
Where the Immatures of Triassic Diptera Developed

Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow 117647, Russia
Diversity 2023, 15(4), 582; https://doi.org/10.3390/d15040582
Submission received: 1 April 2023
/
Revised: 14 April 2023
/
Accepted: 19 April 2023
/
Published: 21 April 2023
(This article belongs to the Special Issue Paleoecology of Insects)
Abstract
:Immature Diptera are more diverse and abundant in fresh water than any other insect order. The question arises whether the earliest dipterans, known from the Upper Buntsandstein of Europe (early Anisian, Middle Triassic), already developed in water and whether such mode of life was inherited from the ancestor of the order or if, alternatively, the dipteran larvae were ancestrally terrestrial. Nematoceran immatures have become common and diverse in the Mesozoic fossil record since the Middle Jurassic, but the vast majority of them represent only two culicomorphan families, Chaoboridae and Chironomidae. Earlier records and records of immatures of other families from the Mesozoic are extremely rare. A total of four larvae and about 30 pupae have been described from Anisian assemblages of France and Spain. Among these, one larva clearly belongs to Culicomorpha, the infraorder most closely associated with fresh waters, and one larva to Bibionomorpha, the infraorder most closely associated with terrestrial habitats, while the rest are hard to classify. Nevertheless, most of the pupae are morphologically similar to the semi-aquatic pupae of extant Limoniidae and Ptychopteridae from wet habitats. The oligopneustic respiratory systems of the Anisian larvae and their apparently allochthonous burial also suggest their semi-aquatic development, which appears to have been the ancestral mode of life of Diptera. The absence of dipteran immatures in lacustrine Konservat-Lagerstätten of the Madygen and the Cow Branch Formations (Ladinian–Carnian and Norian, respectively) is explained by the aquatic conditions being unfavorable for insect immatures and invertebrates in general.
1. Introduction
Diptera is the most aquatic of all insect orders, even though the majority of dipteran species develop in terrestrial habitats. If all the free-living dipterans that require a wet environment during the larval stage are classified as aquatic, then “forty-one (26%) of the 158 families of Diptera have aquatic representatives. These 41 families are scattered throughout the dipteran phylogeny and include nearly 46,000 aquatic species worldwide…, representing about 30% of all formally described species of Diptera. Thus, Diptera have more aquatic representatives than any other order of insects—at least three times more than the Coleoptera and Trichoptera” [1]. Among the nematocerans, to which this review will be limited, the share of aquatic taxa is even higher. More than half of the nematoceran families have such members; these include all 17 families of Psychodomorpha, Culicomorpha, and Tipulomorpha, with the exception of Trichoceridae (the infraorder concept generally follows the classification of Hennig [2,3]). Additionally, selected species of some bibionomorphan families (e.g., Bibionidae, Scatopsidae, Cecidomyiidae, Sciaridae) are known to be water dependent [4,5].
Immatures of only a few nematoceran families use dissolved oxygen for respiration and are strictly aquatic. For example, the larvae and pupae of Blephariceridae and Simuliidae dwell in streams without any contact with the atmosphere. The larvae of all the six extant genera of Chaoboridae are also strictly aquatic, but, in some of these genera, they are capable of aquatic respiration and never rise to the water surface to breathe, while, in others, the larvae rise to the surface to take in oxygen directly from the atmosphere. Moreover, the chaoborid Mochlonyx velutinus (Ruthe, 1831) alternates the two modes of larval respiration between its two annual generations [6].
Much more often, the larvae of large families, such as Limoniidae, Psychodidae, Chironomidae, and Ceratopogonidae, occur in a wide variety of habitats, ranging from entirely terrestrial to strictly aquatic ones, including lakes and rivers [5,7]. The larvae of only a few nematoceran taxa can live at depths exceeding their own body length, while the majority is restricted to edges of pools and streams and other habitats very near water level, where they can reach the atmospheric air by spiracles at the abdominal apex [8]. For example, the larvae of the psychodid subfamily Psychodinae are strictly aquatic in some species, but, in the vast majority of species, they develop in diverse semi-aquatic habitats on exposed muddy shorelines “where conditions fluctuate between aquatic and terrestrial according to seasonal and climatic variation… Some burrow in marginal accumulations of mud and silt; others creep between layers of fallen leaves…, in films of water over half submerged leaves or stones…, among fluviatile mosses and filamentous algae on rocks…, in water filled tree-holes and other phytotelmata” [9] (p. 754).
2. Hypothetical Ancestral Lifestyle of Dipteran Larvae
The question arises whether the dipteran larvae have been ancestrally terrestrial or aquatic and whether paleontological data can solve this long-standing issue. Edwards [10] considered feeding on decaying organic matters, such as rotten wood or rich humus, as ancestral for dipteran larvae, whereas the lineages with aquatic larvae have derived from the terrestrial ones. Hinton [11] argued that the development of spiracles during the ontogenesis clearly indicated that the larvae of the first dipterans had been submerged, but a reduction of legs could rather occur in the larvae living not in water but in a liquid or semi-liquid nutritious substrate. “The ancestral dipterous larva could hardly have been aquatic, since among recent insects there is not a single instar of larva losing its legs after it has become aquatic; and the complete absence of legs in the Order is one of its most conspicuous features. I believe that the prototype of the Diptera had a larva which lived in liquid or semi-liquid decaying vegetable matter, possibly fungi” [11] (p. 469).
Kovalev [12] suggested that the first dipteran larvae dwelt under bark of submerged rotting trees, such as the larvae of extant Eutanyderus Alexander, 1928 from Australia [13]. He came to this idea by observing a resemblance between nematoceran wings from the Triassic locality Dzhailoucho (Madygen Formation, Kyrgyzstan) and the wings of Tanyderidae (the family not found at this locality). My own experience of collecting tanyderid larvae and pupae in Chile and Australia [14,15] suggested that such a boring mode of life is indeed widespread among extant Tanyderidae of the Southern Hemisphere. Both larvae and pupae stay inside submerged rotting logs until the adult moult; thus, as a rule, neither larval nor pupal exuviae move into the water.
For a long time, Triassic deposits yielded only adult dipterans, usually as isolated wings. At first sight, the hypothetical ancestral lifestyle, suggested by Kovalev, would well explain not only the loss of the larval legs, useless under the bark, but also the rarity of the oldest dipteran immatures in the fossil record. However, new Triassic findings allow us to take a fresh look at this problem.
3. Larval Respiratory System and Mode of Life
The respiratory system seems to be the key to life in water. Seven types of tracheal systems are recognized in Diptera after Keilin [16]. Three of these are polypneustic, with numerous functional spiracles: holopneustic, with 10 pairs (on the prothorax, metathorax, and each of eight abdominal segments); peripneustic, with 9 pairs; and hemipneustic, with 8 pairs. The remaining four types are oligopneustic: amphipneustic, with two pairs of spiracles, anterior and posterior; propneustic and metapneustic, with only one pair, anterior or posterior, respectively; and apneustic, without functional spiracles. Keilin came to the conclusion that all holometabolous larvae have 10 pairs of spiracles during all stages, but some spiracle pairs usually remain non-functional. Holopneustic larvae occur rarely, for example, in the dipteran families Bibionidae, Cramptonomyiidae, and Pachyneuridae [11,17,18]. Polypneustic respiratory systems are ancestral for holometabolous larvae. They are typical of Mecoptera [11], including the only extant mecopteran family with aquatic larvae, Nannochoristidae (hemipneustic) [19]. Among dipteran larvae, polypneustic systems are known only in the larvae of Bibionomorpha, which are terrestrial. This can be viewed as an argument in favor of the terrestrial development of the first Diptera.
The larvae of Bibionomorpha are predominantly terrestrial and, therefore, are not usually included in manuals on freshwater Diptera [20,21]. However, the larvae of some bibionomorphans develop in phytotelmata, i.e., small water-filled cavities on terrestrial plants such as tree holes and leaf bases of Bromeliaceae, and other water environments. Even some holopneustic bibionid larvae occur in semi-aquatic conditions. For example, Pentethria funebris Meigen, 1804 has been collected from wet soil along the side of a brook [5,22]. Peripneustic and hemipneustic respiratory systems of various bibionomorph larvae do not seem to preclude aquatic or semi-aquatic life. The larvae of some Cecidomyiidae and Sciaridae occur in water-filled bamboo stumps [23], and the larvae of some Scatopsidae are aquatic [24]. The larvae of the scatopsid Holoplagia richardsi (Edwards, 1934) occur in water-filled tree holes together with typical dendrolimnobic psychodids, Telmatoscopus advena (Eaton, 1893). Other scatopsid larvae have been found to be madicolous, developing in small springs and brooklets inside a very thin, not exceeding 1–2 mm, film of flowing water together with the larvae of Ephemeroptera, Trichoptera, and Psychodidae. “Compared with those of terrestrial Scatopsidae, the aquatic larvae apparently do not display any peculiar adaptation to aquatic life. The respiratory system is apparently peripneustic in the last instar as in all other known genera of the family and there is no unusual regression either of the tracheal system or of the spiracles” [24] (p. 49).
The indication “in the last instar” is very important. During the course of larval development, the number of functional spiracles often increases [11]. Among Diptera, this has been described in detail, for example, in Tanyderidae: the larvae of Tanyderus pictus Philippi, 1865 progressively change from the apneustic 1st instar through the metapneustic 2nd instar to the amphipneustic 3rd and 4th instars (Figures 27–30 and 33–41) in [14].
Polypneustic dipteran larvae also pass during their development through oligopneustic stages [16]. Usually the youngest stage is metapneustic, with only the posterior spiracles functional, as in the first instar of Bibionidae [25]. The second-instar larvae of Bibio Geoffroy, 1764 and Dilophus Meigen, 1803 also lack a complete set of functional spiracles [26]. The developmental sequence metapneustic–amphipneustic–peripneustic has been recorded in Scatopsidae [11] and can be presumed for other bibionomorphan families developing in aquatic and semi-aquatic habitats. Hinton concluded [11] (p. 466) that “a metapneustic first instar is an adaptation to life in fluids, and its common occurrence in the Diptera is further evidence that the archetype of the Diptera had a larva that lived in fluids”.
The last-instar larvae of Tipulomorpha, Psychodomorpha, and Culicomorpha are oligopneustic (Table 1). Such respiratory systems occur in both aquatic and terrestrial larvae.
In a broad general way, the degree to which the functional spiracles are reduced indicates the degree to which the larva is adapted to life in water [11]. Thus, it is not surprising that only the 4th instar is hemipneustic in the mecopteran family Nannochoristidae, whereas the first three aquatic instars are apneustic, with all of their spiracles non-functional [19]. In other Mecoptera and in numerous other insect orders, terrestrial larvae are polypneustic in all instars since the first one [11,27,28].
4. Larval Respiratory System in Mesozoic Members of Extant Families
It would be worth comparing the tracheal systems of extinct and extant members of the extant families; however, Mesozoic larvae are so insufficiently known that no comparison can yet be made. Nematoceran immatures have become common and diverse in the Mesozoic fossil record since the Middle Jurassic, but the vast majority of them represent only two culicomorphan families, Chaoboridae and Chironomidae [29,30] both known from the Triassic [31,32]. Mesozoic records of larvae of other extant families are extremely rare and, to my knowledge, only a few such larvae have been described (Table 1). These represent three oligopneustic respiratory types:
In all these cases, the tracheal systems are similar to those of the recent members of each family. Additionally, two large larvae of a supposed trichocerid “Trichonomites” aquaticus Kalugina, 1986 have been described, without any mention of the tracheal system, from the Lower Cretaceous of Mongolia [36]. Later, the species was synonymized under Paleotrichocera mongolica Kalugina, 1986, the only trichocerid species at the locality, based on its compact head capsule and the secondary partition of abdominal segments typical of Trichoceridae [37]. The authors agreed that the proportions and length of the larvae suggested an aquatic habitat. “Recent terrestrial larvae are of more stout, compact shape. If our interpretation were right, then this species would be exceptional among Trichoceridae in having aquatic larvae” [37] (p. 72).
Re-examination of the holotype in the course of the present study revealed the presence of a distinct pair of large posterior spiracles and, possibly, a pair of smaller, anterior spiracles (Figure 1). Thus, the larva appeared to be amphipneustic, as in the recent Trichoceridae. However, its familial affinity is doubtful because the pattern of abdominal pseudo-segmentation is rather similar in the larvae of three families, Trichoceridae, Psychodidae, and Anisopodidae. In extant Trichoceridae, larval abdominal segments are each secondary subdivided by folds into two or three subequal parts. In contrast, in the larva under discussion, one incomplete ring in the anterior one-third of each of the seven abdominal segments looks like a narrow tergal plate or a dorsal creeping welt (Figure 1), similar to those of the psychodid genus Trichomyia Haliday in Curtis, 1839, which is a specialized wood-borer (Figure 59) in [17]. Moreover, the Cretaceous larva is also micropilose, thin, more than 10× as long as wide, without terminal setae, and less sclerotized, similar to Trichomyia [38]. A Cretaceous adult Trichomyia has been described from Burmese amber [39] but has not been confirmed yet. However, the larvae of Trichomyia are distinctly smaller; moreover, no members of Psychodidae or Psychodomorpha are known from the Lower Cretaceous of Western Mongolia [36]. Thus, wood-boring can be hypothesized, but the larvae cannot be classified with confidence, and the synonymy proposed by Krzemińska et al. [37] is not accepted herein.
Lacustrine deposits are the main source of paleoentomological material [40]. Therefore, the abundance of fossil immatures of Chaoboridae and Chironomidae appearing in the record since the Middle Jurassic clearly indicates that at least these culicomorphans dwelt in aquatic, usually lacustrine, habitats. The rarity of other nematoceran immatures suggests their occasional, allochthonous burial. Apparently, they developed far from the taphonomic window, for example, in lotic habitats (streams and seeps), phytotelmata, or riparian wetlands (edges of streams and marshes), where the extant members of Simuliidae, Tanyderidae, Psychodidae, and Ptychopteridae occur [21].
5. On the Absence of Dipteran Immatures in Some Triassic Lacustrine Konservat-Lagerstätten
Molecular data have placed the origin of the crown group and main infraorders of Diptera at approximately 265–267 Ma [41]. Such an estimated Permian origin is not supported by fossil record. No Diptera have yet been found in the Permian, despite the fact that numerous Permian localities have yielded a great number of diverse insect fossils. The latter included some 10,000 impressions from Russia, housed at the Paleontological Institute in Moscow. The oldest dipterans are known since the early Middle Triassic, i.e., the Upper Buntsandstein (early Anisian) of Spain, Germany, and France [42,43,44,45,46].
The fossil record of Diptera in the Triassic is poor: only a little over a dozen sites from all continents except Antarctica have yielded dipterans [47]. Most such sites have yielded but a few dipteran wings. Diverse and relatively numerous dipterans have been collected only at three localities: the Grès à Voltzia in France (early Anisian), the Madygen Formation in Kyrgyzstan (Ladinian–Carnian), and the Cow Branch Formation in the USA (Norian) [43,48,49]. Sediments at the two latter localities appear to have been formed in lakes with harsh water conditions, which adversely affected aquatic organisms.
In lacustrine sediments of the Cow Branch Formation, no gill-bearing nymphs of the amphibiotic orders Odonata, Plecoptera, Trichoptera, and Ephemeroptera have been found (and no adult mayflies), which strongly suggests poor water quality [50]. Indeed, sedimentological and geochemical analyses reconstructed the ancient “Lake Solite” as saline, alkaline, and probably shallow. It could have been similar to modern dolomite lakes of South Australia, where the water is toxic to gill-bearing nymphs, aquatic larvae, and other sensitive groups [51]. Therefore, the diversity of adults combined with the lack of immatures of Diptera at the Cow Branch Formation [49] suggested that the dipterans developed some place nearby but not in the lake.
The diversity of adults and the lack of immatures of Diptera in lacustrine sediments of Dzhailoucho, the northern area of the Madygen Formation [48], can be explained in a similar way. Amphibiotic Odonata, Plecoptera, and Trichoptera account for only 1% of more than 20,000 fossil insects collected there and about 10% of some 500 described species, being represented exclusively by adults. No adult Ephemeroptera and only two fragments of mayfly nymphs have been reported among specimens collected there by expeditions of the Paleontological Institute of the Soviet (now Russian) Academy of Sciences, Moscow, in 1957–1971 [52]; one nymph was listed later [53]. In fact, one of those nymphs does not belong to mayflies [54], and another has been erroneously reported from Dzhailoucho, while it actually came from Lower Jurassic deposits overlying the Madygen Formation in the southeastern area of Madygen (Sinitshenkova, pers. comm., 2023). No nymphs have been found at Dzhailoucho during more recent (2006–2009) expeditions either (Bashkuev, pers. comm., 2023). The complete absence of immatures of amphibiotic insects suggests that water characteristics of the ancient lake at Dzailoucho have been unfavorable for them.
One recent find may be interpreted as contradicting the above hypothesis. An isolated wing from Dzhailoucho has been described as Triassomyia Lukashevich, 2022, the first record of Culicomorpha at the locality and the oldest known representative of Chaoboridae, one of those few families with extant species developing exclusively in water and never in wet ground or other semi-aquatic habitats [32]. Since the Middle Jurassic, chaoborids flourished in Jurassic and Early Cretaceous fresh waters of Asia; their immatures and/or adults often quantitatively dominate in taphocenoses [29,30]. In contrast, no immatures and only two chaoborid wings have been described from the Lower Jurassic [55,56] and a single wing from the Triassic [32]. The absence of larval imprints from a collection usually leaves a chance that the larvae have been overlooked in the field since they are often small, delicate, and transparent in a fossilized state. However, the absence of immatures of Chaoboridae cannot be explained by such a collection bias. The head capsules with mandibles of the chaoborid larvae and their anal fans are easily spotted in the field by an expert collector (such as all PIN paleoentomologists who worked in Dzhailoucho). Moreover, the lion’s share of chaoborid imprints usually represent the pupae—dark, rather rigid, and, unlike the larvae, not easily damaged during transportation. Chaoborid pupae often dominate in insect taphocenoses, whereas no chaoborid immatures have been found at Dzhailoucho in the field or, subsequently, in the lab. Most probably, Triassomyia larvae developed in small and/or temporary water bodies near the large lake at Dzhailoucho, which helped them avoid pressure of abundant and diverse fish predators, as has been observed in some extant chaoborids [57]. Such ecology may account for the extreme rarity of Triassic chaoborids in lacustrine deposits.
In a comprehensive paper by Voigt and coauthors, the second trophic level of the food pyramid at the Madygen Formation is represented by molluscs, crustaceans, aquatic insects, and insect larvae, the latter being illustrated as a chaoborid (Figure 15) in [58]. In fact, only ostracods are known from Dzhailoucho in large numbers; only a single specimen of conchostracans and kazacharthrans each, and no insect immatures, bivalves, or gastropods have been recorded there [58].
As for the Coleoptera, which constitute more than half of the insects collected at Dzhailoucho, different opinions have been expressed about their habitats. It is usually assumed that most beetles from the Madygen Formation had been unspecialized aquatic or semi-aquatic predators and detritivores occurring on both sides of the shoreline, although no forms adapted for swimming have been found among them [53,59]. Most of these beetles have been classified as Schizophoridae and Triaplidae; however, neither of these families display specific adaptations typical of aquatic beetles. For example, the “schiza”—an elytro-thoracic interlocking device—can scarcely be a reliable indicator of aquatic or semi-aquatic modes of life; moreover, it has not been recorded in most Schizophoridae from Dzhailoucho [60,61].
6. Anisian Diptera
The oldest known fossil Diptera come from European Anisian deposits of similar age. The fossiliferous beds of the Estellencs Formation of Mallorca, Spain are dated as Aegean, which are slightly older than the Röt Formation in Germany and the Grès à Voltzia of the Vosges (hereinafter referred to as the Vosges) in France, both dated as Bithynian [44,62,63,64,65]. The number of collected dipterans varies between the localities dramatically: a single wing was found in the Röt Formation [45], a single larva and two pupae at the Pedra Alta site (upper part of the Estellencs Formation) [46], and about 70 adults, 30 pupae, and 3 larvae in the Vosges [66].
The assemblage of Anisian insects of the Vosges is diverse, and, among the nematocerans, three of the four infraorders sensu Hennig [2,3], Tipulomorpha, Psychodomorpha, and Bibionomorpha have been described based on adults [42,43]. No adult Culicomorpha, the infraorder most closely associated with fresh waters, were found.
Among these three Anisian outcrops, only in the Rőt Formation, the fossil fauna is not distinctly aquatic. The only autochthonous element in that assemblage, which may have been associated with water, are quite numerous Dasyleptus Brongniart, 1885 (Monura), whose lifestyle was probably semi-aquatic [67]. There is no evidence of other aquatic insects, such as mayfly nymphs, in the Rőt Formation [44]. In contrast, in both the French Vosges and the Spanish Pedra Alta site, diverse aquatic communities occurred without doubts. For outcrops of the Vosges a deltaic area is reconstructed, an environment transitional from nearshore to terrestrial, with unstable and occasionally harsh conditions, with numerous shallow drying pools, thus “it is evident that organisms must have suffered great variations in temperature, oxygen and salinity as time passed” [68]. Nevertheless, these unstable pools were suitable for numerous insect immatures, including nymphs of eight families of Ephemeroptera [69]. Sinitshenkova [70] characterized the Anisian insect assemblages of France and Spain as assemblages of one type: “taxonomically diverse assemblages dominated by various mayflies (including the burrowing ones), with considerable diversity of other aquatic insects and absence of aquatic bugs, recorded from deltaic sediments and probably representing the fauna of slow-flowing lowland channels and small ponds with soft bottom”. This conclusion was recently confirmed based on new data [65]. Such rich aquatic assemblages must have preserved dipteran larvae if they occurred in or near water during that time.
Indeed, Grauvogel [71] noted dipteran pupae among his finds in the first paper on the fauna of the Grès à Voltzia of the Vosges; later, numerous dipteran larvae were reported by Marchal-Papier [72]. As a result, the share of Diptera in the total number of collected insect specimens of the Vosges was estimated as 5.3%, which was unusually high for Triassic assemblages, with more than half of these being larvae [72], which is a unique case in the Triassic. Our revision of the Vosges collection demonstrated that numerous pupae (about 30, mostly exuviae) were diverse and, undoubtedly, belonged to Diptera [66]. This contradicts the hypothesis suggested by Kovalev [12] discussed above that the earliest dipteran larvae developed and pupated inside submerged tree trunks. At the same time, only three dipteran larvae have been confirmed, whereas many supposed ones turned out to represent other orders, mostly Ephemeroptera; thus, the previous estimate of the share of Diptera was at least twice the real value [66].
Two of three larvae from the Vosges are nematoceran without a doubt, but the third larva (gen. sp. 7) lacks the head capsule; thus, its attribution is unclear; all three described larvae are metapneustic (Figure 2). One larva, Anisinodus Lukashevich et al., 2010, turned out to be a member of Chironomoidea of an uncertain family. Therefore, this most aquatic infraorder is now known from as early as the Anisian [66]. It is worth to mention that only madicolous midges Thaumaleidae are amphipneustic at the last larval stage among extant Chironomoidea, whereas Simuliidae, Ceratopogonidae, and the vast majority of Chironomidae are apneustic (Table 1). Only several extant genera with metapneustic larvae have been recorded among Chironomidae: the posterior spiracles on the abdominal segment VIII are functional in madicolous larvae of some closely related genera of Podonominae, Afrochlus Freeman, 1964, Archaeochlus Brundin, 1966, and Austrochlus Cranston, 2002. Their larvae develop in small temporary streams shortly after seasonal rains flood the vegetative soil on rocks; they aggregate in shallow films of water or occupy moist interstitial spaces in the moss, keeping their posterior spiracles at the water surface [73,74,75]. “Metapneustic spiracles are probably beneficial in these hygropetric or moss habitats, where dissolved oxygen may be scarce” [75] (p. 326).
Such modes of life during the larval stage are currently widespread among Diptera: immatures of over 40 recent dipteran families develop in semi-aquatic habitats. In most of these families, the larvae depend on the atmospheric air and cannot survive prolonged submersion in water; thus, they are usually restricted to very shallow water or wet shores [66]. As already mentioned above, most species of the large families, such as Limoniidae, have metapneustic larvae, which rise to the water surface to breathe atmospheric air (Table 1). Fully aquatic larvae are less common; they can occur in the benthic zone of fast-flowing rivers and streams, being apneustic and living entirely submerged [7].
During the early Anisian, Central Europe and Iberia (including Mallorca) have been areas of the palaeoequatorial Pangaea with similar, semiarid climates [65]. A deltaic area with slow-flowing lowland channels in wet warm climate was reconstructed for the Vosges [62,68] and meandering rivers and floodplain for the Estellencs Formation [65]. Both deltaic environments, apparently, provided ample microhabitats for semi-aquatic development, such as wet or saturated mats or cushions of mosses and liverworts, fallen leaves, and organic-rich mud along edges of streams, lakes, small oxbow ponds, and other water bodies. Yet, another possible niche could have been phytotelmata in diverse Anisian plants, common in the outcrops of European Buntsandstein [68], such as the herbaceous fern Neuropteridium Weiss, 1869, the cycadophyte Zamites Brongiart, 1828, the shrubby conifer Pelourdea Seward, 1917 and diverse horsetails. For example, the broad clasping leaf bases of Pelourdea [76] and numerous broken stems of horsetails collected water and could be colonized by diverse dipteran larvae such as water-filled funnels of recent Bromeliaceae and bamboo stumps, respectively [4,23]. The larvae developing in such microhabitats could have become buried and fossilized only by chance, possibly when flood waters uprooted herbaceous or shallow-rooted plants and transferred them downstream.
Anisian Diptera are, evidently, too diverse to represent the common ancestor of the order, but some of them could have preserved the ancestral mode of life. The morphologies of, at least, some of the numerous nematoceran pupae (not associated with any of the three larvae) from the Vosges do not contradict semi-aquatic scenarios. The most abundant pupae, Voltziapupa Lukashevich et al., 2010, have well-sclerotized bodies, relatively well-developed armatures of transverse rows of spines or isolated large spines on the abdominal segments, and long unbranched respiratory horns with apical widening. Although the Vosges pupae cannot be classified even at the infraorder level, their mode of life can be hypothesized based on their morphological similarity with pupae of the extant Ptychopteridae and Limoniidae, which occur in wet microhabitats near water margins with soft substrate (fine-particulate detritus, mud) and breathe atmospheric air through their long, apically widened respiratory horns [66]. Similar pupae of Ptychopteridae and Limoniidae have been described from the Jurassic and Cretaceous deposits of Eurasia [77,78,79]. For example, the pupae of Limoniidae (sometimes several pupae on one slab) with well-sclerotized bodies, transverse rows of large tubercles on each abdominal segment, and long respiratory horns with apical widening have been described from the Upper Jurassic locality Shar Teg (Mongolia) (Figure 12) in [78].
Most of the modern classifications and phylogenetic systems of Diptera follow Hennig [3] in recognizing the five major groups, Tipulomorpha, Psychodomorpha, Culicomorpha, Bibionomorpha, and Brachycera. However, recent studies disagreed in various details concerning the composition and relationships of the nematoceran infraorders [80]. Sometimes authors considered the two highly specialized small families Deuterophlebiidae and Nymphomyiidae as the most basal among the extant fly lineages [80], but their unique morphologies are difficult to interpret phylogenetically. The separation of Tipulomorpha has been considered the first divergence in dipteran history already by Hennig [2,3], and many recent phylogenetic analyses (including those that examined both extant and extinct taxa) also recovered Tipulomorpha as a sister group to the remaining Diptera [48,49,81]. From the traditional point of view, which I share [82], the most abundant pupae from the Vosges, Voltziapupa, combining the characters of both Tipulomorpha and Psychodomorpha, must be close to the pupae of ancestral dipterans. Therefore, they could have preserved the ancestral mode of life.
Among the limited material collected at the Pedra Alta site, no dipteran adults, a single dipteran larva, and two pupae have been found. At least one pupa has been assigned to Voltziapupa cf. cornuta Lukashevich et al., 2010, described from the Vosges [46]. It is not surprising—the plants and insects of the Estellencs Formation show close affinities with those of the Upper Bundsandstein of central Europe. For example, the assemblages of Ephemeroptera are similar between all three Anisian outcrops, including three species from Mallorca, which have originally been described from the Vosges [65,69]. Additionally, collections from the French and the Spanish Buntsandstein share common certain larvae with, apparently, swimming hind legs, referred to as the “Sialidae-type” larvae, which remain undescribed [83].
A perfectly preserved single larva, Protoanisolarva Peñalver et al., 2022, with an amphipneustic respiratory system, has been described as a member of Anisopodoidea. Morphological similarities between Protoanisolarva and the larvae of extant Anisopodidae suggested a similar biology, i.e., terrestrial habitats, rich in decaying organic matter, along pool margins, with probable feeding on fungal mycelia and/or fungal fruiting bodies [46]. Any conclusions about widespread mycetophagy as early as the Triassic and about larval development in semi-liquid fungal fruiting bodies as the ancestral for Diptera are premature because only a single fungal fossil, hyphae of Palaeofibulus Osborn et al., 1989 (Basidiomycota), has thus far been described, only from Antarctica (Fremouw Formation, Early Triassic) [84,85]. Most larvae of the recent Anisopodidae are, indeed, terrestrial and have amphipneustic tracheal systems [46]. However, the larvae of a common and widespread extant genus Sylvicola Harris, 1776 have been found in a great variety of situations, where fermenting organic matter was available. These included phytotelmata, such as water-filled tree holes, leaf bases of Bromeliaceae [86], and fallen leaves of the New Zealand nikau palm, Rhopalostylis sapida Wendl et Drude, 1878, where the larvae of Sylvicola co-occured with the larvae of Psychodidae, Limoniidae, and Culicidae [87].
The extant family Anisopodidae has been known since the Early Jurassic (Sinemurian) [88], while members of anisopodid stem group, family Protorhyphidae, now extinct, had been widely distributed in the Triassic assemblages of Eurasia and North America, with the oldest occurrence in the Vosges [43]. Peñalver and coauthors reasonably pointed out that Protoanisolarva may be a representative of Protorhyphidae [46]; however, undoubted protorhyphid larvae remain unknown. Semi-aquatic development cannot be ruled out for the Triassic Protorhyphidae; however, in any case, these nematocerans must have occurred near water, as indicated by multiple records of their adults in Triassic outcrops.
It is logical to conclude that the larvae and pupae of Diptera dwelt in wet environments from the beginning. I agree with Hinton that the ancestor of the Diptera had larva that lived in liquid or semi-liquid decaying plant matter. At least some of the Anisian immatures of Diptera most probably preserved this ancestral mode of life and occurred near edges of small temporary streams, oxbow lakes, and marshes, in accumulations of mud and silt, or among filamentous algae and fluviatile mosses. The metapneustic tracheal system, found in three of the four Anisian larvae, is now the most common during the first larval stage because the common ancestor of Diptera lived in liquid or semi-liquid medium with occasional contact with the air. This mode of life has also been preserved in numerous recent nematoceran families. It has been described for the anisopodid larvae by Edwards and Keilin [86] (p. 9) as follows: “The early stages…are passed in situations where there is plenty of wet decaying and fermenting matter… These situations may be merely sodden or may be actually under water; the larvae are able to swim actively as well as crawl, and so long as they have occasional access to the air the amount of water present is immaterial”. Development in deeper water, without access to the atmospheric air, has evolved secondarily multiple times in different families with apneustic larvae, sometimes as a rule and sometimes as an exception within the family. The exceptional rarity of dipteran larvae and pupae in Triassic lacustrine deposits indirectly confirms their allochthonous origin due to dwelling in wet but not lacustrine environments.
Funding
This research was funded by the Russian Science Foundation grant number 21–14–00284.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No shareable research data were generated in this research.
Acknowledgments
The author is deeply indebted to Roman Rakitov (Borissiak Paleontological Institute RAS, Moscow) for fruitful discussion, linguistic advice, and assistance in taking scanning electron images. I am also grateful to A. Prokin (Papanin Institute for Biology of Inland Waters, RAS, Borok), N. Sinitshenkova, D. Shcherbakov, and A. Bashkuev for information about Dzhailoucho insect fauna and E. Karasev for discussion of Anisian plants (all at Borissiak Paleontological Institute RAS, Moscow). I thank the three anonymous reviewers for their insightful comments on the initial version of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Adler, P.H.; Courtney, G.W. Ecological and societal services of aquatic Diptera. Insects 2019, 10, 70. [Google Scholar] [CrossRef]
- Hennig, W. Kritische Bemerkungen über den Bauder Flügelwurzel bei den Dipteren und die Frage nach der Monophylie der Nematocera. Stuttg. Beitr. Naturk. 1968, 193, 1–23. [Google Scholar]
- Hennig, W. Insect Phylogeny; John Wiley & Sons: Hoboken, NJ, USA, 1981; pp. 1–514. [Google Scholar]
- Greeney, H.F. The insects of plant-held waters: A review and bibliography. J. Trop. Ecol. 2001, 17, 241–260. [Google Scholar] [CrossRef]
- Wagner, R.; Barták, M.; Borkent, A.; Courtney, G.; Godddeeris, B.; Haenni, J.-P.; Knutson, L.; Pont, A.; Rotheray, G.E.; Rozkosny, R.; et al. Global diversity of dipteran families (Insecta Diptera) in freshwater (excluding Simulidae, Culicidae, Chironomidae, Tipulidae and Tabanidae). Hydrobiologia 2008, 595, 489–519. [Google Scholar] [CrossRef]
- Monchadsky, A.S. On seasonal dimorphism of larvae of Mochlonyx culiciformis De Geer (Diptera, Culicidae). Trudy ZIN AN SSSR 1953, 13, 363–372. (In Russian) [Google Scholar]
- de Jong, H.; Oosterbroek, P.; Gelhaus, J.; Reusch, H.; Young, C. Global diversity of craneflies (Insecta, Diptera: Tipulidea or Tipulidae sensu lato) in freshwater. Hydrobiologia 2008, 595, 457–467. [Google Scholar] [CrossRef]
- Drake, M. The importance of temporary waters for Diptera (true-flies). AquaDoc 2001, 17, 26–39. [Google Scholar]
- Duckhouse, D.A.; Duckhouse, S.R. Insecta: Diptera, Psychodidae. In Freshwater Invertebrates of the Malaysian Region; Yule, C.M., Sen, Y.H., Eds.; Academy of Sciences Malaysia: Kuala Lumpur, Malaysia, 2004; pp. 750–762. [Google Scholar]
- Edwards, F.W. The phylogeny of nematocerous Diptera: A critical review of some recent suggestions. Verh. III Int. Entomol.-Kongr. 1926, 2, 111–130. [Google Scholar]
- Hinton, H.E. On the reduction of functional spiracles in the aquatic larvae of the Holometabola, with notes on the moulting process of spiracles. Trans. R. Entomol. Soc. Lond. 1947, 98, 449–473. [Google Scholar] [CrossRef]
- Kovalev, V.G. New data on the initial stages of evolution of Diptera. In Diptera (Insecta), Their Systematics, Geographic Distribution and Ecology; Academy of Sciences USSR: Leningrad, Former Soviet Union, 1983; pp. 60–66. (In Russian) [Google Scholar]
- Colless, D.H.; McAlpine, D.K. Diptera. In The Insects of Australia; Melbourne University Press: Carlton, Australia, 1970; pp. 656–740. [Google Scholar]
- Lukashevich, E.D.; Shcherbakov, D.E. First description of Tanyderidae (Diptera) larvae from South America. Russ. Entomol. J. 2014, 23, 121–138. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Shcherbakov, D.E. On morphology of Tanyderus pictus (Diptera: Tanyderidae) pupa and adult from Chile. Russ. Entomol. J. 2016, 25, 79–95. [Google Scholar] [CrossRef]
- Keilin, D. Respiratory systems and respiratory adaptations in larvae and pupae of Diptera. Parasitology 1944, 36, 1–66. [Google Scholar] [CrossRef]
- Krivosheina, N.P.; Mamaev, B.M. A Key to Wood-Inhabiting Larvae of Diptera; Nauka: Moscow, Russia, 1967; pp. 1–367. (In Russian) [Google Scholar]
- Krivosheina, N.P.; Mamaev, B.M. Cramptonomyiidae (Diptera, Nematocera), a dipteran family new for the fauna of the USSR: Its morphology, ecology, and phylogenetic relationships. Entomol. Rev. 1970, 49, 886–898. [Google Scholar]
- Pilgrim, R.L.C. The aquatic larva and the pupa of Choristella philpotti Tillyard, 1917 (Mecoptera: Nannochoristidae). Pac. Insects 1972, 14, 151–168. [Google Scholar]
- Narchuk, E.P. Diptera. Generic part. In Key to Freshwater Invertebrates of Russia and Adjacent Lands; Tsalolikhin, S.J., Ed.; ZIN RAS: St. Petersburg, Russia, 1999; Volume 4, pp. 8–32. [Google Scholar]
- Bouchard, R.W. Guide to Aquatic Invertebrates of the Upper Midwest; Water Resources Center, University of Minnesota: St. Paul, MN, USA, 2004; pp. 1–215. [Google Scholar]
- Salmela, J.; Autio, O.; Ilmonen, J. A survey on the nematoceran (Diptera) communities of southern Finnish wetlands. Memo. Soc. Fauna Flora Fenn. 2007, 83, 33–47. [Google Scholar]
- Sota, T.; Mogi, M. Species richness and altitudinal variation in the aquatic metazoan community in bamboo phytotelmata from North Sulawesi. Res. Popul. Ecol. 1996, 38, 275–281. [Google Scholar] [CrossRef]
- Haenni, J.P.; Vaillant, F. Description of dendrolimnobiontic larvae of Scatopsidae (Diptera) with a review of our knowledge of the preimaginal stages of the family. Mitt. Schweiz. Entomol. Ges. 1994, 67, 43–59. [Google Scholar]
- Krivosheina, N.P. European larvae Bibionidae (Diptera, Nematocera), with key to some species. Pedobiologia 1962, 1, 210–227. (In Russian) [Google Scholar]
- Skartveit, J. The larvae of European Bibioninae (Diptera, Bibionidae). J. Nat. Hist. 2002, 36, 449–485. [Google Scholar] [CrossRef]
- Tan, J.; Hua, B. Morphology of immature stages of Bittacus choui (Mecoptera: Bittacidae) with notes on its biology. J. Nat. Hist. 2008, 42, 2127–2142. [Google Scholar] [CrossRef]
- Jiang, L.; Yue, C.; Hua, B. Larval morphology of Panorpoidea kuandianensis (Insecta, Mecoptera, Panorpodidae) and its evolutionary implication. Zookeys 2014, 398, 69–82. [Google Scholar]
- Kalugina, N.S.; Kovalev, V.G. Jurassic Diptera of Siberia; Nauka: Moscow, Russia, 1985; pp. 1–198. (In Russian) [Google Scholar]
- Lukashevich, E.D. Larvae—A key to evolution of Culicoidea (Diptera) in the Mesozoic. Alavesia 2008, 2, 59–72. [Google Scholar]
- Krzemiński, W.; Jarzembowski, E. Aenne triassica sp. n., the oldest representative of the family Chironomidae (Insecta: Diptera). Polish J. Entomol. 1999, 68, 445–449. [Google Scholar]
- Lukashevich, E.D. The oldest occurrence of Chaoboridae (Insecta: Diptera). Russ. Entomol. J. 2022, 31, 417–421. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Krzemiński, W. New Jurassic Tanyderidae (Diptera) from Asia with first find of larvae. Zoosymposia 2009, 3, 155–172. [Google Scholar] [CrossRef]
- Kalugina, N.S. New psychodomorph dipterans from the Mesozoic of Siberia (Diptera: Eoptychopteridae, Ptychopteridae). Paleontol. Zh. 1989, 1, 65–77. [Google Scholar]
- Jell, P.A.; Duncan, P.M. Invertebrates, mainly insects, from the freshwater, Lower Cretaceous, Koonwarra Fossil Bed (Korumburra Group), South Gippsland, Victoria. Mem. Ass. Austral. Palaeontol. 1986, 3, 111–205. [Google Scholar]
- Kalugina, N.S. True flies. Muscida (=Diptera). Infraorders Tipulomorpha and Culicomorpha. Insects in the Early Cretaceous ecosystems of the West Mongolia. Moscow: Nauka. Trans. Joint Sov.-Mongol. Palaeontol. Exped. 1986, 28, 112–125. (In Russian) [Google Scholar]
- Krzemińska, E.; Krzemiński, W.; Dahl, C. Monograph of Fossil Trichoceridae (Diptera): Over 180 Million Years of Evolution; Institute of Systematics and Evolution of Animals PAS: Krakow, Poland, 2009; pp. 1–172. [Google Scholar]
- Kvifte, G.M.; Wagner, R. Psychodidae (sand flies, moth flies or owl flies). In Manual of Afrotropical Diptera. Volume 2. Nematocerous Diptera and Lower Brachycera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; pp. 607–632. [Google Scholar]
- Cockerell, T.D.A. Fossil insects. Ann. Entomol. Soc. Am. 1917, 10, 1–22. [Google Scholar] [CrossRef]
- Zherikhin, V.V. Pattern of insect burial and conservation. In History of Insects; Rasnitsyn, A.P., Quicke, D.L.J., Eds.; Kluver Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2002; pp. 17–64. [Google Scholar]
- Bertone, M.A.; Courtney, G.W.; Wiegmann, B.M. Phylogenetics and temporal diversification of the earliest true flies (Insecta: Diptera) based on multiple nuclear genes. Syst. Entomol. 2008, 33, 668–687. [Google Scholar] [CrossRef]
- Krzemiński, W.; Krzemińska, E.; Papier, F. Grauvogelia arzvilleriana sp. n.—The oldest Diptera species (Lower/Middle Triassic of France). Acta Zool. Cracov. 1994, 37, 95–99. [Google Scholar]
- Krzemiński, W.; Krzemińska, E. Triassic Diptera: Descriptions, revisions and phylogenetic relations. Acta Zool. Cracov. 2003, 46, 153–184. [Google Scholar]
- Bashkuev, A.; Sell, J.; Aristov, D.; Ponomarenko, A.; Sinitshenkova, N.; Mahler, H. Insects from the Buntsandstein of Lower Franconia and Thuringia. Paläontol. Z. 2012, 86, 175–185. [Google Scholar] [CrossRef]
- Lukashevich, E.D. The oldest Diptera (Insecta) from the Upper Buntsandstein (early Middle Triassic) of Europe. Zootaxa 2021, 5067, 135–143. [Google Scholar] [CrossRef]
- Peñalver, E.; Matamales-Andreu, R.; Nel, A.; Pérez-de la Fuente, R. Early adaptation of true flies (Diptera) to moist and continental environments. Pap. Palaeontol. 2022, 8, e1472. [Google Scholar] [CrossRef]
- Lara, M.B.; Lukashevich, E.D. The first Triassic dipteran (Insecta) from South America, with review of Hennigmatidae. Zootaxa 2013, 3710, 81–92. [Google Scholar] [CrossRef]
- Shcherbakov, D.E.; Lukashevich, E.D.; Blagoderov, V.A. Triassic Diptera and initial radiation of the order. Int. J. Dipterol. Res. 1995, 6, 75–115. [Google Scholar]
- Blagoderov, V.; Grimaldi, D.A.; Fraser, N.C. How time flies for flies: Diverse Diptera from the Triassic of Virginia and early radiation of the order. Am. Mus. Novit. 2007, 3572, 1–39. [Google Scholar] [CrossRef]
- Criscione, J.; Grimaldi, D. The oldest predaceous water bugs (Insecta, Heteroptera, Belostomatidae), with implications for paleolimnology of the Triassic Cow Branch Formation. J. Paleontol. 2017, 91, 1166–1177. [Google Scholar] [CrossRef]
- Liutkus, C.M.; Beard, J.S.; Fraser, N.C.; Ragland, P.C. Use of fine-scale stratigraphy and chemostratigraphy to evaluate conditions of deposition and preservation of a Triassic Lagerstatte, south-central Virginia. J. Paleolimnol. 2010, 44, 645–666. [Google Scholar] [CrossRef]
- Sinitshenkova, N.D. A review of Triassic mayflies, with a description of new species from Western Siberia and Ukraine (Ephemerida=Ephemeroptera). Paleontol. J. 2000, 34, S275–S283. [Google Scholar]
- Shcherbakov, D.E. Madygen, Triassic Lagerstätte number one, before and after Sharov. Alavesia 2008, 2, 113–124. [Google Scholar]
- Sukacheva, I.D.; Sinitshenkova, N.D. A review of the Triassic caddisflies with a description of new species from the Middle–Upper Triassic of Kyrgyzstan. Paleontol. Zhurnal 2023, 1, 42–48. [Google Scholar]
- Ansorge, J. Insekten aus dem oberen Lias von Grimmen (Vorpommern, Norddeutschland). Neue Paläont. Abhandl. 1996, 2, 1–132. [Google Scholar]
- Lukashevich, E.D. Mesozoic Dixidae (Insecta: Diptera) and systematic position of Dixamima Rohdendorf, 1964 and Rhaetomyia Rohdendorf, 1962. Paleontol. Zhurnal 1996, 1, 48–53. [Google Scholar]
- Monchadsky, A.S. To the knowledge of mosquito larvae (Diptera, Culicidae). I. Larva Cryophila lapponica Mart. Parasitol. Sb. ZIN AN SSSR 1939, 7, 142–169. (In Russian) [Google Scholar]
- Voigt, S.; Buchwitz, M.; Fischer, J.; Kogan, I.; Moisan, P.; Schneider, J.W.; Spindler, F.; Brosig, A.; Preusse, M.; Scholze, F.; et al. Triassic life in an inland lake basin of the warm-temperate biome—The Madygen Lagerstätte (Southwest Kyrgyzstan, Central Asia). In Terrestrial Conservation Lagerstätten. Windows into the Evolution of Life on Land; Fraser, N.C., Sues, H.-D., Eds.; Dunedin: Edinburgh, UK, 2017; pp. 65–104. [Google Scholar]
- Ponomarenko, A.G. Composition and ecological characteristics of Mesozoic Coleoptera. In Mesozoic Coleoptera. Tr. Paleontol. Ins. AN SSSR 1977, 161, 1–16. (In Russian) [Google Scholar]
- Ponomarenko, A.G.; Prokin, A.A. Review of the paleontological data on the evolution of aquatic beetles (Coleoptera). Paleontol. J. 2015, 49, 1383–1412. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Prokin, A.A. The position of the Paleozoic genus Tunguskagyrus Yan, Beutel et Lawrence in the family Triaplidae sensu n. (Coleoptera, Archostemata: Schizophoroidea). Entomol. Rev. 2018, 98, 872–882. [Google Scholar] [CrossRef]
- Gall, J.C. Faunes et paysages du Grès à Voltzia du Nord des Vosges. Essai paléoécologique sur le Buntsandstein supérieur. Mém. Serv. Carte Géol. Als. Lorr. 1971, 3, 1–318. [Google Scholar]
- Selden, P.; Nudds, J. Evolution of Fossil Ecosystems; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Brauckmann, C.; Schlüter, T. Neue Insekten aus der Trias von Unter-Franken. Geol. Palaeontol. 1993, 27, 181–199. [Google Scholar]
- Matamales-Andreu, R.; Peñalver, E.; Mujal, E.; Oms, O.; Scholze, F.; Juárez, J.; Galobart, A.; Fortuny, J. Early–Middle Triassic fluvial ecosystems of Mallorca (Balearic Islands): Biotic communities and environmental evolution in the equatorial western peri-Tethys. Earth Sci. Rev. 2021, 222, 103783. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Przhiboro, A.A.; Marchal-Papier, F.; Grauvogel-Stamm, L. The oldest occurrence of immature Diptera (Insecta), Middle Triassic, France. Ann. Soc. Entomol. Fr. 2010, 46, 4–22. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P. Taxonomy and morphology of Dasyleptus Brongniart, 1885, with description of a new species (Insecta: Machilida: Dasyleptidae). Russ. Entomol. J. 1999, 8, 145–154. [Google Scholar]
- Gall, J.-C.; Grauvogel-Stamm, L. The early middle Triassic ‘Grès à Voltzia’ formation of Eastern France: A model of environmental refugium. C. R. Palevol. 2005, 4, 637–652. [Google Scholar] [CrossRef]
- Sinitshenkova, N.D.; Marchal-Papier, F.; Grauvogel-Stamm, L.; Gall, J.-C. The Ephemeridea (Insecta) from the Grès à Voltzia (early Middle Triassic) of the Vosges (NE France). Paläontol. Z. 2005, 79, 377–397. [Google Scholar] [CrossRef]
- Sinitshenkova, N.D. Ecological history of the aquatic insects. In History of Insects; Rasnitsyn, A.P., Quicke, D.L.J., Eds.; Kluver Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2002; pp. 388–417. [Google Scholar]
- Grauvogel, L. Contribution a l’étude du Grès à Voltzia. Bull. Soc. Géol. Fr. 1947, 5, 35–37. [Google Scholar]
- Marchal-Papier, F. Les Insectes du Buntsandstein des Vosges (NE de la France). Biodiversité et Contributions aux Modalités de la Crise Biologique du Permo-Trias. Ph.D. Dissertation, Université Louis Pasteur, Strasbourg, France, 1998. [Google Scholar]
- Brundin, L. Transantarctic relationships and their significance, as evidenced by chironomid midges with a monograph of the subfamilies Podonominae and Aphroteniinae and the Austral Heptagyiae. Kungl. Sven. Vetensk. Handle Fjärde S. 1966, 11, 1–472. [Google Scholar]
- Cranston, P.S.; Edward, D. Afrochlus Freeman: An African gondwanan midge and the phylogeny of the Podonominae (Diptera: Chironomidae). Syst. Entomol. 1998, 23, 77–90. [Google Scholar] [CrossRef]
- Cranston, P.S.; Edward, D.H.D.; Colless, D.H. Archaeochlus Brundin: A midge out of time (Diptera: Chironomidae). Syst. Entomol. 1987, 12, 313–334. [Google Scholar] [CrossRef]
- Ash, S.R. Growth habit and systematics of the Upper Triassic plant Pelourdea poleoensis, Southwestern USA. Rev.Palaeobot. Palynol. 1987, 5, 37–49. [Google Scholar] [CrossRef]
- Lukashevich, E.D. First pupae of the Eoptychopteridae and Ptychopteridae from the Mesozoic of Siberia (Insecta: Diptera). Paleontol. J. 1995, 29, 164–170. [Google Scholar]
- Lukashevich, E.D. Limoniidae (Diptera) in the Upper Jurassic of Shar Teg, Mongolia. Zoosymposia 2009, 3, 131–154. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Arillo, A. New Eoptychoptera (Insecta: Diptera, Ptychopteridae) from the Lower Cretaceous of Spain. Cret. Res. 2016, 58, 254–264. [Google Scholar] [CrossRef]
- Wiegmann, B.M.; Yeates, D.K. Phylogeny of Diptera. In Manual of Afrotropical Diptera. Volume 1. Introductory Chapters and Keys to Diptera Families; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; pp. 253–265. [Google Scholar]
- Wood, D.M.; Borkent, A. Phylogeny and classification of the Nematocera. In Manual of Nearctic Diptera; McAlpine, J.F., Wood, D.M., Eds.; Biosystematics Research Centre: Ottawa, ON, Canada, 1989; Volume 3, pp. 1333–1370. [Google Scholar]
- Lukashevich, E.D.; Ribeiro, G.C. Mesozoic fossils and phylogeny of Tipulomorpha (Insecta: Diptera). J. Syst. Palaeontol. 2019, 17, 635–652. [Google Scholar] [CrossRef]
- Prokin, A.A.; Bashkuev, A.S. Trialarva coburgensis gen et sp. nov., a remarkable fossil holometabolan larva (Insecta: Coleoptera) from the Triassic of Germany. PalZ 2021, 95, 55–60. [Google Scholar] [CrossRef]
- Osborn, J.M.; Taylor, T.N.; White, J.A. Palaeofibulus gen. nov., a clamp-bearing fungus from the Triassic of Antarctica. Mycologia 1989, 81, 622–626. [Google Scholar] [CrossRef]
- Halbwachs, H.; Harper, C.J.; Krings, M. Fossil Ascomycota and Basidiomycota, with notes on fossil lichens and Nematophytes. Encycl. Mycol. 2021, 1, 378–395. [Google Scholar]
- Edwards, F.W.; Keilin, D. Diptera. Fam. Protorhyphidae, Anisopodidae, Pachyneuridae, Trichoceridae. Genera Insectorum 1928, 190, 1–41. [Google Scholar]
- Derraik, J.G.; Heath, A.C. Immature Diptera (excluding Culicidae) inhabiting phytotelmata in the Auckland and Wellington regions. N. Z. J. Mar. Freshw. Res. 2005, 39, 981–987. [Google Scholar] [CrossRef]
- Rohdendorf, B.B. Order Diptera. In Osnovy Paleontologii. Chlenistonogie: Trakheinye, Khelitserovye (Fundamentals of Paleontology: Arthropoda: Tracheata, Chelicerata), Akad; Nauk SSSR: Moscow, Russia, 1962; pp. 307–345. (In Russian) [Google Scholar]
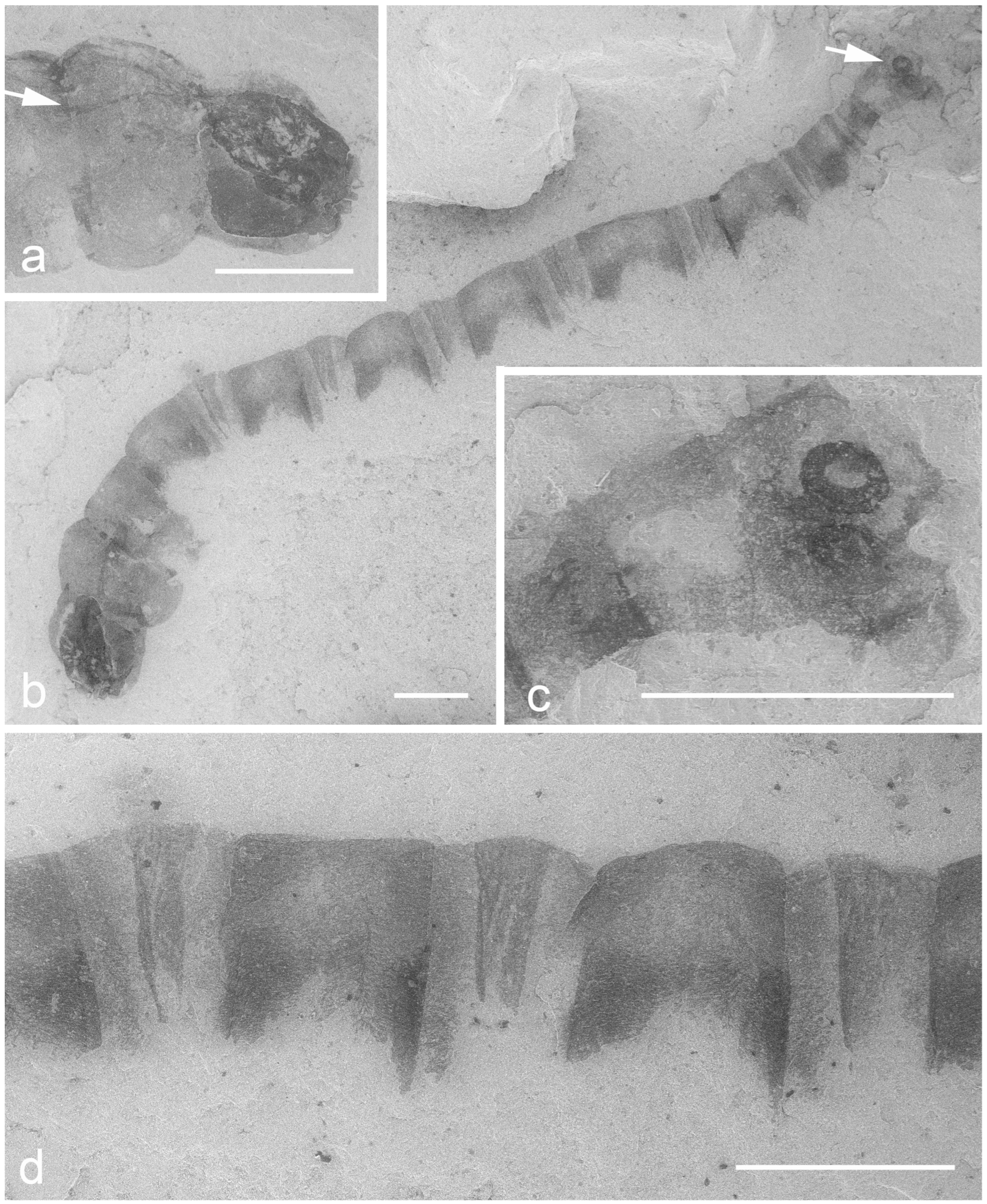
Figure 1.
Larva of “Trichonomites” aquaticus Kalugina, 1986, holotype, from Gurvan (Early Cretaceous); SEM photo, uncoated specimen, LVSTD detector. (a) head and thorax on the counterpart. (b) total view of the part. (c) apex of the abdomen on the part. (d) first abdominal segments on the part. Scale bars, 1 mm. Arrows mark the anterior and posterior spiracles.
Figure 1.
Larva of “Trichonomites” aquaticus Kalugina, 1986, holotype, from Gurvan (Early Cretaceous); SEM photo, uncoated specimen, LVSTD detector. (a) head and thorax on the counterpart. (b) total view of the part. (c) apex of the abdomen on the part. (d) first abdominal segments on the part. Scale bars, 1 mm. Arrows mark the anterior and posterior spiracles.

Figure 2.
Metapneustic dipteran larvae from the Vosges (early Anisian). (a) total view of the holotype of Anisinodus crinitus Lukashevich et al., 2010. (b) gen. sp.7; no. 9073, apex of the abdomen. (c,d) gen. sp. 8; no. 9347, apex of the abdomen on the counterpart and total view of the part. The arrows mark the posterior spiracles.
Figure 2.
Metapneustic dipteran larvae from the Vosges (early Anisian). (a) total view of the holotype of Anisinodus crinitus Lukashevich et al., 2010. (b) gen. sp.7; no. 9073, apex of the abdomen. (c,d) gen. sp. 8; no. 9347, apex of the abdomen on the counterpart and total view of the part. The arrows mark the posterior spiracles.


{kind=link}
{kind=link}
Table 1.
Respiratory systems in the last larval stage and finds of Mesozoic immatures in aquatic families of Tipulomorpha, Psychodomorpha, and Culicomorpha sensu Hennig.
Table 1.
Respiratory systems in the last larval stage and finds of Mesozoic immatures in aquatic families of Tipulomorpha, Psychodomorpha, and Culicomorpha sensu Hennig.
| Family | Amphipneustic | Metapneustic | Apneustic | Other Types | Mesozoic Larvae | Mesozoic Pupae |
|---|---|---|---|---|---|---|
| Limoniidae | + | + | pp | |||
| Pediciidae | + | |||||
| Tipulidae | + | |||||
| Cylindrotomidae | + | |||||
| Ptychopteridae | + | ll | pp | |||
| Tanyderidae | + | + | l | |||
| Psychodidae | + | + | l | |||
| Blephariceridae | + | |||||
| Deuterophlebiidae | + | |||||
| Nymphomyiidae | + | |||||
| Dixidae | + | |||||
| Corethrellidae | + | |||||
| Chaoboridae | + | + | lll | ppp | ||
| Culicidae | + | |||||
| Chironomidae | + | + | lll | ppp | ||
| Ceratopogonidae | + | |||||
| Simuliidae | + | ll | p | |||
| Thaumaleidae | + |
l—larva; p—pupa; ll and pp—more than 5 specimens of larvae or pupae, respectively; and lll and ppp—more than 100 specimens of larvae or pupae, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lukashevich, E.D. Where the Immatures of Triassic Diptera Developed. Diversity 2023, 15, 582. https://doi.org/10.3390/d15040582
AMA Style
Lukashevich ED. Where the Immatures of Triassic Diptera Developed. Diversity. 2023; 15(4):582. https://doi.org/10.3390/d15040582
Chicago/Turabian StyleLukashevich, Elena D. 2023. "Where the Immatures of Triassic Diptera Developed" Diversity 15, no. 4: 582. https://doi.org/10.3390/d15040582
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.