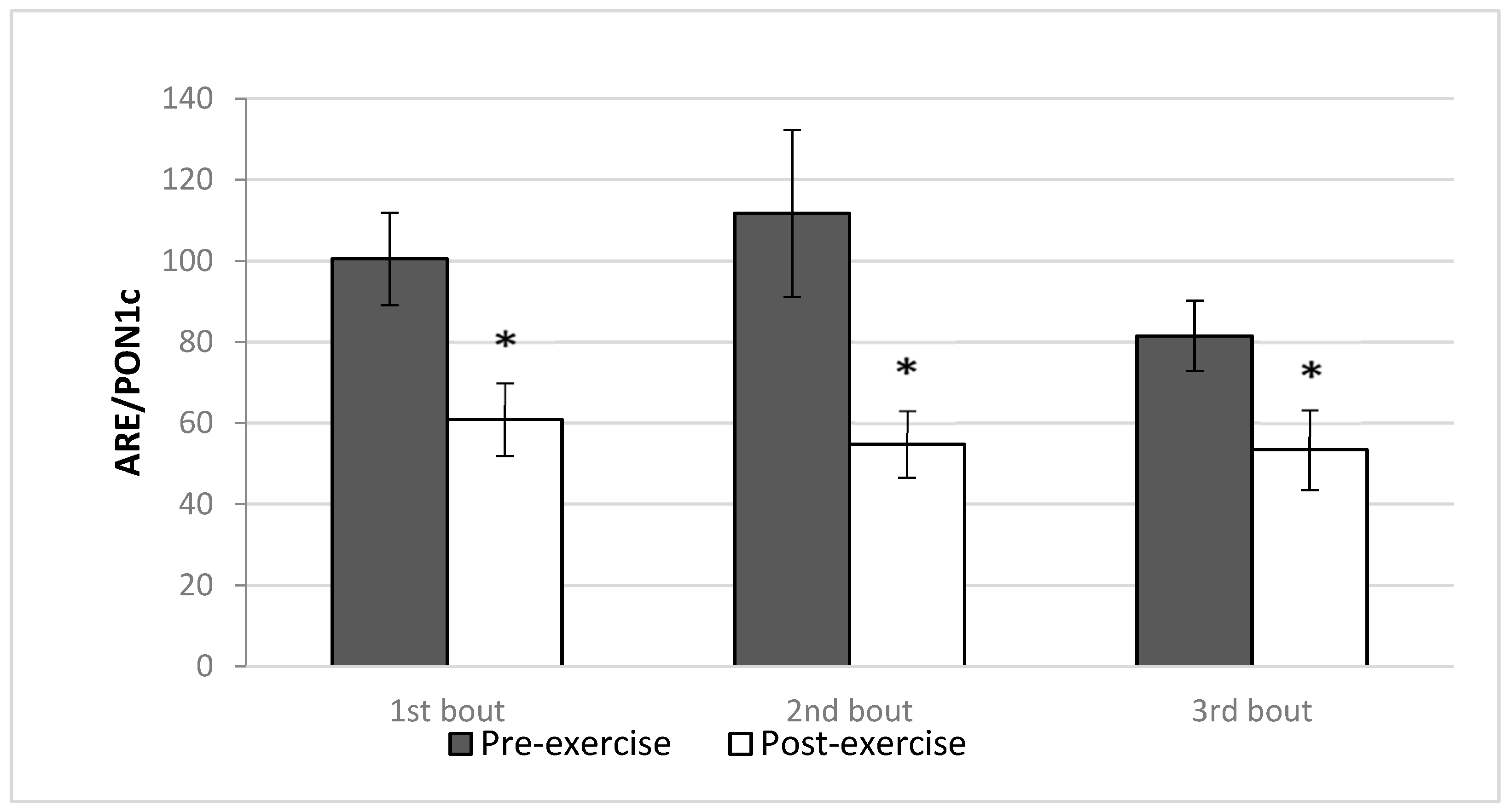
- Dissociation is present between PON1 concentration and ARE activity after strenuous exercise.
- ARE activity may be depleted and consumed under conditions of oxidative stress related to strenuous exercise.
- ARE might influence the intensity of the inflammatory response and muscle damage in response to strenuous exercise.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Romani, R.; De Medio, G.E.; Di Tullio, S.; Lapalombella, R.; Pirisinu, I.; Margonato, V.; Veicsteinas, A.; Marini, M.; Rosi, G. Modulation of Paraoxonase 1 and 3 Expression after Moderate Exercise Training in the Rat. J. Lipid Res. 2009, 50, 2036–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertucci, R.H.; Levada-Pires, A.C.; Rossoni, L.V.; Curi, R.; Pithon-Curi, T.C. Effects of Aerobic Exercise Training on Antioxidant Enzyme Activities and MRNA Levels in Soleus Muscle from Young and Aged Rats. Mech. Ageing Dev. 2007, 128, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Simioni, C.; Zauli, G.; Martelli, A.M.; Vitale, M.; Sacchetti, G.; Gonelli, A.; Neri, L.M. Oxidative Stress: Role of Physical Exercise and Antioxidant Nutraceuticals in Adulthood and Aging. Oncotarget 2018, 9, 17181–17198. [Google Scholar] [CrossRef] [Green Version]
- Radak, Z.; Sasvari, M.; Nyakas, C.; Kaneko, T.; Tahara, S.; Ohno, H.; Goto, S. Single Bout of Exercise Eliminates the Immobilization-Induced Oxidative Stress in Rat Brain. Neurochem. Int. 2001, 39, 33–38. [Google Scholar] [CrossRef]
- Li, L.I.; Gomez-Cabrera, M.-C.; Vina, J. Exercise and Hormesis Activation of Cellular Antioxidant Signaling Pathway. Underst. Modul. Aging 2006, 1067, 425–435. [Google Scholar] [CrossRef]
- Gomez-Cabrera, M.C.; Domenech, E.; Viña, J. Moderate Exercise Is an Antioxidant: Upregulation of Antioxidant Genes by Training. Free Radic. Biol. Med. 2008, 44, 126–131. [Google Scholar] [CrossRef]
- Otocka-Kmiecik, A.; Orłowska-Majdak, M. The Role of Genetic (PON1 Polymorphism) and Environmental Factors, Especially Physical Activity, in Antioxidant Function of Paraoxonase. Postpy Hig. Med. Dosw. 2009, 63, 668–677. [Google Scholar]
- Brites, F.; Martin, M.; Guillas, I.; Kontush, A. Antioxidative Activity of High-Density Lipoprotein (HDL): Mechanistic Insights into Potential Clinical Benefit. BBA Clin. 2017, 8, 66–77. [Google Scholar] [CrossRef]
- Perla-Kaján, J.; Jakubowski, H. Paraoxonase 1 Protects against Protein N-homocysteinylation in Humans. FASEB J. 2010, 24, 931–936. [Google Scholar] [CrossRef]
- Mackness, B.; Durrington, P.; McElduff, P.; Yarnell, J.; Azam, N.; Watt, M.; Mackness, M. Low Paraoxonase Activity Predicts Coronary Events in the Caerphilly Prospective Study. Circulation 2003, 107, 2775–2779. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, T.; Nicholls, S.J.; Topol, E.J.; Zhang, R.; Yang, X.; Schmitt, D.; Fu, X.; Shao, M.; Brennan, D.M.; Ellis, S.G.; et al. Relationship of Paraoxonase 1 (PON1) Gene Polymorphisms and Functional Activity with Systemic Oxidative Stress and Cardiovascular Risk. JAMA J. Am. Med. Assoc. 2008, 299, 1265–1276. [Google Scholar] [CrossRef] [Green Version]
- Maturu, V.N.; Gupta, N.; Singh, G.; Gill, K.; Sharma, Y.P.; Singh, S. Serum Paraoxonase (PON1) Activity in North-West Indian Punjabi’s with Acute Myocardial Infarction. Indian J. Clin. Biochem. 2013, 28, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Mackness, B.; Mackness, M.I.; Arrol, S.; Turkie, W.; Durrington, P.N. Effect of the Human Serum Paraoxonase 55 and 192 Genetic Polymorphisms on the Protection by High Density Lipoprotein against Low Density Lipoprotein Oxidative Modification. FEBS Lett. 1998, 423, 57–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.H.W.; Hartiala, J.; Fan, Y.; Wu, Y.; Stewart, A.F.R.; Erdmann, J.; Kathiresan, S.; Roberts, R.; McPherson, R.; Allayee, H.; et al. Clinical and Genetic Association of Serum Paraoxonase and Arylesterase Activities with Cardiovascular Risk. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Humbert, R.; Adler, D.A.; Disteche, C.M.; Hassett, C.; Omiecinski, C.J.; Furlong, C.E. The Molecular Basis of the Human Serum Paraoxonase Activity Polymorphism. Nat. Genet. 1993, 3, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Adkins, S.; Gan, K.N.; Mody, M.; La Du, B.N. Molecular Basis for the Polymorphic Forms of Human Serum Paraoxonase/Arylesterase: Glutamine or Arginine at Position 191, for the Respective A or B Allozymes. Am. J. Hum. Genet. 1993, 52, 598–608. [Google Scholar]
- Camps, J.; Marsillach, J.; Joven, J. Pharmacological and Lifestyle Factors Modulating Serum Paraoxonase-1 Activity. Mini Rev. Med. Chem. 2009, 9, 911–920. [Google Scholar] [CrossRef]
- Nalcakan, G.R.; Varol, S.R.; Turgay, F.; Nalcakan, M.; Ozkol, M.Z.; Karamizrak, S.O. Effects of Aerobic Training on Serum Paraoxonase Activity and Its Relationship with PON1-192 Phenotypes in Women. J. Sport Health Sci. 2016, 5, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Otocka-Kmiecik, A.; Bortnik, K.; Szkudlarek, U.; Nowak, D.; Orłowska-Majdak, M. Effect of Exercise on Plasma Paraoxonase1 Activity in Rugby Players: Dependance on Training Experience. Redox Rep. 2013, 18, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckerson, H.W.; Wyte, C.M.; La Du, B.N. The Human Serum Paraoxonase/Arylesterase Polymorphism. Am. J. Hum. Genet. 1983, 35, 1126–1138. [Google Scholar] [PubMed]
- Otocka-Kmiecik, A.; Lewandowski, M.; Stolarek, R.; Szkudlarek, U.; Nowak, D.; Orlowska-Majdak, M. Effect of Single Bout of Maximal Excercise on Plasma Antioxidant Status and Paraoxonase Activity in Young Sportsmen. Redox Rep. 2010, 15, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Otocka-Kmiecik, A.; Lewandowski, M.; Szkudlarek, U.; Nowak, D.; Orlowska-Majdak, M. Aerobic Training Modulates the Effects of Exercise-Induced Oxidative Stress on PON1 Activity: A Preliminary Study. Sci. World J. 2014, 2014, 230271. [Google Scholar] [CrossRef] [Green Version]
- Otocka-Kmiecik, A.; Orłowska-Majdak, M.; Stawski, R.; Szkudlarek, U.; Kosielski, P.; Padula, G.; Gałczyński, S.; Nowak, D. Repetitions of Strenuous Exercise Consistently Increase Paraoxonase 1 Concentration and Activity in Plasma of Average-Trained Men. Oxid. Med. Cell. Longev. 2021, 2021, 2775025. [Google Scholar] [CrossRef] [PubMed]
- Tomás, M.; Elosua, R.; Sentí, M.; Molina, L.; Vila, J.; Anglada, R.; Fitó, M.; Covas, M.I.; Marrugat, J. Paraoxonase1-192 Polymorphism Modulates the Effects of Regular and Acute Exercise on Paraoxonasel Activity. J. Lipid Res. 2002, 43, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.K.; Plaisance, E.P.; Jack Mahurin, A.; Mestek, M.L.; Moncada-Jimenez, J.; Grandjean, P.W. Paraoxonase Responses to Exercise and Niacin Therapy in Men with Metabolic Syndrome. Redox Rep. 2015, 20, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Gharakhanlou, R.; Esmaeil Afzalpour, M.; Gaeini, A.A.; Rahnama, N. Effects of Aerobic Exercises on the Serum Paraoxonase 1/Arylesterase Activity and Lipid Profile in Non-Active Healthy Men. Int. J. Sport. Sci. Eng. 2007, 1, 105–112. [Google Scholar]
- Tsakiris, S.; Karikas, G.A.; Parthimos, T.; Tsakiris, T.; Bakogiannis, C.; Schulpis, K.H. Alpha-Tocopherol Supplementation Prevents the Exercise-Induced Reduction of Serum Paraoxonase 1/Arylesterase Activities in Healthy Individuals. Eur. J. Clin. Nutr. 2009, 63, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Harel, M.; Aharoni, A.; Gaidukov, L.; Brumshtein, B.; Khersonsky, O.; Meged, R.; Dvir, H.; Ravelli, R.B.G.; McCarthy, A.; Toker, L.; et al. Structure and Evolution of the Serum Paraoxonase Family of Detoxifying and Anti-Atherosclerotic Enzymes. Nat. Struct. Mol. Biol. 2004, 11, 412–419. [Google Scholar] [CrossRef]
- Khersonsky, O.; Tawfik, D.S. The Histidine 115-Histidine 134 Dyad Mediates the Lactonase Activity of Mammalian Serum Paraoxonases. J. Biol. Chem. 2006, 281, 7649–7656. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Billecke, S.; Sorenson, R.; Bisgaier, C.; Newton, R.; Rosenblat, M.; Erogul, J.; Hsu, C.; Dunlop, C.; La Du, B. Paraoxonase Active Site Required for Protection against LDL Oxidation Involves Its Free Sulfhydryl Group and Is Different from That Required for Its Arylesterase/Paraoxonase Activities: Selective Action of Human Paraoxonase Allozymes Q and R. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Evelson, P.; Gambino, G.; Travacio, M.; Jaita, G.; Verona, J.; Maroncelli, C.; Wikinski, R.; Llesuy, S.; Brites, F. Higher Antioxidant Defences in Plasma and Low Density Lipoproteins from Rugby Players. Eur. J. Clin. Investig. 2002, 32, 818–825. [Google Scholar] [CrossRef]
- Richter, B.; Niessner, A.; Penka, M.; Grdić, M.; Steiner, S.; Strasser, B.; Ziegler, S.; Zorn, G.; Maurer, G.; Simeon-Rudolf, V.; et al. Endurance Training Reduces Circulating Asymmetric Dimethylarginine and Myeloperoxidase Levels in Persons at Risk of Coronary Events. Thromb. Haemost. 2005, 94, 1306–1311. [Google Scholar] [CrossRef]
- Brites, F.; Zago, V.; Verona, J.; Muzzio, M.L.; Wikinski, R.; Schreier, L. HDL Capacity to Inhibit LDL Oxidation in Well-Trained Triathletes. Life Sci. 2006, 78, 3074–3081. [Google Scholar] [CrossRef]
- Myers, J.; Buchanan, N.; Walsh, D.; Kraemer, M.; McAuley, P.; Hamilton-Wessler, M.; Froelicher, V.F. Comparison of the ramp versus standard exercise protocols. J. Am. Coll. Cardiol. 1991, 17, 1334–1342. [Google Scholar] [CrossRef] [Green Version]
- Stawski, R.; Walczak, K.; Kosielski, P.; Meissner, P.; Budlewski, T.; Padula, G.; Nowak, D. Repeated Bouts of Exhaustive Exercise Increase Circulating Cell Free Nuclear and Mitochondrial DNA without Development of Tolerance in Healthy Men. PLoS ONE 2017, 12, e0178216. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, M.; Takanami, Y.; Maruyama, T.; Murata, M.; Motohashi, Y.; Nakano, S.; Uchida, K.; Maruyama, C.; Kyotani, S.; Tsushima, M. The Ratio of Serum Paraoxonase/Arylesterase Activity Using an Improved Assay for Arylesterase Activity to Discriminate PON1(R192) from PON1(Q192). J. Atheroscler. Thromb. 2003, 10, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peternelj, T.T.; Coombes, J.S. Antioxidant Supplementation during Exercise Training: Beneficial or Detrimental? Sport. Med. 2011, 41, 1043–1069. [Google Scholar] [CrossRef] [PubMed]
- Oda, M.N.; Bielicki, J.K.; Berger, T.; Forte, T.M. Cysteine Substitutions in Apolipoprotein A-I Primary Structure Modulate Paraoxonase Activity. Biochemistry 2001, 40, 1710–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblat, M.; Karry, R.; Aviram, M. Paraoxonase 1 (PON1) Is a More Potent Antioxidant and Stimulant of Macrophage Cholesterol Efflux, When Present in HDL than in Lipoprotein-Deficient Serum: Relevance to Diabetes. Atherosclerosis 2006, 187, 74.e1–74.e10. [Google Scholar] [CrossRef]
- Dullaart, R.P.F.; Kwakernaak, A.J.; Dallinga-Thie, G.M. The Positive Relationship of Serum Paraoxonase-1 Activity with Apolipoprotein E Is Abrogated in Metabolic Syndrome. Atherosclerosis 2013, 230, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Blatter Garin, M.C.; James, R.W.; Dussoix, P.; Blanché, H.; Passa, P.; Froguel, P.; Ruiz, J. Paraoxonase Polymorphism Met-Leu54 Is Associated with Modified Serum Concentrations of the Enzyme. A Possible Link between the Paraoxonase Gene and Increased Risk of Cardiovascular Disease in Diabetes. J. Clin. Investig. 1997, 99, 62–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bełtowski, J.; Wójcicka, G.; Mydlarczyk, M.; Jamroz, A. Cerivastatin Modulates Plasma Paraoxonase/Arylesterase Activity and Oxidant-Antioxidant Balance in the Rat. Pol. J. Pharmacol. 2002, 54, 143–150. [Google Scholar] [PubMed]
- Gan, K.N.; Smolen, A.; Eckerson, H.W.; La Du, B.N. Purification of Human Serum Paraoxonase/Arylesterase. Evidence for One Esterase Catalyzing Both Activities. Drug Metab. Dispos. 1991, 19, 100–106. [Google Scholar] [PubMed]
- Arslan, C.; Gulcu, F.; Gursu, M.F. Effects of Oxidative Stress Caused by Acute and Regular Exercise on Levels of Some Serum Metabolites and the Activities of Paraoxonase and Arylesterase. Biol. Sport 2005, 22, 375–383. [Google Scholar]
- Rosenblat, M.; Gaidukov, L.; Khersonsky, O.; Vaya, J.; Oren, R.; Tawfik, D.S.; Aviram, M. The Catalytic Histidine Dyad of High Density Lipoprotein-Associated Serum Paraoxonase-1 (PON1) Is Essential for PON1-Mediated Inhibition of Low Density Lipoprotein Oxidation and Stimulation of Macrophage Cholesterol Efflux. J. Biol. Chem. 2006, 281, 7657–7665. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Hardak, E.; Vaya, J.; Mahmood, S.; Milo, S.; Hoffman, A.; Billicke, S.; Draganov, D.; Rosenblat, M. Human Serum Paraoxonases (PON1) Q and R Selectively Decrease Lipid Peroxides in Human Coronary and Carotid Atherosclerotic Lesions: PON1 Esterase and Peroxidase-like Activities. Circulation 2000, 101, 2510–2517. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Rosenblat, M.; Billecke, S.; Erogul, J.; Sorenson, R.; Bisgaier, C.L.; Newton, R.S.; La Du, B. Human Serum Paraoxonase (PON 1) Is Inactivated by Oxidized Low Density Lipoprotein and Preserved by Antioxidants. Free Radic. Biol. Med. 1999, 26, 892–904. [Google Scholar] [CrossRef]
- Abuhandan, M.; Cakmak, A.; Taskin, A.; Karakaya, E.; Kocyigit, A.; Kilic, H. The Effect of Age on the Paraoxonase and Arylesterase Activity of Pregnant Mothers and Their Infants. J. Clin. Lab. Anal. 2012, 26, 302–306. [Google Scholar] [CrossRef]
- Allen, J.; Sun, Y.; Woods, J.A. Exercise and the Regulation of Inflammatory Responses. Prog. Mol. Biol. Transl. Sci. 2015, 135, 337–354. [Google Scholar] [CrossRef]
- Fatouros, I.G.; Destouni, A.; Margonis, K.; Jamurtas, A.Z.; Vrettou, C.; Kouretas, D.; Mastorakos, G.; Mitrakou, A.; Taxildaris, K.; Kanavakis, E.; et al. Cell-Free Plasma DNA as a Novel Marker of Aseptic Inflammation Severity Related to Exercise Overtraining. Clin. Chem. 2006, 52, 1820–1824. [Google Scholar] [CrossRef] [Green Version]
- Draganidis, D.; Chatzinikolaou, A.; Jamurtas, A.Z.; Carlos Barbero, J.; Tsoukas, D.; Theodorou, A.S.; Margonis, K.; Michailidis, Y.; Avloniti, A.; Theodorou, A.; et al. The Time-Frame of Acute Resistance Exercise Effects on Football Skill Performance: The Impact of Exercise Intensity. J. Sport. Sci. 2013, 31, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Dullaart, R.P.F.; De Vries, R.; Sluiter, W.J.; Voorbij, H.A.M. High Plasma C-Reactive Protein (CRP) Is Related to Low Paraoxonase-I (PON-I) Activity Independently of High Leptin and Low Adiponectin in Type 2 Diabetes Mellitus. Clin. Endocrinol. Oxf. 2009, 70, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Kannampuzha, J.; Darling, P.B.; Maguire, G.F.; Donnelly, S.; McFarlane, P.; Chan, C.T.; Connelly, P.W. Paraoxonase 1 Arylesterase Activity and Mass Are Reduced and Inversely Related to C-Reactive Protein in Patients on Either Standard or Home Nocturnal Hemodialysis. Clin. Nephrol. 2010, 73, 131–138. [Google Scholar]
- Gugliucci, A.; Kotani, K.; Kimura, S. Paraoxonase 1 in Chronic Kidney Failure. J. Lipids 2012, 2012, 726048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzystek-Korpacka, M.; Patryn, E.; Hotowy, K.; Czapińska, E.; Majda, J.; Kustrzeba-Wójcicka, I.; Noczyńska, A.; Gamian, A. Paraoxonase-1 Activity in Overweight and Obese Children and Adolescents: Association with Obesity-Related Inflammation and Oxidative Stress. Adv. Clin. Exp. Med. 2013, 22, 229–236. [Google Scholar]
- Bains, Y.; Caccavello, R.; Kotani, K.; Gugliucci, A. Paraoxonase 1, HDL Subclasses and Post Surgery Acute Inflammation: A Pilot Study. Antioxidants 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahrach, H.; Ghalim, N.; Taki, H.; Kettani, A.; Er-Rachdi, L.; Ramdani, B.; Saïle, R. Serum Paraoxonase Activity, High-Sensitivity C-Reactive Protein, and Lipoprotein Disturbances in End-Stage Renal Disease Patients on Long-Term Hemodialysis. J. Clin. Lipidol. 2008, 2, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, É.; Marinho, D.A.; Neiva, H.P.; Lourenço, O. Inflammatory Effects of High and Moderate Intensity Exercise-A Systematic Review. Front. Physiol. 2020, 10, 1055. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, A.; Hauth, M.; Walter, M.; Hudemann, J.; Wank, V.; Niess, A.M.; Northoff, H. Exhaustive Exercise Modifies Different Gene Expression Profiles and Pathways in LPS-Stimulated and Un-Stimulated Whole Blood Cultures. Brain. Behav. Immun. 2014, 39, 130–141. [Google Scholar] [CrossRef]
- Magal, M.; Dumke, C.L.; Urbiztondo, Z.G.; Cavill, M.J.; Triplett, N.T.; Quindry, J.C.; McBride, J.M.; Epstein, Y. Relationship between Serum Creatine Kinase Activity Following Exercise-Induced Muscle Damage and Muscle Fibre Composition. J. Sport. Sci. 2010, 28, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Eraldemir, F.C.; Korak, T. Paraoxonases, Oxidative Stress, and Breast Cancer. In Cancer Oxidative Stress Diet. Antioxidants; Academic Press: Cambridge, MA, USA, 2021; pp. 3–14. [Google Scholar] [CrossRef]
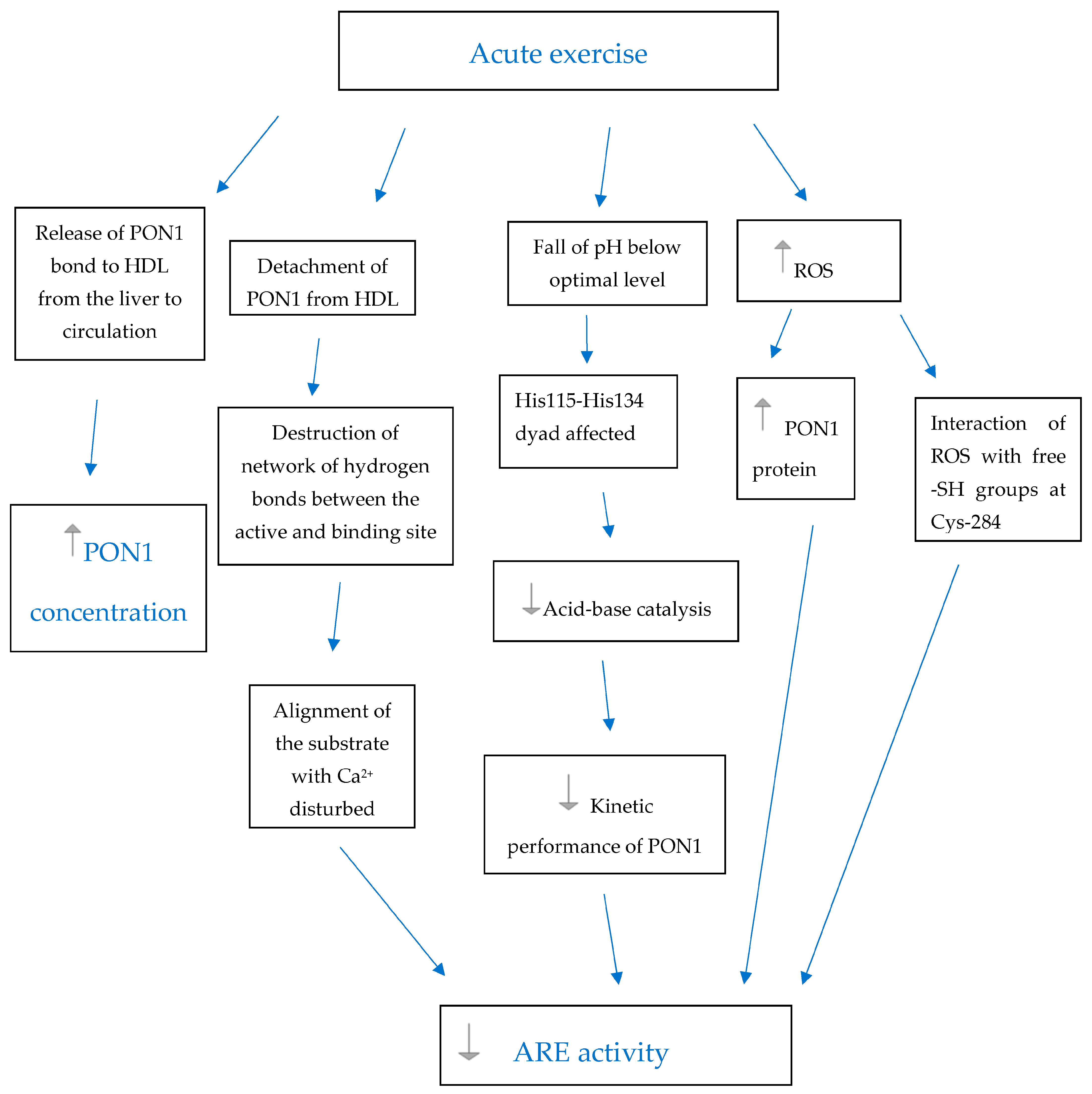
 -increase,
-increase,  - decrease, PON1—paraoxonase 1, HDL—high-density lipoprotein, ROS—reactive oxygen species, Ca2+—catalytic calcium ion in the active site tunnel of PON1, His—Histidine, Cys—Cysteine, -SH groups—sulfhydryl groups.
-increase, - decrease, PON1—paraoxonase 1, HDL—high-density lipoprotein, ROS—reactive oxygen species, Ca2+—catalytic calcium ion in the active site tunnel of PON1, His—Histidine, Cys—Cysteine, -SH groups—sulfhydryl groups.
- decrease, PON1—paraoxonase 1, HDL—high-density lipoprotein, ROS—reactive oxygen species, Ca2+—catalytic calcium ion in the active site tunnel of PON1, His—Histidine, Cys—Cysteine, -SH groups—sulfhydryl groups.
-increase, - decrease, PON1—paraoxonase 1, HDL—high-density lipoprotein, ROS—reactive oxygen species, Ca2+—catalytic calcium ion in the active site tunnel of PON1, His—Histidine, Cys—Cysteine, -SH groups—sulfhydryl groups.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participants Characteristics (N = 11) | |
|---|---|
| Age (years) | 34.0 ± 5.2 |
| Gender | male |
| Race | Caucasian |
| Training experience (years) | 10–15 |
| Body mass (kg) | 87.4 ±13.8 |
| BMI (kg/m2) | 26.2 ± 3.1 |
| VO2max (ml/kg/min) | 49.6 ± 4.5 |
| Resting HR (beats/min) | 72 ± 11 |
| Resting BP (mmHg) | 127 ± 6/80 ± 4 |
| FVC (L) | 6.09 ± 0.41 |
| % of predicted FEV1 | 106.4 ± 6.4 |
| FEV1/FVC (%) | 80.9 ± 5.6 |
| Parameter | Exercise Bout | ||
|---|---|---|---|
| 1st Bout | 2nd Bout | 3rd Bout | |
| HRmax (beats/min) | 184 ± 10 | 183 ± 11 | 176 ± 12 *^ |
| Post-exercise BP (mmHg) | 172 ± 20/82 ± 11 | 169 ± 13/80 ± 10 | 166 ± 13/79 ± 11 |
| Running distance (km) | 8.6 ± 5.5 | 10.7 ± 7.6 * | 10.4 ± 7.2 * |
| Running time (min) | 47 ± 31 | 57 ± 41 * | 56 ± 40 * |
| Biochemical Measurements | First Exercise | Second Exercise | Third Exercise | |||
|---|---|---|---|---|---|---|
| Before | Bout | Before | Bout | Before | Bout | |
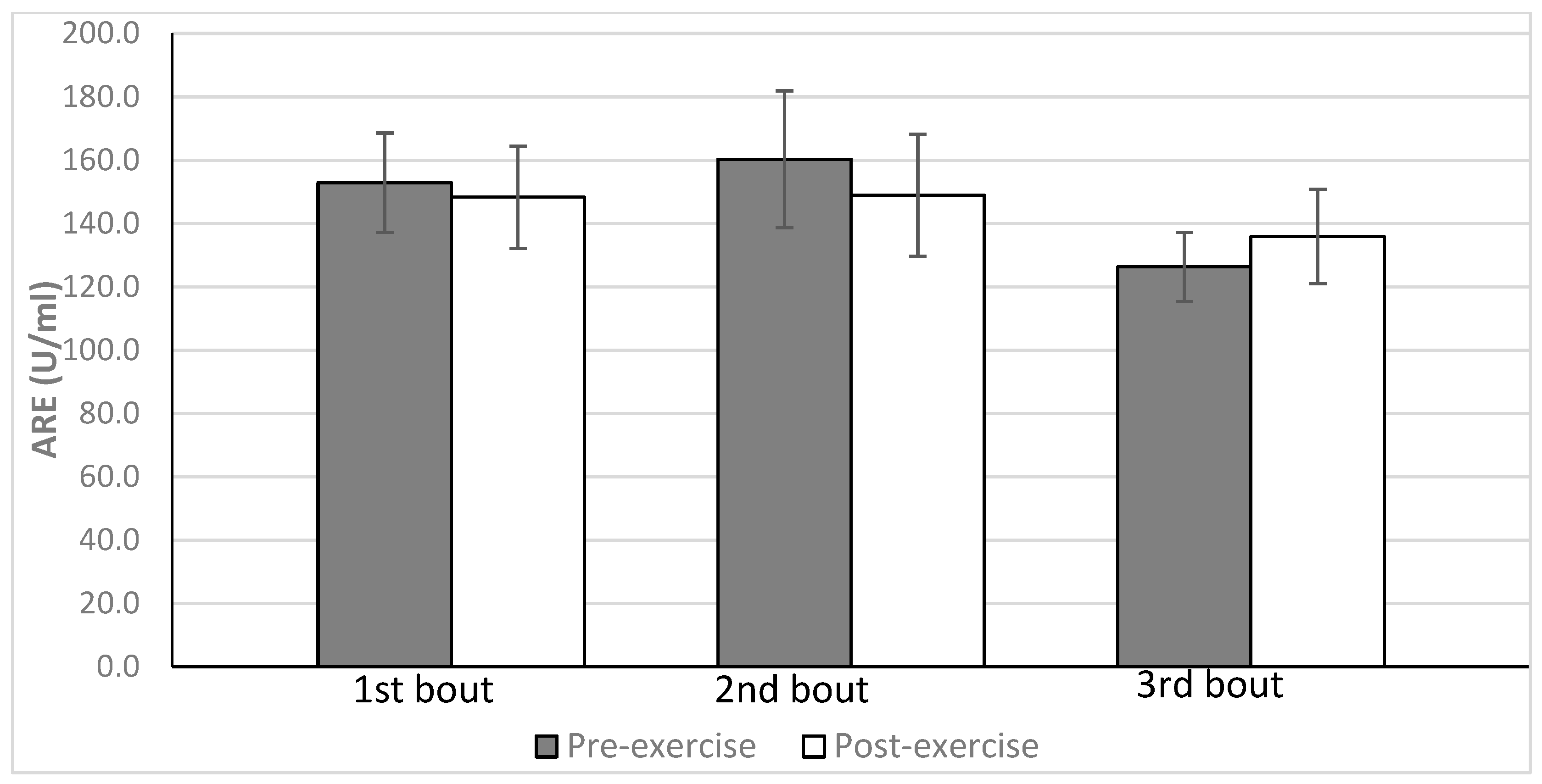
| Arylesterase activity (ARE) (U/mL) | 152.9 ± 15.7 | 148.3 ± 16.1 | 160.3 ± 21.6 | 148.9 ± 19.2 | 126.3 ± 10.9 | 135.9 ± 15 |
| Paraoxonase activity (PON) (U/L) | 884.4 ± 96.2 | 941.8 ± 93.1 * | 831 ± 83.8 | 914.6± 98.8 * | 858.2 ± 83.5 | 964 ± 86.8 * |
| Paraoxonase 1 concentration (PON1c) (µg/mL) | 1.62 ± 0.14 | 2.67 ± 0.2 * | 1.55 ± 0.11 | 2.99 ± 0.25 * | 1.61 ± 0.1 | 2.85 ± 0.26 * |
| Arylesterase activity/paraoxonase 1 concentration (ARE/PON1c) | 100.5 ± 37.8 | 60.9 ± 29.9 * | 111.7 ± 68.3 | 54.8 ± 29.3 * | 81.5 ± 28.5 | 54.8 ± 32.8 * |
| Ferric-reducing activity of plasma (FRAP) (mM/L Fe+2) | 1.15 ± 0.06 | 1.34 ± 0.07 * | 1.07 ± 0.05 | 1.29 ± 0.06 * | 1.18 ± 0.05 | 1.32 ± 0.04 * |
| High-density lipoprotein (HDL) (mM/L) | 1.34 ± 0.05 | 1.45 ± 0.05 *# | 1.29 ± 0.05 | 1.39 ± 0.06 * | 1.32 ± 0.04 | 1.4 ± 0.05 * |
| C-reactive protein (CRP) (mg/L) | 0.99 ± 0.19 | 1.53 ± 0.41 * | 1.31 ± 0.35 | 1.29 ± 0.41 | 1.73 ± 0.56 | 1.92 ± 0.60 |
| Lactate (mM/L) | 1.72 ± 0.24 | 8.95 ± 1.46 * | 1.42 ± 0.21 | 8.83 ± 1.59 * | 1.56 ± 0.15 | 7.97 ± 1.42 * |
| Creatine kinase (CK) (U/L) | 162.2 ± 20.1 | 210.8 ± 35.5 * | 266.6 ± 68.4 | 301.8 ± 58.3 | 300.8 ± 41.7 | 348.4 ± 41.0 * |
| White blood cells (WBC) (×103/mm3) | 5.86 ± 0.62 | 9.45 ±1.94 * | 5.68 ± 0.59 | 9.99 ± 2.87 * | 5.76 ± 0.49 | 10.7 ± 3.24 * |
| polymorphonuclear leukocytes (PMN) (×103/mm3) | 3.98 ± 0.71 | 6.02 ± 1.88 * | 3.85 ± 0.68 | 6.46 ± 2.92 * | 3.97 ± 0.60 | 7.27 ± 3.49 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otocka-Kmiecik, A.; Orłowska-Majdak, M.; Stawski, R.; Szkudlarek, U.; Padula, G.; Gałczyński, S.; Nowak, D. Effect of Exercise Repetitions on Arylesterase Activity of PON1 in Plasma of Average-Trained Men—The Dissociation between Activity and Concentration. Antioxidants 2023, 12, 1296. https://doi.org/10.3390/antiox12061296
Otocka-Kmiecik A, Orłowska-Majdak M, Stawski R, Szkudlarek U, Padula G, Gałczyński S, Nowak D. Effect of Exercise Repetitions on Arylesterase Activity of PON1 in Plasma of Average-Trained Men—The Dissociation between Activity and Concentration. Antioxidants. 2023; 12(6):1296. https://doi.org/10.3390/antiox12061296
Chicago/Turabian StyleOtocka-Kmiecik, Aneta, Monika Orłowska-Majdak, Robert Stawski, Urszula Szkudlarek, Gianluca Padula, Szymon Gałczyński, and Dariusz Nowak. 2023. "Effect of Exercise Repetitions on Arylesterase Activity of PON1 in Plasma of Average-Trained Men—The Dissociation between Activity and Concentration" Antioxidants 12, no. 6: 1296. https://doi.org/10.3390/antiox12061296