


Barranca variabilis sp. nov.—A New Terrestrial Alga of the Genus Barranca (Chaetophorales, Chlorophyta) from the Baikal Region (Russia)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey Area and Alga Origin
2.2. Culture Conditions and Microscopic Observations
2.3. DNA Isolation, PCR Amplification, and Sequencing
2.4. Phylogenetic Reconstruction and Secondary Structure Analysis of ITS2
3. Results
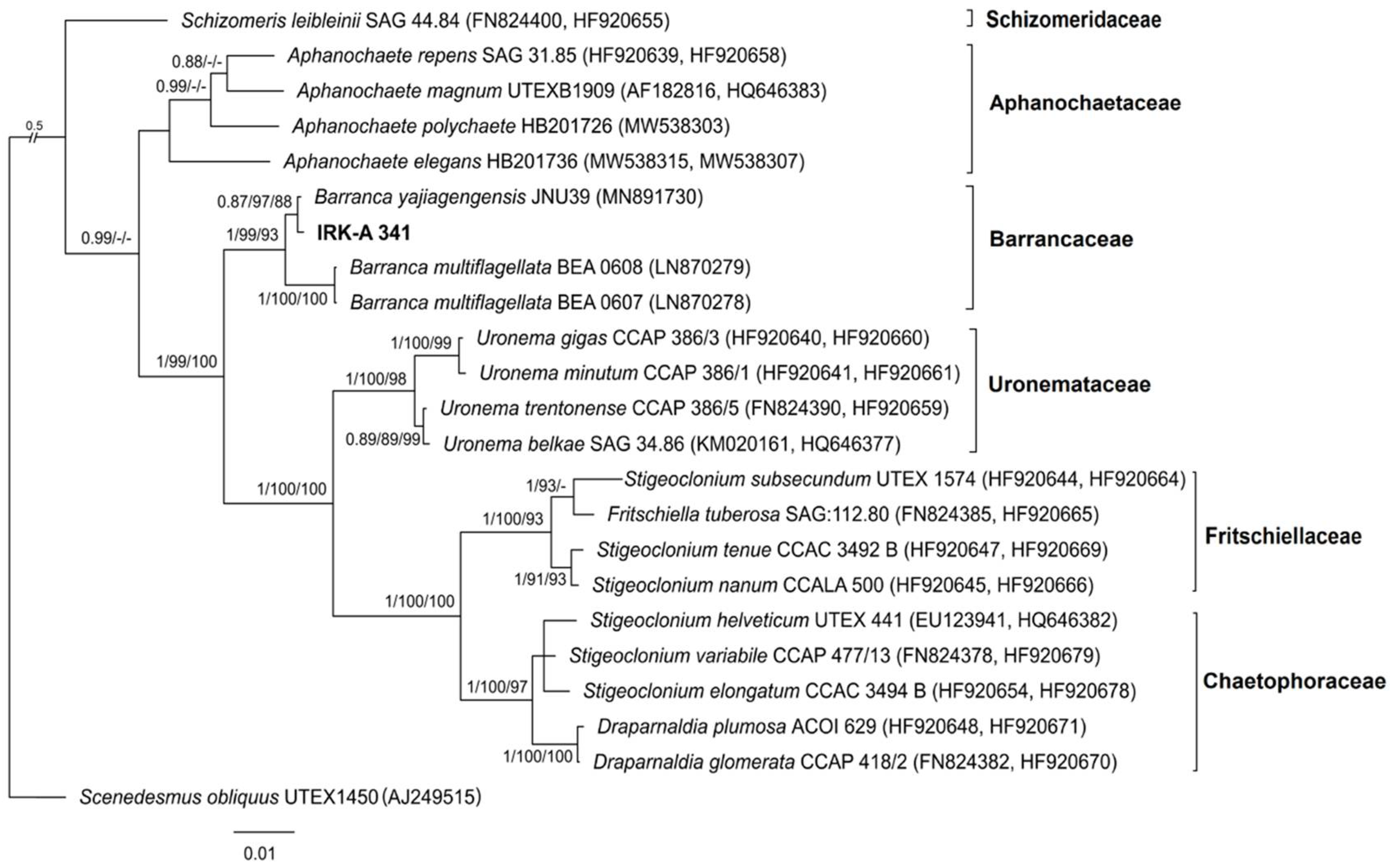
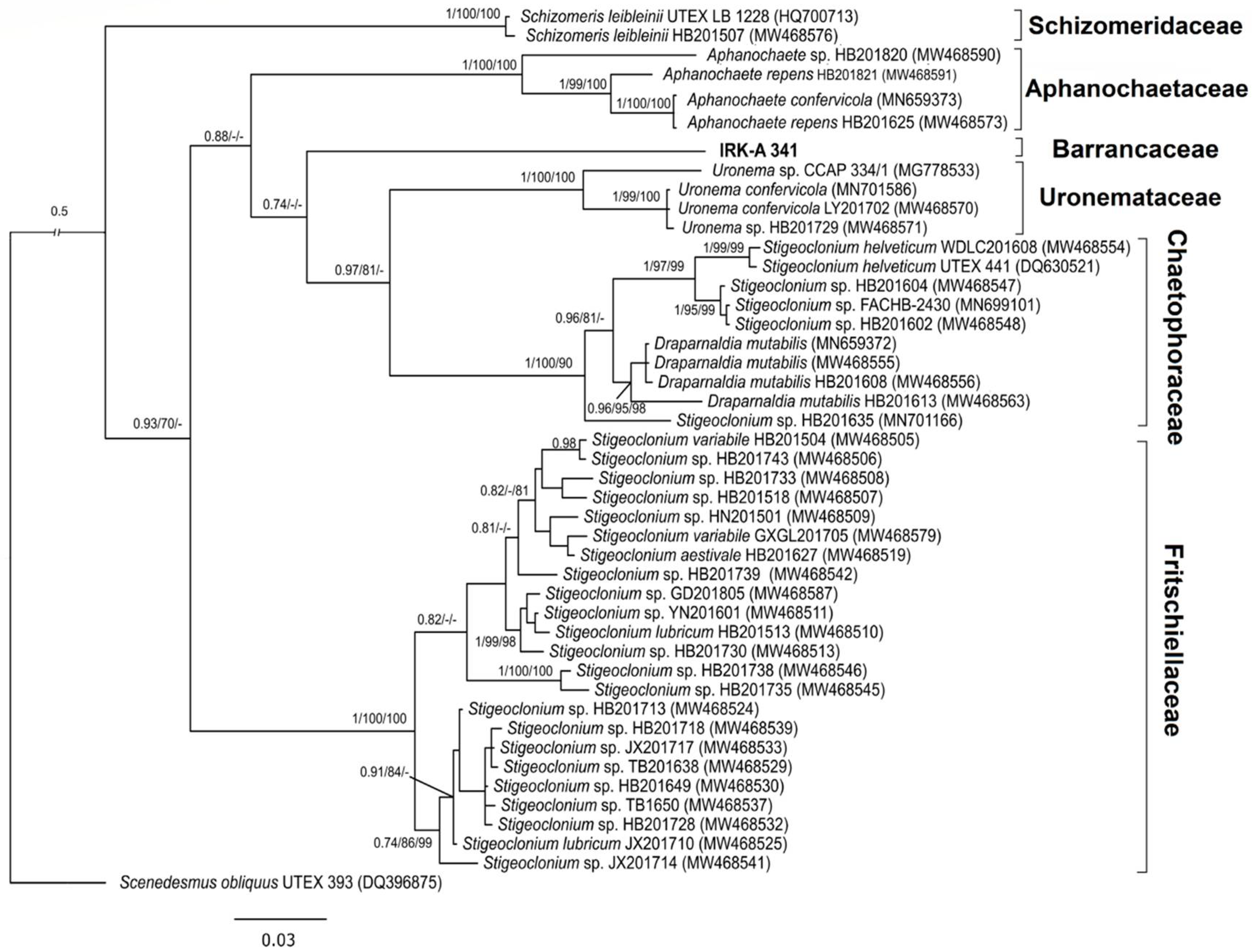
3.1. Phylogenetic Analysis
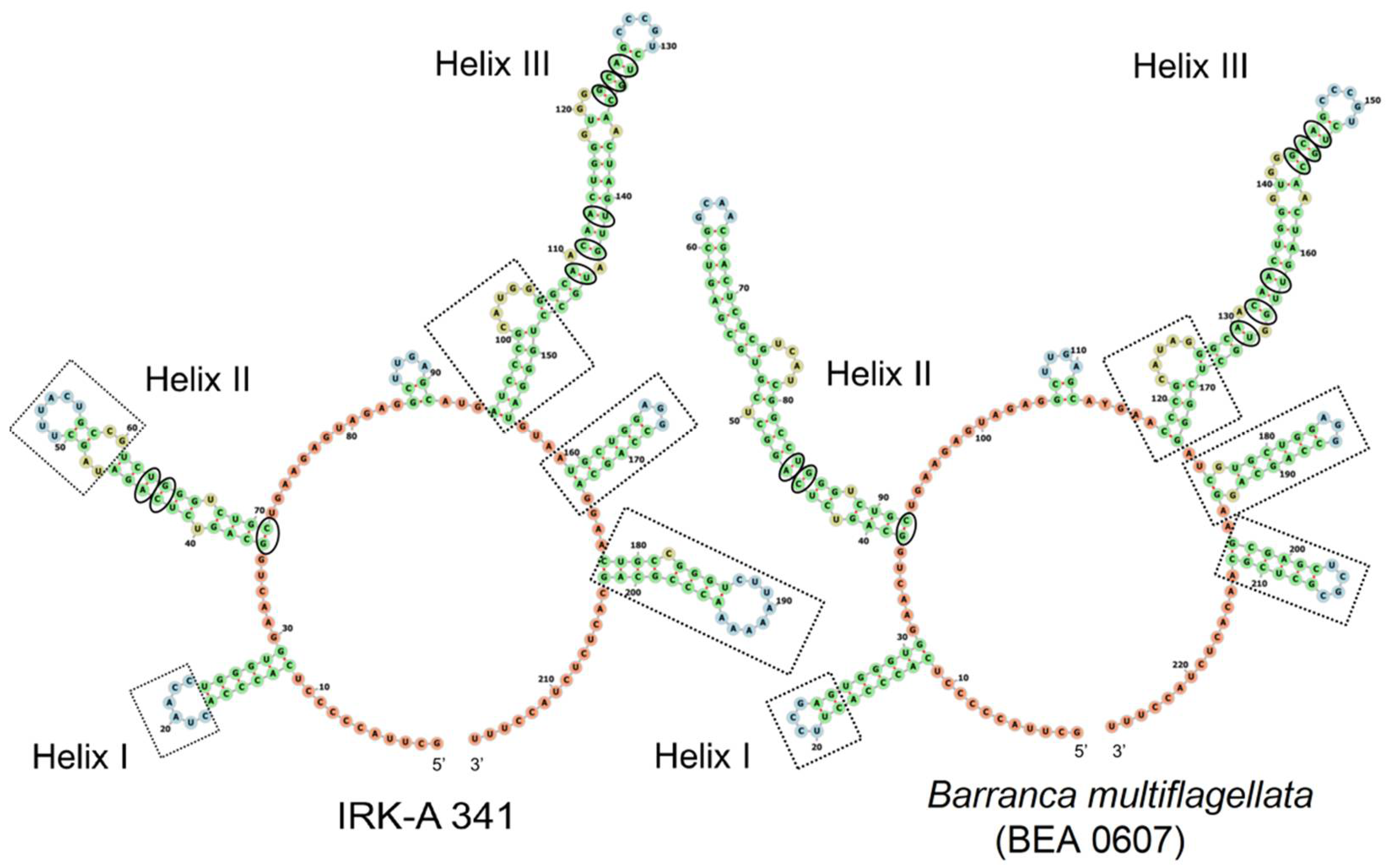
3.2. The Secondary Structure of ITS2
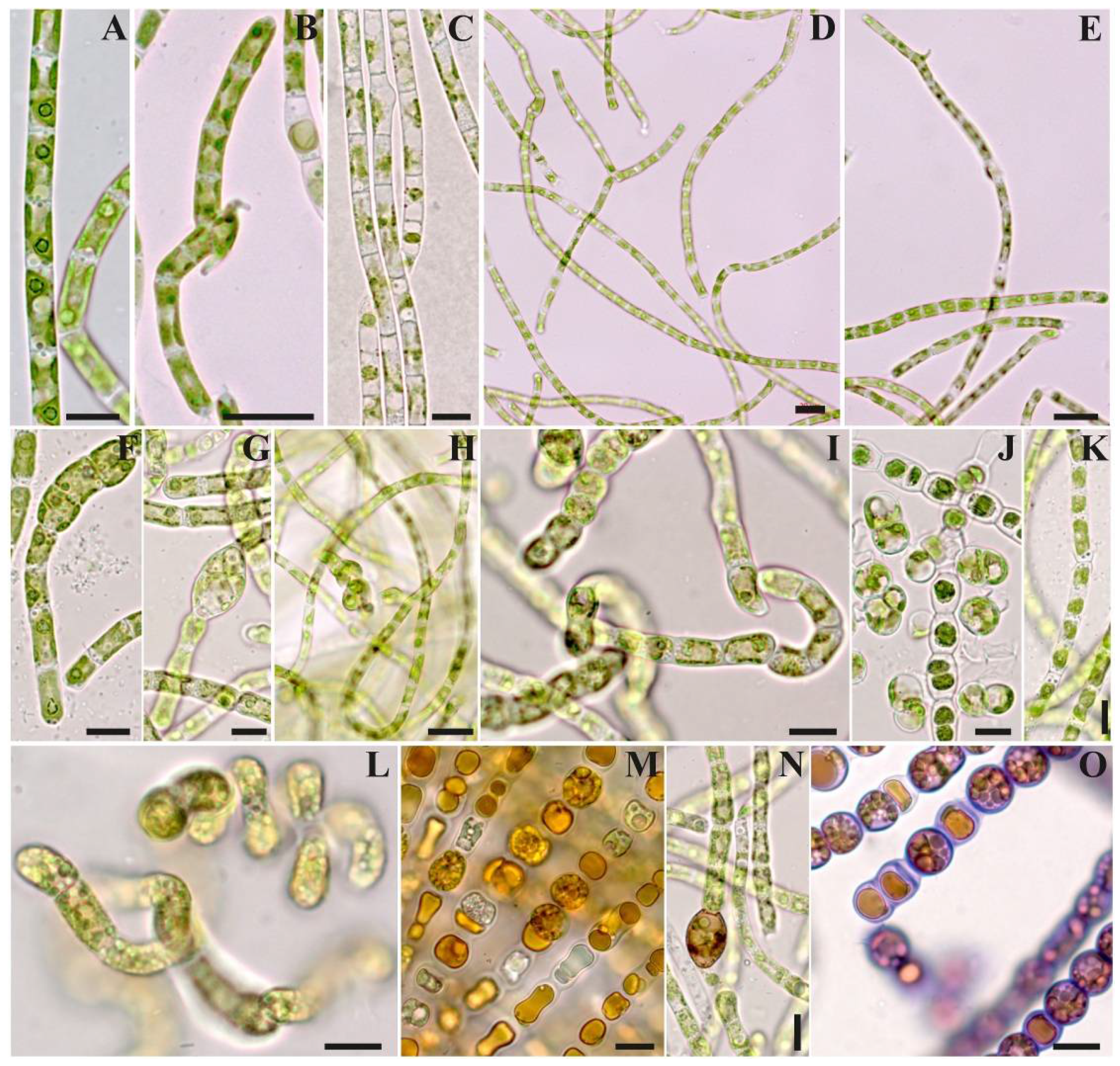
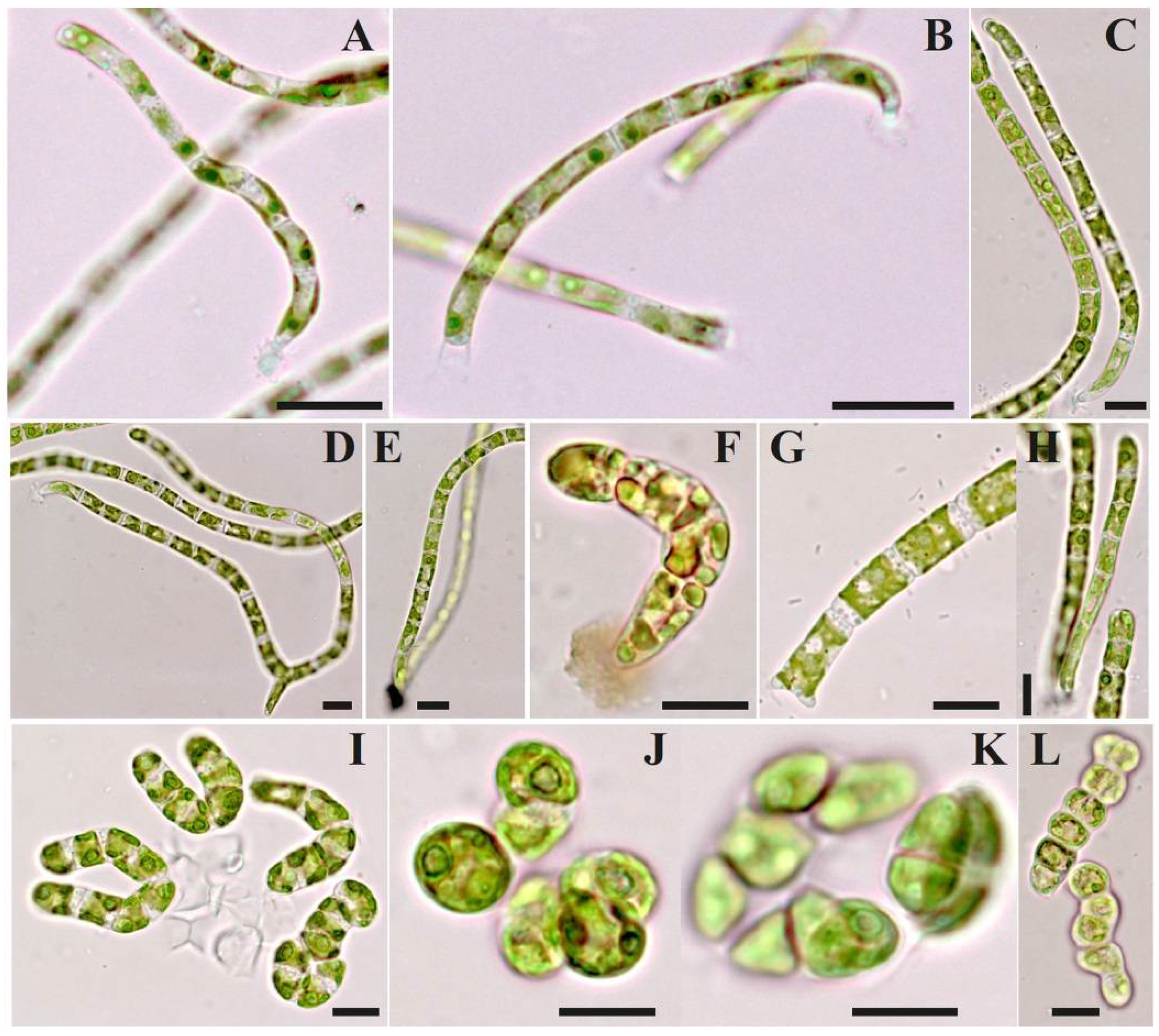
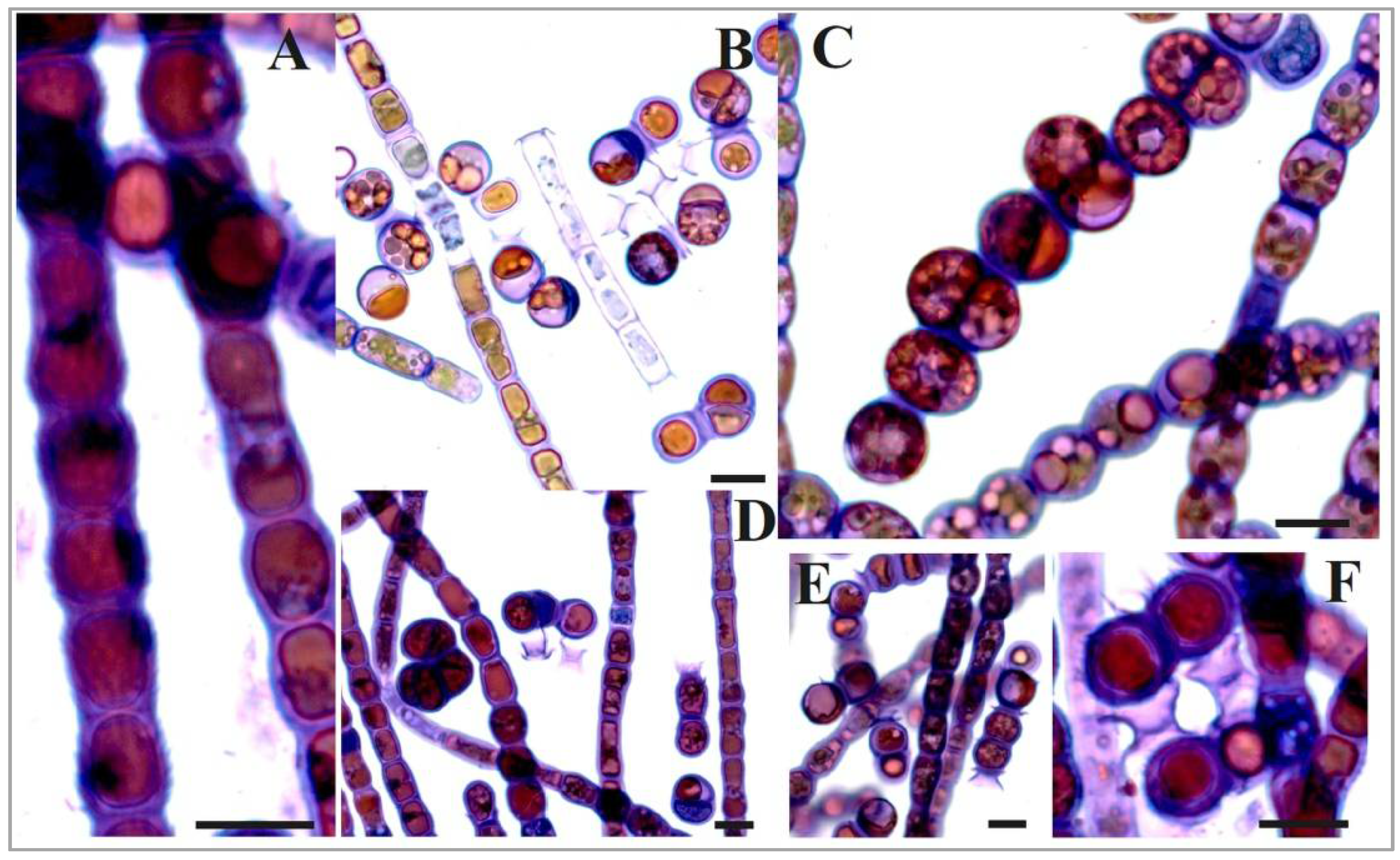
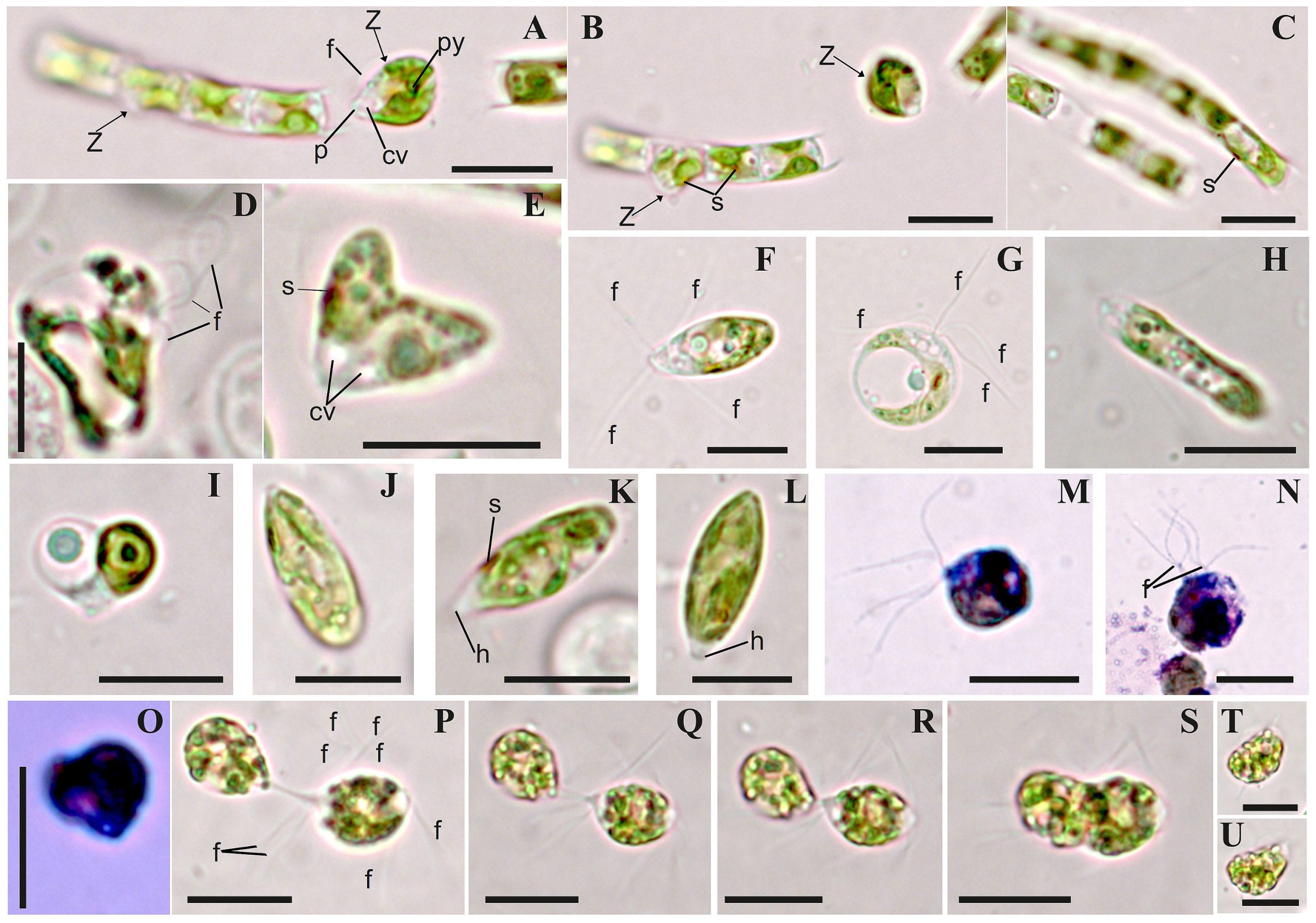
3.3. Morphology
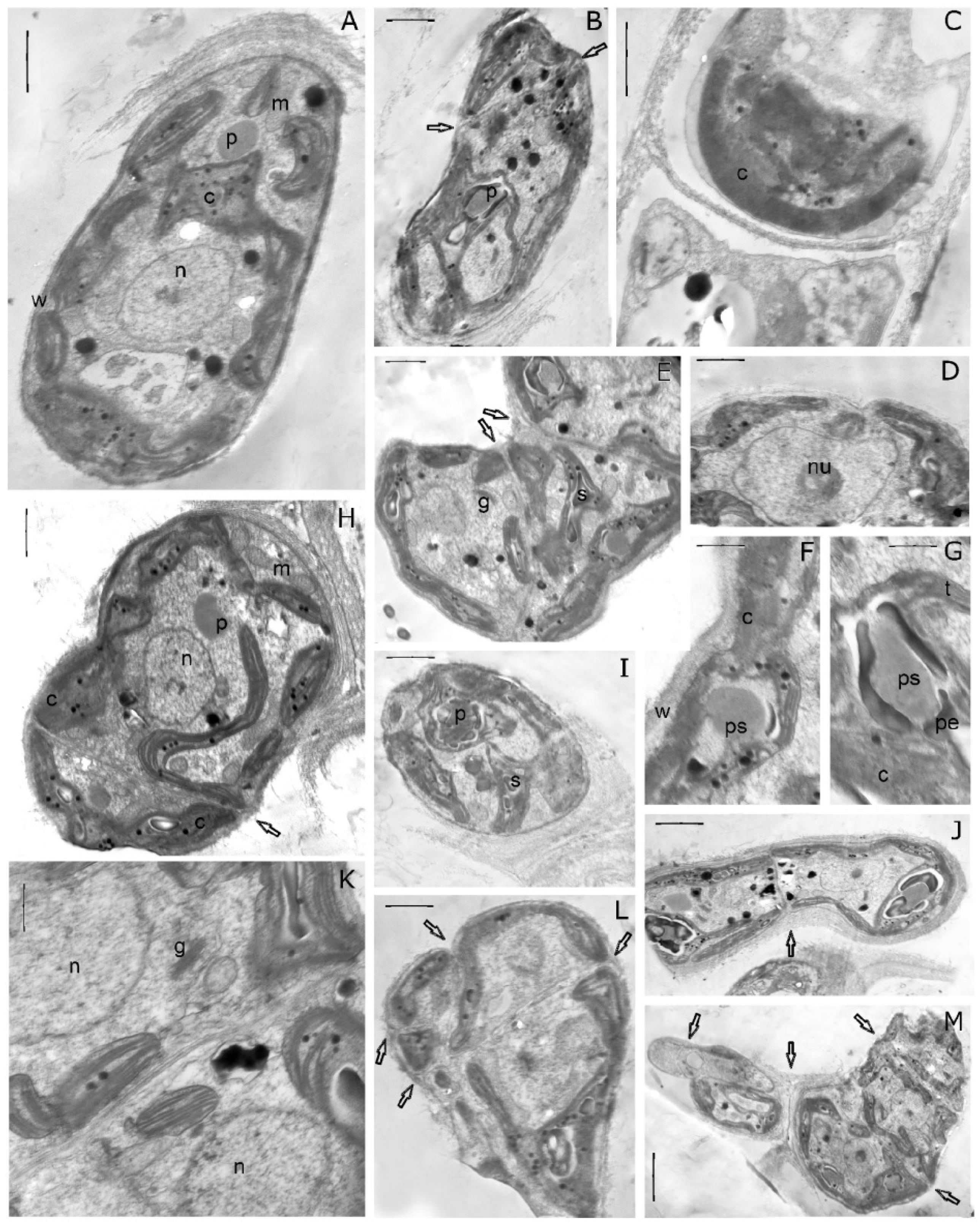
3.4. Ultrastructure
3.5. Taxonomic Conclusion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manton, I. Observations on the Fine Structure of the Zoospore and Young Germling of Stigeoclonium. J. Exp. Bot. 1964, 15, 399–411. Available online: http://www.jstor.org/stable/23686762 (accessed on 3 April 2023). [CrossRef]
- McBride, G.E. Cytokinesis in the Green Alga Fritschiella. Nature 1967, 216, 939. [Google Scholar] [CrossRef]
- Floyd, G.L.; Stewart, K.D.; Mattox, K.R. Comparative cytology of Ulothrix and Stigeoclonium. J. Phycol. 1972, 8, 68–81. [Google Scholar] [CrossRef]
- Stewart, K.D.; Mattox, K.R.; Floyd, G.L. Mitosys, cytokinesis, the distribution of plasmodesmata, and other cytological characteristics in the Ulotrichales, Ulvales and Chaetophorales: Phylogenetic and taxonomic considerations. J. Phycol. 1973, 9, 128–141. [Google Scholar] [CrossRef]
- Mattox, K.R.; Kenneth, D.; Stewart, K.D.; Floyd, G.L. The cytology and classification of Schizomeris leibleinii (Chlorophyceae). I. The vegetative thallus. Phycologia 1974, 13, 63–69. [Google Scholar] [CrossRef]
- Melkonian, M. The fine structure of the zoospores of Fritschiella tuberosa, Iyeng. (Chaetophorineae, Chlorophyceae), with special reference to the flagellar apparatus. Protoplasma 1975, 86, 391–404. [Google Scholar] [CrossRef]
- Lokhorst, G.M.; Bakker, M.E.; Star, W. Ultrastructure of Draparnaldia glomerata (Chaetophorales, Chlorophyceae) II. Mitosis and cytokinesis. Nord. J. Bot. 1984, 4, 553–562. [Google Scholar] [CrossRef]
- Mattox, K.R.; Stewart, K.D. Classification of the green algae: A concept based on comparative cytology. In Systematics of the Green Algae; John, D.M., Irvine, D.E.G., Eds.; Academic Press: New York, NY, USA, 1984; pp. 29–72. [Google Scholar]
- Watanabe, S.; Floyd, G.L. Ultrastructure of the Quadriflagellate Zoospores of the Filamentous Green Algae Chaetophora incrassata and Pseudoschizomeris caudata (Chaetophorales, Chlorophyceae) with Emphasis on the Flagellar Apparatus. Bot. Mag. Tokyo 1989, 102, 533–546. [Google Scholar] [CrossRef]
- Michetti, K.M.; Leonardi, P.I.; Cáceres, E.J. Morphology, cytology and taxonomic remarks of four species of Stigeoclonium (Chaetophorales, Chlorophyceae) from Argentina. Phycol. Res. 2010, 58, 35–43. [Google Scholar] [CrossRef]
- Caisová, L.; Melkonian, M. The Chaetophorales (Chlorophyceae)—A taxonomic revision at family level. Eur. J. Phycol. 2018, 53, 381–392. [Google Scholar] [CrossRef]
- Printz, H. Die Chaetophoralen der Binnengewässer. Hydrobiology 1964, XXIV, 1–376. [Google Scholar] [CrossRef]
- Cox, E.R.; Bold, H.C. Phycological Studies. VII. Taxonomic Investigations of Stigeoclonium. Univ. Texas Publ. 1966, 6618, 1–167. [Google Scholar]
- Caisová, L.; Reyes, C.P.; Álamo, V.C.; Quintana, A.M.; Surek, B.; Melkonian, M. Barrancaceae: A new green algal lineage with structural and behavioral adaptations to a fluctuating environment. Am. J. Bot. 2015, 102, 1482. [Google Scholar] [CrossRef] [PubMed]
- Moshkova, N.; Gollerbakh, M.M. Green Algae. Ulotrichophyceae. In Identification Book of Freshwater Algae of the USSR; Iss. 10; Nauka: Leningrad, Russia, 1986; 360p. [Google Scholar]
- Darienko, T.; Pröschold, T. Towards a monograph of non-marine Ulvophyceae using an integrative approach (Molecular phylogeny and systematics of terrestrial Ulvophyceae II). Phytotaxa 2017, 324, 1–41. [Google Scholar] [CrossRef]
- Škaloud, P.; Rindi, F.; Boedeker, C.; Leliaert, F. Chlorophyta: Ulvophyceae. In Freshwater Flora of Central Europe; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum: Berlin, Germany, 2018; Volume 13. [Google Scholar] [CrossRef]
- Caisová, L.; Marin, B.; Sausen, N.; Proschold, T.; Melkonian, M. Polyphyly of Chaetophora and Stigeoclonium within the Chaetophorales (Chlorophyceae), revealed by equence comparisons of nuclear-encoded SSU rRNA genes. J. Phycol. 2011, 47, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, X.; Wang, Q.; Hu, Z.; Liu, G. Reassessment of the species Stigeoclonium polyrhizum (Chaetophoraceae, Chaetophorales) based on morphological and molecular data. Phytotaxa 2018, 333, 086–098. [Google Scholar] [CrossRef]
- Liu, B.W.; Xiong, Q.; Liu, X.D.; Liu, G.X.; Hu, Z.Y. Molecular phylogeny and taxonomy of the genus Chaetophora (Chlorophyceae, Chlorophyta), including descriptions of Chaetophoropsis aershanensis gen. et sp. nov. J. Phycol. 2019, 55, 74–83. [Google Scholar] [CrossRef]
- Liu, B.; Hu, Y.; Hu, Z.; Liu, G.; Zhu, H. Taxonomic scheme of the order Chaetophorales (Chlorophyceae, Chlorophyta) based on chloroplast genomes. BMC Genom. 2020, 21, 442. [Google Scholar] [CrossRef]
- Brouard, J.-S.; Otis, C.; Lemieux, C.; Turme, M. The Chloroplast Genome of the Green Alga Schizomeris leibleinii (Chlorophyceae) Provides Evidence for Bidirectional DNA Replication from a Single Origin in the Chaetophorales. Genome Biol. Evol. 2011, 3, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Buchheim, M.A.; Sutherland, D.M.; Schleicher, T.; Förster, F.; Wolf, M. Phylogeny of Oedogoniales, Chaetophorales and Chaetopeltidales (Chlorophyceae): Inferences from sequence-structure analysis of ITS2. Ann. Bot. 2012, 109, 109–116. [Google Scholar] [CrossRef]
- Fučiková, K.; Lewis, P.O.; Neupane, S.; Karol, K.G.; Lewis, L.A. Order, please! Uncertainty in the ordinal-level classification of Chlorophyceae. PeerJ. 2019, 7, e6899. [Google Scholar] [CrossRef]
- Liu, B.; Zhu, H.; Dong, X.; Yan, Q.; Liu, G.; Hu, Z. Reassessment of suitable markers for taxonomy of Chaetophorales (Chlorophyceae, Chlorophyta) based on chloroplast genomes. Eukaryot. Microbiol. 2021, 68, e12858. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 27 February 2023).
- Iyengar, M.O.P. Fritschiella, a new terrestrial member of the Chaetophoraceae. New Phytol. 1932, 31, 329–335. [Google Scholar] [CrossRef]
- Ettl, H.; Gärtner, G. Syllabus der Boden-, Luft- und Flechtenalgen; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–773. [Google Scholar] [CrossRef]
- Caisová, L. Draparnaldia: A chlorophyte model for comparative analyses of plant terrestrialization. J. Exp. Bot. 2020, 71, 3305–3313. [Google Scholar] [CrossRef]
- Meyer, K.I. Einführung in die Algenflora des Baikalsees. Bull. Société Nat. Moscou. Sect. Biol. 1930, 39, 201–243. [Google Scholar]
- Izhboldina, L.A. Guide and Key to Benthonic and Periphyton Algae of Lake Baikal (meio- and macrophytes) with Short Notes on Their Ecology; Nauka-Center: Novosibirsk, Russia, 2007; pp. 1–248. (In Russian) [Google Scholar]
- Kozhov, M.M. Biology of Lake Baikal; AN USSR Publ.: Moscow, Russia, 1962; pp. 1–315. [Google Scholar] [CrossRef]
- Rosemond, A.D.; Browley, S.H. Species-specific characteristics explain the persistence of Stigeoclonium tenue (Chlorophyta) in a woodland stream. J. Phycol. 1996, 32, 54–63. [Google Scholar] [CrossRef]
- Izhboldina, L.A. Meio- and Benthonic Macrophytes of Lake Baikal (Algae); Batrayeva, A.A., Ed.; Irkutsk State University Press: Irkutsk, Russia, 1990; pp. 1–176. [Google Scholar]
- Timoshkin, O.A.; Vishnyakov, V.S.; Volkova, E.A.; Shirokaya, A.A.; Kulikova, N.N.; Zaytseva, E.P.; Lukhnev, A.G.; Popova, O.V.; Tomberg, I.V.; Potapskaya, N.V.; et al. Biology of the coastal zone of Lake Baikal 2. Accumulated material on the lake shore (splash zone): Classification, seasonal dynamics. Izv. Irkutsk. Gos. Universiteta. Seriya Biologiya. Ecol. 2012, 5, 40–91. [Google Scholar]
- Rusinek, O.T.; Takhteev, V.V.; Khodzher, T.V.; Pleshanov, A.S.; Voronin, V.I.; Arov, I.V.; Azovskii, M.G.; Goryunova, O.I.; Dryukker, V.V.; Zadonina, N.V.; et al. Baicalogy. 2 Books. Book 2; Rusinek, O.T., Takhteev, V.V., Gladkochub, D.P., Khodzer, T.V., Budnev, N.M., Eds.; Nauka: Novosibirsk, Russia, 2012; pp. 1–644. [Google Scholar]
- Kuklin, A.P.; Tsybekmitova, G.T.s.; Gorlacheva, E.P. Status of aquatic ecosystems lakes Onon-Torei plain for the years 1983–2011 (Eastern Transbaikalia). Arid Ecosyst. 2013, 19, 12–22. [Google Scholar] [CrossRef]
- Krupek, R.A.; Branco, C.C.Z. The influence of habitat structure, at different spatial scales, on the ecological distribution of macroalgal communities in streams. Braz. J. Bot. 2016, 39, 547–558. [Google Scholar] [CrossRef]
- Kulikova, N.N.; Paradina, L.F.; Suturin, A.N.; Tanicheva, I.V.; Izhboldina, L.A.; Khanaev, I.V.; Timoshkin, O.A. Trace element composition of all-the-year-round vegetating macroalgae on the stony littoral of lake Baikal (Russia). Algologia 2008, 18, 244–255. [Google Scholar]
- Kulikova, N.N.; Chebykin, E.P.; Volkova, E.A.; Bondarenko, N.A.; Vodneva, E.N.; Suturin, A.N. Determination of the element composition of benthic macro-algae for the indication of water quality of the shallow zone of the Listvennichnyi bay (South Baikal). Int. Res. J. 2017, 12, 166–176. [Google Scholar]
- Timoshkin, O.A.; Bondarenko, N.A.; Volkova, E.A.; Tomberg, I.V.; Vishnyakov, V.S.; Malnik, V.V. Mass development of green filamentous algae of the genera Spirogyra Link and Stigeoclonium Kutz. (Chlorophyta) in the coastal zone of the Southern Baikal. Gidrobiol. Zhurnal (Hydrobiol. J.) 2014, 5, 15–26. [Google Scholar]
- Gao, B.; Dai, C.; Zhang, H.; Zhang, C. Evaluation of a novel oleaginous filamentous green alga, Barranca yajiagengensis (Chlorophyta, Chaetophorales) for biomass, lipids and pigments production. Algal Res. 2022, 64, 102681. [Google Scholar] [CrossRef]
- Zhao, W.; Cui, X.; Wang, Z.-Q.; Yao, R.; Chen, M.-D.; Gao, B.-Y.; Cheng, W.; Zhang, C.-W.; Niu, J. Effects of Barranca yajiagengensis Powder in the Diet of Trachinotus ovatus on the Growth Performance, Antioxidant Capacity, Immunity and Morphology of the Liver and Intestine. Antioxidants 2022, 11, 1220. [Google Scholar] [CrossRef]
- Harding, J.P.C.; Whitton, B.A. Resistance to zinc of Stigeoclonium tenue in the field and the laboratory. Br. Phycol. J. 1976, 11, 417–426. [Google Scholar] [CrossRef]
- Liu, J.Z.; Danneels, B.; Vanormelingen, P.; Vyvermanb, W. Nutrient removal from horticultural waste water benthic filamentous algae Klebsormidium sp., Stigeoclonium spp. and their communities: From laboratory flask to outdoor Algal Turf Scrubber (ATS). Water. Res. 2016, 92, 61–68. [Google Scholar] [CrossRef]
- Aleksakhina, T.I.; Shtina, E.A. Soil Algae in Forest Biogeocenoses; Gollerbakh, M.M., Ed.; Nauka: Moscow, Russia, 1984; pp. 1–150. [Google Scholar]
- Medvedeva, L.A.; Nikulina, T.V. Catalogue of Freshwater Algae of the Southern Part of the Russian Far East; Dalnauka: Vladivostok, Russia, 2014; 271p. [Google Scholar]
- Rundina, L.A. The Zygnematales of Russia (Chlorophyta: Zygnematophyceae); Nauka: St. Peterburg, Russia, 1998; pp. 1–346. [Google Scholar] [CrossRef]
- Perminova, G.N.; Gutishvili, I.S.; Kitayev, E.V. Soil algae of the plant communities of the Baikal nature reserve. In Vodorosli, Lishainiki, Griby i Mokhoobraznyye v Zapovednikakh RSFSR; Sbornik Nauchnykh Trudov TsNIL RSFSR: Moscow, Russia, 1985; pp. 17–26. [Google Scholar]
- Shushuyeva, M.G. Formation of algae groups on the dumps of coal mines in Kuzbass. In Prirodnyye Kompleksy Nizshikh Rastenii Zapadnoi Sibiri; Nauka, Siberian Department: Novosibirsk, Russia, 1977; pp. 57–85. [Google Scholar]
- Gvozdetskii, N.A.; Mikhailov, N.I. Physical Geography of the USSR; Asian part: Moscow, Russia, 1978; 448p. [Google Scholar]
- Engineering Geology; East Siberia: Moscow, Russia, 1977; Volume 3, 657p.
- Starr, R.C.; Zeikus, J.A. UTEX: The Culture Collection of Algae at the University of Texas at Austin. J. Phycol. 1993, 29, 1–106. [Google Scholar] [CrossRef]
- Shergina, O.V.; Mikhailova, T.A.; Kalugina, O.V. Change of biogeochemical indexes in pine forests under technogenic pollution. Sib. Lesn. Zurnal (Sib. J. For. Sci.) 2018, 4, 29–38. [Google Scholar] [CrossRef]
- Egorova, I.N.; Tupikova, G.S.; Shergina, O.V.; Kazanovsky, S.G. Additional data about soil algae of steppe phytocenoses of the Predbaikalie. In Diversity of Soils and Biota of Northern and Central Asia; Proc. IV All-Russian Conference with International Participation; Buryat scientific center SB RAS Publ.: Ulan–Ude, Russia, 2021; pp. 153–155. [Google Scholar]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Gollerbakh, M.M.; Shtina, E.A. Soil Algae; Nauka: Leningrad, Russia, 1969; pp. 1–228. [Google Scholar]
- Egorova, I.N.; Kulakova, N.V.; Boldina, O.N. Amendments to the description of Chloromonas actinochloris (Chlorophyta) inferred from the study of the South Siberian finding. Bot. Pac. 2023, 12, 1–14. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 3–15. [Google Scholar] [CrossRef]
- Marin, B.; Palm, A.; Klingberg, M.; Melkonian, M. Phylogeny and taxonomic revision of plastid-containing euglenophytes based on SSU rDNA sequence comparisons and synapomorphic signatures in the SSU rRNA secondary structure. Protist 2003, 154, 99–145. [Google Scholar] [CrossRef]
- Fama, P.; Wysor, B.; Kooistra, W.H.C.F.; Zuccarello, G.C. Molecular phylogeny of the genus Caulerpa (Caulerpales, Chlorophyta) inferred from chloroplast tufA gene. J. Phycol. 2002, 38, 1040–1050. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Keller, A.; Schleicher, T.; Schultz, J.; Müller, T.; Dandekar, T.; Wolf, M. 5.8S–28S rRNA interaction and HMM-based ITS2 annotation. Gene 2009, 430, 50–57. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Caisová, L.; Marin, B.; Melkonian, M.A. consensus secondary structure of ITS2 in the Chlorophyta identified by phylogenetic reconstruction. Protist 2013, 164, 482. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.W. Pan-eukaryote ITS2 homologies revealed by RNA secondary structure. Nucleic Acids Res. 2007, 35, 3322–3329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tian, W.; Gao, Z.; Wang, G.; Zhao, H. Phylogenetic Utility of rRNA ITS2 Sequence-Structure under Functional Constraint. Int. J. Mol. Sci. 2020, 3, 6395. [Google Scholar] [CrossRef] [PubMed]
- Sudakova, E.A. Soil algae of meadow biogeocenoses. In Ekologiya Lugov Zapadnogo Uchastka Zony BAM; Nauka, Siberian department: Novosibirsk, Russia, 1986; pp. 35–44. [Google Scholar]
- Sudakova, E.A.; Egorova, I.N.; Maksimova, E.N.; Vysokikh, E.M. To the flora of the soil algae of the northern territories of Baikal region: Baikal-Patom and Stanovoi uplands (Russia). Probl. Bot. Yuzhnoi Sib. I Mong. 2020, 19, 254–259. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/ Trait | B. multiflagellata BEA 0607 a | B. yajiagengensis JNU39 b | B. variabilis IRK–A 341 c |
|---|---|---|---|
| 1. Filaments: | |||
| uniseriate | + | + | + |
| biseriate | – | – | + |
| branching | no or rarely simply | no or rarely simply | no or expressed, simply |
| polarity: a holdfast and a ring-like structure | + | + | + |
| 2. Vegetative cells: | |||
| shape of apical cells | straight, mostly rounded, slightly acute or acuminate | –//– | –//–, slightly expanded |
| shape of intercalary cells | cylindrical | –//– | –//–, frequently with a bulge on the middle, globose, dumbbell-, pear-, barrel-shaped, ovoid, square, wrong polygonal |
| length × width, µm | 4 − 2 × 4 − 13 | 4 − 30 × 4 − 11 | 2.3 − 32 × 4.5 − 13(15) |
| nucleus | One | –//– | –//– |
| chloroplast | parietal, single, lobed | –//– | –//–, or with a smooth unlobed edges |
| pyrenoid (P) | 1–4 | –//– | 1-2(4) |
| P starch envelope | + | + | 3 and more, sometimes absent |
| 3. Reproduction: | |||
| vegetative | fragmentation of filaments | –//– | –//– |
| asexual | |||
| by zoospores (Z) | + | + | + |
| shape of Z | oval or nearly spherical # | –//– | fusiform, rod-shaped, oval, spherical |
| sizes of Z, µm | 10–20 of length # | 7 − 13 × 4 − 9 | 6 − 20 × 4.5 − 8 |
| number of flagella per Z | 4, 8, 12–24 | 2, 4 | 4, 8 and more |
| by aplanospores (A) | + | + | + |
| shape of A | spherical, cylindrical, occasionally constricted | spherical | spherical, cylindrical, ellipsoid, occasionally constricted |
| sizes of A, µm | 14–15 in diameter, or 10 − 21 × 8 − 16 | 8–11 in diameter | 12 − 17 × 6 − 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, I.N.; Kulakova, N.V.; Boldina, O.N.; Tupikova, G.S. Barranca variabilis sp. nov.—A New Terrestrial Alga of the Genus Barranca (Chaetophorales, Chlorophyta) from the Baikal Region (Russia). Diversity 2023, 15, 583. https://doi.org/10.3390/d15040583
Egorova IN, Kulakova NV, Boldina ON, Tupikova GS. Barranca variabilis sp. nov.—A New Terrestrial Alga of the Genus Barranca (Chaetophorales, Chlorophyta) from the Baikal Region (Russia). Diversity. 2023; 15(4):583. https://doi.org/10.3390/d15040583
Chicago/Turabian StyleEgorova, Irina N., Nina V. Kulakova, Olga N. Boldina, and Galina S. Tupikova. 2023. "Barranca variabilis sp. nov.—A New Terrestrial Alga of the Genus Barranca (Chaetophorales, Chlorophyta) from the Baikal Region (Russia)" Diversity 15, no. 4: 583. https://doi.org/10.3390/d15040583