
Nitrogen Allocation Tradeoffs Within-Leaf between Photosynthesis and High-Temperature Adaptation among Different Varieties of Pecan (Carya illinoinensis [Wangenh.] K. Koch)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
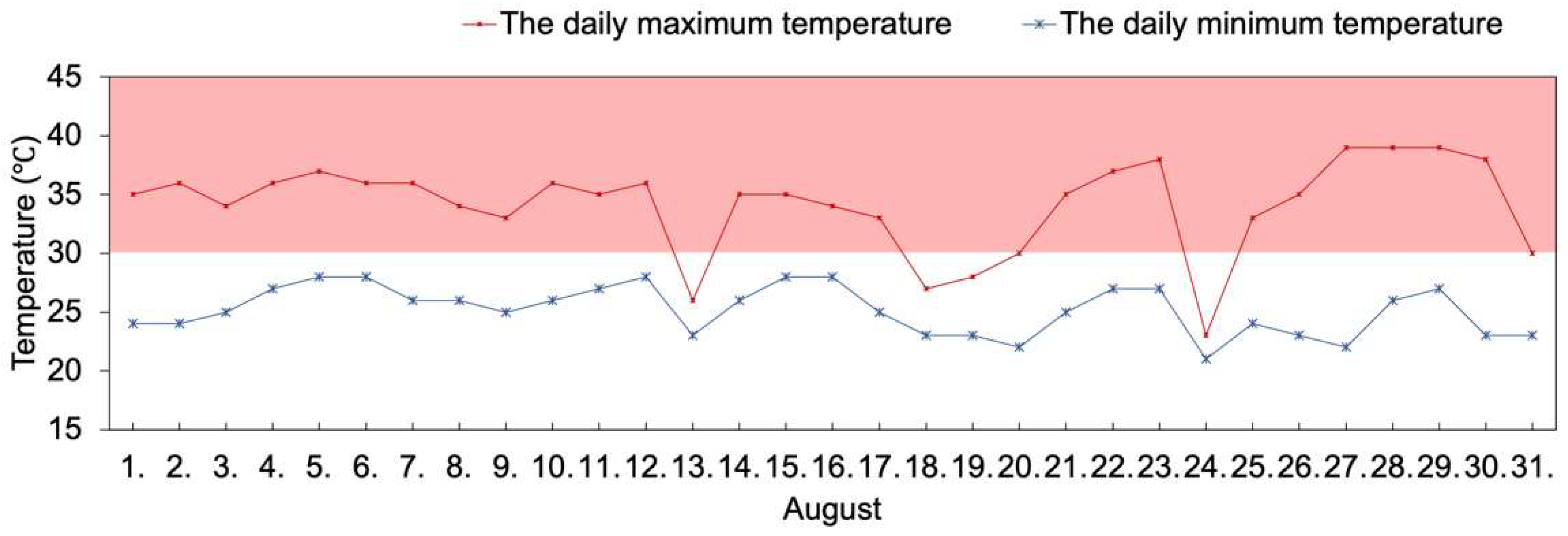
2.1. Daily Temperature Records
2.2. Differences in Leaf Nitrogen and Photosynthesis from Eight Varieties under High Temperatures
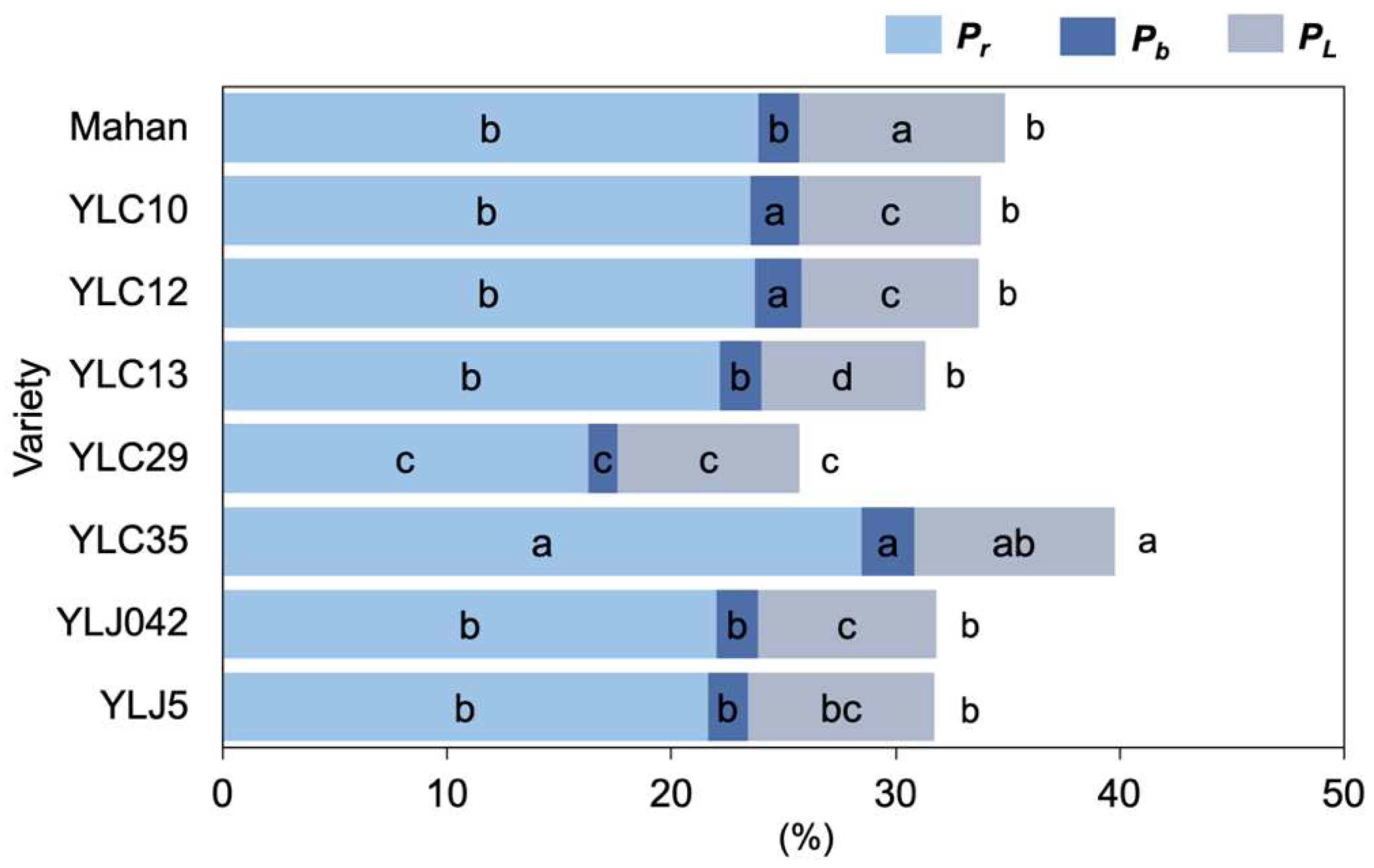
2.3. Changes in Leaf Nitrogen Allocation of Eight Varieties under High Temperatures
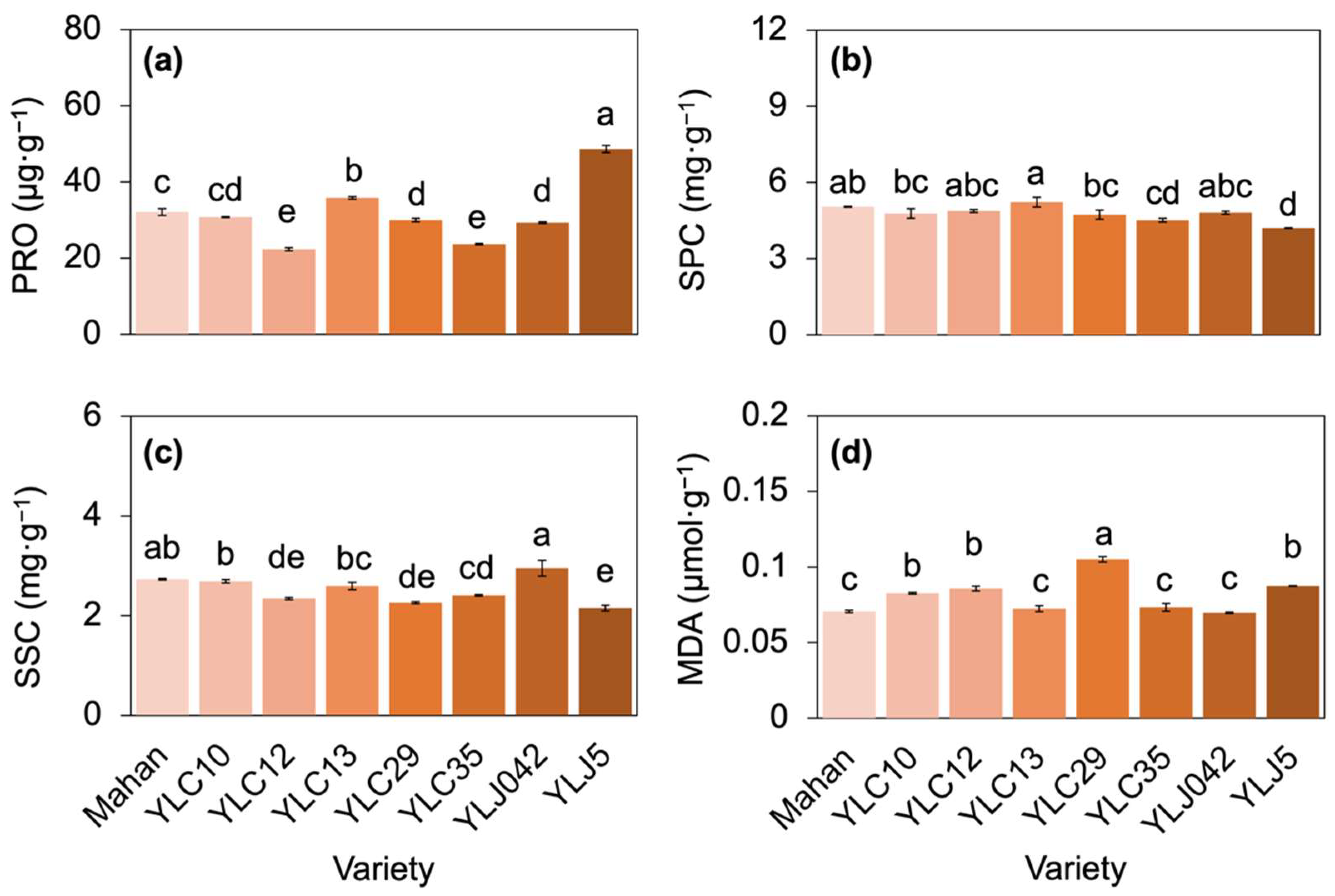
2.4. Responses of Osmolytes’ Accumulation and Lipid Peroxidation to High Temperatures
2.5. Principal Component Analysis
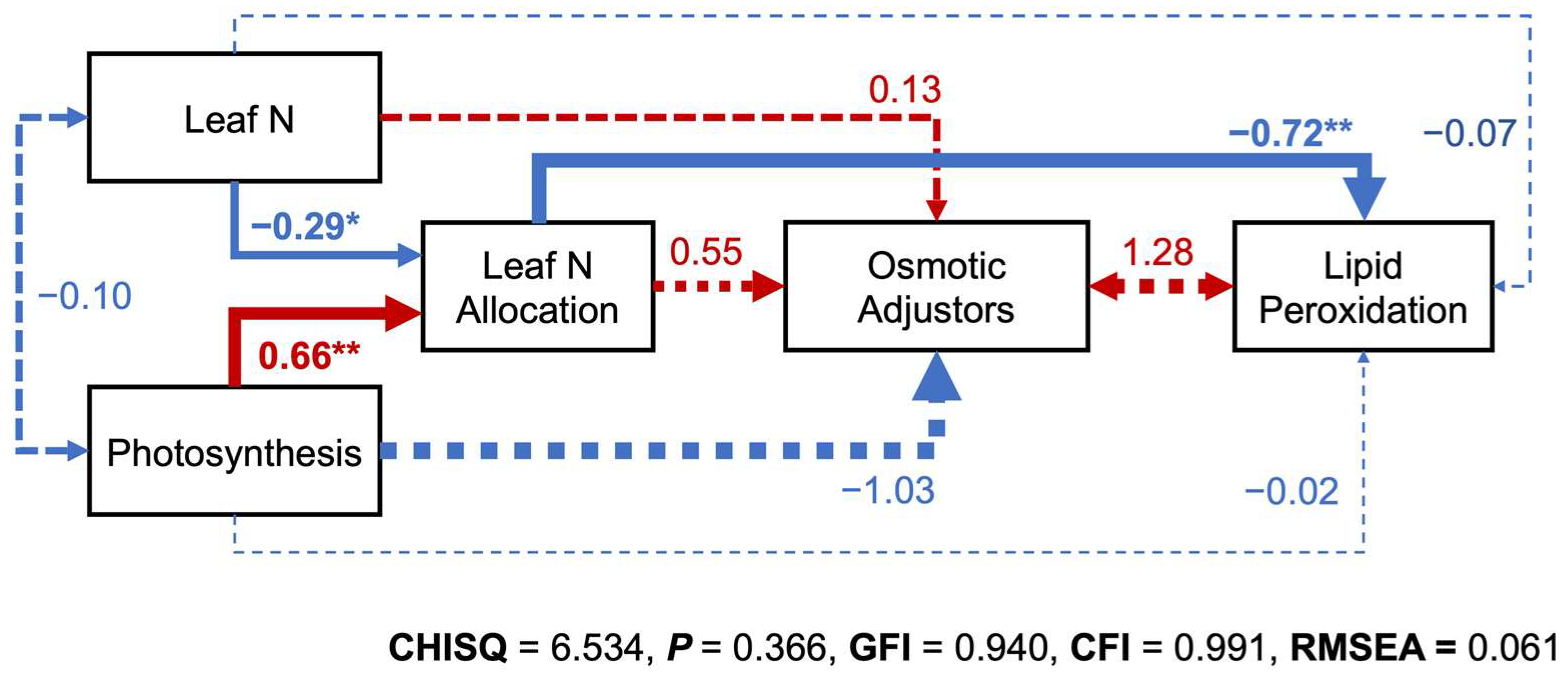
2.6. Interrelations
3. Discussion
3.1. Resource Conservative and Acquisitive Strategies
3.2. Leaf N Allocation Reflected the Growth-Defense Tradeoffs
3.3. Tolerance to High Temperatures May Rely on Compensation Mechanisms
3.4. Tradeoffs between Growth and Defense Varied with Varietal Characteristic
4. Materials and Methods
4.1. Study Area and Plant Material
4.2. Measurements of Leaf Gas Exchange
4.3. Determinations of Leaf Chlorophyll Content, Leaf Mass per Area, and Nitrogen Content
4.4. Calculations of Leaf Photosynthetic Nitrogen-Use Efficiency and Nitrogen Allocation
4.5. Assay of Osmolytes’ Accumulation and Lipid Peroxidation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhao, X.; Huang, L.J.; Sun, X.F.; Zhao, L.L.; Wang, P.C. Differential Physiological, Transcriptomic, and Metabolomic Responses of Paspalum wettsteinii Under High-Temperature Stress. Front. Plant Sci. 2022, 13, 865608. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; Leach, J.E. High Temperature-induced Plant Disease Susceptibility: More than the Sum of its Parts. Curr. Opin. Plant Biol. 2020, 56, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Kebert, M.; Vuksanovic, V.; Stefels, J.; Bojovic, M.; Horak, R.; Kostic, S.; Kovacevic, B.; Orlovic, S.; Neri, L.; Magli, M.; et al. Species-Level Differences in Osmoprotectants and Antioxidants Contribute to Stress Tolerance of Quercus robur L., and Q. cerris L. Seedlings under Water Deficit and High Temperatures. Plants 2022, 11, 1744. [Google Scholar] [CrossRef]
- Landman, W. Climate change 2007: The Physical Science Basis. S. Afr. Geogr. J. 2010, 92, 86–87. [Google Scholar] [CrossRef] [Green Version]
- Pachauri, R.; Meyer, L.; Working Group, I; Working Group, III. Climate Change: Synthesis Report; IPCC: Geneva, Switzerland, 2016. [Google Scholar]
- Lobell, D.B.; Field, C.B. Global Scale Climate—Crop Yield Relationships and the Impacts of Recent Warming. Environ. Res. Lett. 2007, 2, 014002. [Google Scholar] [CrossRef]
- Lobell, D.B.; Asner, G.P. Climate and Management Contributions to Recent Trends in US Agricultural Yields. Science 2003, 299, 1032. [Google Scholar] [CrossRef]
- Dreyer, I. Nutrient Cycling is an Important Mechanism for Homeostasis in Plant Cells. Plant Physiol. 2021, 187, 2246–2261. [Google Scholar] [CrossRef]
- Perdomo, J.A.; Carmo-Silva, E.; Hermida-Carrera, C.; Flexas, J.; Galmes, J. Acclimation of Biochemical and Diffusive Components of Photosynthesis in Rice, Wheat, and Maize to Heat and Water Deficit: Implications for Modeling Photosynthesis. Front. Plant Sci. 2016, 7, 01719. [Google Scholar] [CrossRef] [Green Version]
- Berry, J.; Bjorkman, O. Photosynthetic Response and Adaptation to Temperature in Higher—Plants. Annu. Rev. Plant Phys. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing Yield Gaps through Nutrient and Water Management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, X.C.; Zhao, Y.; Korpelainen, H.; Li, C.Y. Sex-specific Nitrogen Allocation Tradeoffs in the Leaves of Populus cathayana Cuttings under Salt and Drought Stress. Plant Physiol. Bioch. 2022, 172, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Photosynthesis and Nitrogen Relationships in Leaves of C3 Plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Siso, S.; Flexas, J.; Galmes, J.; Niinemets, U.; Sancho-Knapik, D.; Gil-Pelegrin, E. Coordinated Modifications in Mesophyll Conductance, Photosynthetic Potentials and Leaf Nitrogen Contribute to Explain the Large Variation in Foliage Net Assimilation Rates across Quercus ilex Provenances. Tree Physiol. 2017, 37, 1084–1094. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Clarke, V.C. The Nitrogen Cost of Photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef]
- Ridenour, W.M.; Vivanco, J.M.; Feng, Y.L.; Horiuchi, J.; Callaway, R.M. No Evidence for Trade-offs: Centaurea Plants from America are Better Competitors and Defenders. Ecol. Monogr. 2008, 78, 369–386. [Google Scholar] [CrossRef] [Green Version]
- Takashima, T.; Hikosaka, K.; Hirose, T. Photosynthesis or Persistence: Nitrogen Allocation in Leaves of Evergreen and Deciduous Quercus species. Plant Cell Environ. 2004, 27, 1047–1054. [Google Scholar] [CrossRef]
- Zhong, C.; Jian, S.F.; Huang, J.; Jin, Q.Y.; Cao, X.C. Trade-off of within Leaf Nitrogen Allocation between Photosynthetic Nitrogen-use Efficiency and Water Deficit Stress Acclimation in Rice (Oryza sativa L.). Plant Physiol. Biochem. 2019, 135, 41–50. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The Worldwide Leaf Economics Spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Le Roux, X.; Walcroft, A.S.; Daudet, F.A.; Sinoquet, H.; Chaves, M.M.; Rodrigues, A.; Osorio, L. Photosynthetic Light Acclimation in Peach Leaves: Importance of Changes in Mass: Area ratio, Nitrogen concentration, and Leaf Nitrogen Partitioning. Tree Physiol. 2001, 21, 377–386. [Google Scholar] [CrossRef]
- Byeon, S.; Song, W.; Park, M.; Kim, S.; Kim, S.; Lee, H.; Jeon, J.; Kim, K.; Lee, M.; Lim, H.; et al. Canopy Height Affects the Allocation of Photosynthetic Carbon and Nitrogen in Two Deciduous Tree Species under Elevated CO2. J. Plant Physiol. 2022, 268, 153584. [Google Scholar] [CrossRef] [PubMed]
- Osada, N.; Onoda, Y.; Hikosaka, K. Effects of Atmospheric CO2 Concentration, Irradiance, and Soil Nitrogen Availability on Leaf Photosynthetic Traits of Polygonum sachalinense around Natural CO2 Springs in Northern Japan. Oecologia 2010, 164, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, Z.; Sun, Z.; Wan, X. Analysis of Pecan Cultivars Mahan and Western in East China. Genet. Mol. Res. 2016, 15, gmr.150387. [Google Scholar] [CrossRef] [PubMed]
- Mekouar, M.A. Food and Agriculture Organization of the United Nations (FAO). Yearb. Int. Environ. Law 2015, 26, 617–631. [Google Scholar] [CrossRef]
- Yang, B.B.; Yao, X.H.; Zeng, Y.R.; Zhang, C.C. Genome-Wide Identification, Characterization, and Expression Profiling of AP2/ERF Superfamily Genes under Different Development and Abiotic Stress Conditions in Pecan (Carya illinoinensis). Int. J. Mol. Sci. 2022, 23, 2920. [Google Scholar] [CrossRef]
- Jiao, Y.; Zhang, J.H.; Pan, C.D. Genome-Wide Analysis of the GDSL Genes in Pecan (Carya illinoensis K. Koch): Phylogeny, Structure, Promoter Cis-Elements, Co-Expression Networks, and Response to Salt Stresses. Genes 2022, 13, 1103. [Google Scholar] [CrossRef]
- Palma-Cano, L.E.; Pinon-Castillo, H.A.; Tarango-Rivero, S.H.; Carbon, A.; Salas-Leiva, J.; Munoz-Castellanos, L.N.; Cravo-Laureau, C.; Duran, R.; Orrantia-Borunda, E. Effect of Organic and Conventional Farming on Soil Bacterial Diversity of Pecan Tree (Carya illinoensis K. Kosh) Orchard across Two Phenological Stages. Lett. Appl. Microbiol. 2021, 72, 556–569. [Google Scholar] [CrossRef]
- Momayyezi, M.; Rippner, D.A.; Duong, F.V.; Raja, P.V.; Brown, P.J.; Kluepfel, D.A.; Earles, J.M.; Forrestel, E.J.; Gilbert, M.E.; McElrone, A.J. Structural and Functional Leaf Diversity Lead to Variability in Photosynthetic Capacity across a Range of Juglans regia Genotypes. Plant Cell Environ. 2022, 45, 2351–2365. [Google Scholar] [CrossRef]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant Domestication through an Ecological Lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef]
- Vilela, A.E.; Gonzalez-Paleo, L. Changes in Resource-use Strategy and Phenotypic Plasticity associated with Selection for Yield in Wild Species Native to Arid Environments. J. Arid Environ. 2015, 113, 51–58. [Google Scholar] [CrossRef]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and Processes in Crop Domestication: An Historical Review and Quantitative Analysis of 203 Global Food Crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.Y.; Wang, H.; Wright, I.J.; Zhu, X.G.; Niinemets, L.; Li, Z.L.; Sun, D.S.; Dong, N.; Zhang, W.F.; Zhou, Z.L. Enhanced Photosynthetic Nitrogen Use Efficiency and Increased Nitrogen Allocation to Photosynthetic Machinery under Cotton Domestication. Photosynth. Res. 2021, 150, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.D.; Qian, R.J.; Zhang, Y.J.; Zhang, X.L.; Ma, X.H.; Zheng, J. Physiological and Gene Expression Changes of Clematis crassifolia and Clematis cadmia in Response to Heat Stress. Front. Plant Sci. 2021, 12, 624875. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.H.; Lam, H.M.; Zhang, J.H. Inhibition of Photosynthesis and Energy Dissipation Induced by Water and High Light Stresses in Rice. J. Exp. Bot. 2007, 58, 1207–1217. [Google Scholar] [CrossRef]
- Liu, T.; Ren, T.; White, P.J.; Cong, R.H.; Lu, J.W. Storage Nitrogen Co-ordinates Leaf Expansion and Photosynthetic Capacity in Winter Oilseed Rape. J. Exp. Bot. 2018, 69, 2995–3007. [Google Scholar] [CrossRef]
- Onoda, Y.; Wright, I.J.; Evans, J.R.; Hikosaka, K.; Kitajima, K.; Niinemets, U.; Poorter, H.; Tosens, T.; Westoby, M. Physiological and Structural Tradeoffs Underlying the Leaf Economics Spectrum. New Phytol. 2017, 214, 1447–1463. [Google Scholar] [CrossRef] [Green Version]
- Ripullone, F.; Grassi, G.; Lauteri, M.; Borghetti, M. Photosynthesis-nitrogen Relationships: Interpretation of Different Patterns between Pseudotsuga menziesii and Populus × uroamericana in a Mini-stand Experiment. Tree Physiol. 2003, 23, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Ma, Y.C.; Zheng, D.; Wisniewski, M.; Cheng, Z.M. Meta-Analysis of the Effect of Overexpression of Dehydration-Responsive Element Binding Family Genes on Temperature Stress Tolerance and Related Responses. Front. Plant Sci. 2018, 9, 713. [Google Scholar] [CrossRef]
- Jing, J.G.; Guo, S.Y.; Li, Y.F.; Li, W.H. The Alleviating Effect of Exogenous Polyamines on Heat stress Susceptibility of Different Heat Resistant Wheat (Triticum aestivum L.) Varieties. Sci. Rep. 2020, 10, 7467. [Google Scholar] [CrossRef]
- Feng, Y.L. Nitrogen Allocation and Partitioning in Invasive and Native Eupatorium Species. Physiol. Plant. 2008, 132, 350–358. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting Photosynthetic Carbon Dioxide Response Curves for C3 Leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Caemmerer, S.V.; Berry, J.A. A Biochemical-Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, U.; Tenhunen, J.D. A Model Separating Leaf Structural and Physiological Effects on Carbon Gain along Light Gradients for the Shade-tolerant Species Acer saccharum. Plant Cell Environ. 1997, 20, 845–866. [Google Scholar] [CrossRef]
- Hikosaka, K.; Terashima, I. A Model of the Acclimation of Photosynthesis in the Leaves of C3 Plants to Sun and Shade with Respect to Nitrogen Use. Plant Cell Environ. 1995, 18, 605–618. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A Colorimetric Method for the Determination of Sugars. Nature. 1951, 168, 167. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts.I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Schermelleh-Engel, K.; Moosbrugger, H.; Müller, H. Evaluating the Fit of Structural Equation Models: Tests of Significance and Descriptive Goodness-of-Fit Measures. Methods Psychol. Res. Online 2003, 8, 23–74. [Google Scholar]
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; He, H.; He, B.; Li, T.; Liu, Y.; Zhu, S.; Zhang, G. Nitrogen Allocation Tradeoffs Within-Leaf between Photosynthesis and High-Temperature Adaptation among Different Varieties of Pecan (Carya illinoinensis [Wangenh.] K. Koch). Plants 2022, 11, 2828. https://doi.org/10.3390/plants11212828
Xu Q, He H, He B, Li T, Liu Y, Zhu S, Zhang G. Nitrogen Allocation Tradeoffs Within-Leaf between Photosynthesis and High-Temperature Adaptation among Different Varieties of Pecan (Carya illinoinensis [Wangenh.] K. Koch). Plants. 2022; 11(21):2828. https://doi.org/10.3390/plants11212828
Chicago/Turabian StyleXu, Qiwen, Huichuan He, Binghui He, Tianyang Li, Yumin Liu, Shunyao Zhu, and Gaoning Zhang. 2022. "Nitrogen Allocation Tradeoffs Within-Leaf between Photosynthesis and High-Temperature Adaptation among Different Varieties of Pecan (Carya illinoinensis [Wangenh.] K. Koch)" Plants 11, no. 21: 2828. https://doi.org/10.3390/plants11212828