

Propagation Methods Decide Root Architecture of Chinese Fir: Evidence from Tissue Culturing, Rooted Cutting and Seed Germination

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design
2.3. Harvest and Data Collection
2.3.1. Measure of Root Morphological Index
2.3.2. Measurement of Fractal Dimension of Roots
2.3.3. Measurement of the Topological Index of Roots
2.4. Statistical Analyses
3. Results
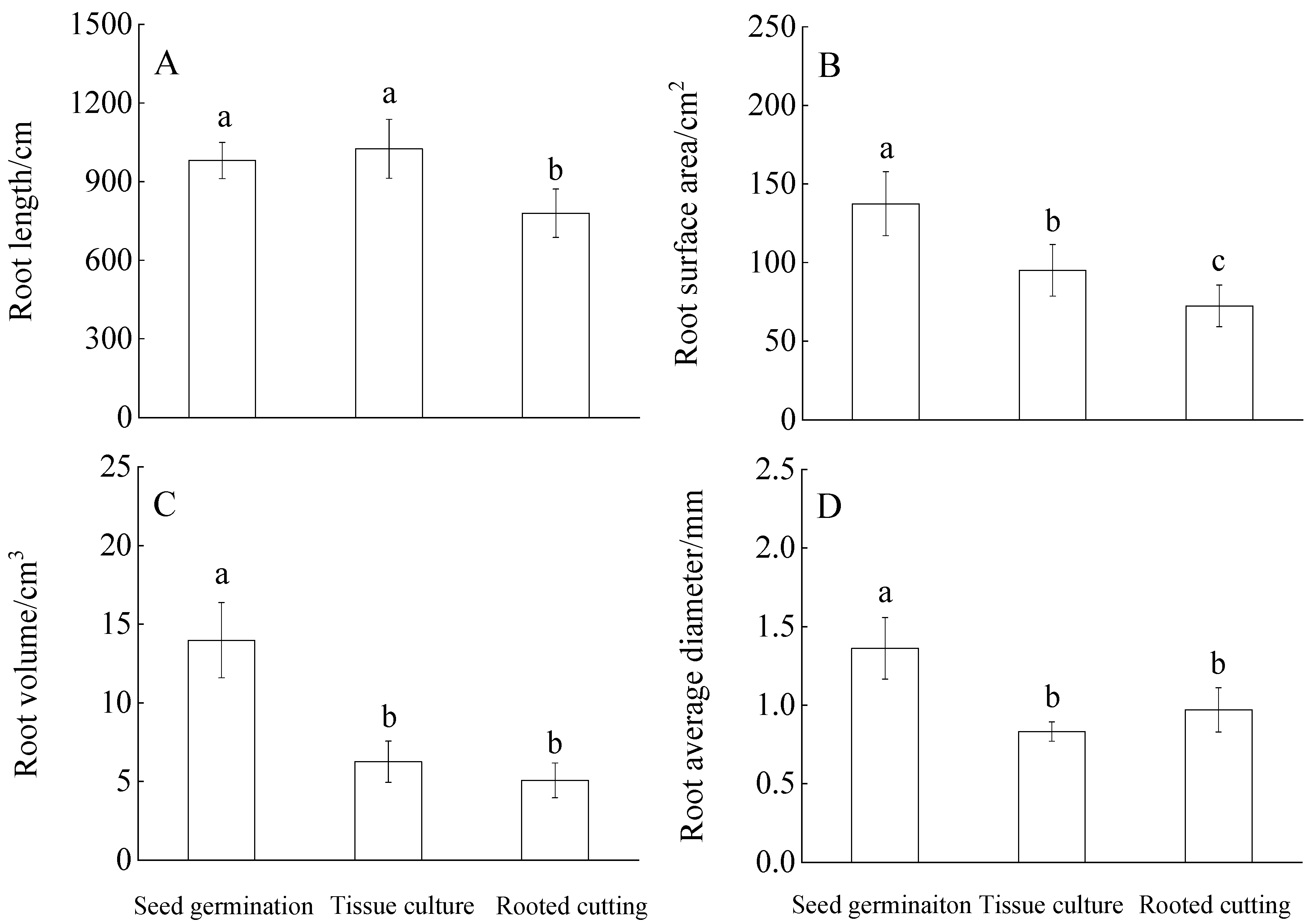
3.1. Root Plasticity under Different Propagation Methods
3.2. Root Geometric Architecture
3.3. Root Spatial Distribution
3.4. Correlation between Spatial Distribution and Geometric Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Barrett, S.C.H. Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 8859–8866. [Google Scholar] [CrossRef] [PubMed]
- Hazubska-Przyby, T. Propagation of juniper species by plant tissue culture: A mini-review. Forests 2019, 10, 1028. [Google Scholar] [CrossRef]
- Douterlungne, D.; Ferguson, B.G.; Siddique, I.; Soto-Pinto, L.; Jímenez-Ferrer, G.; Gavito, M.E. Microsite determinants of variability in seedling and cutting establishment in tropical forest restoration plantations. Restor. Ecol. 2015, 23, 861–871. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New For. 2017, 49, 1–34. [Google Scholar] [CrossRef]
- Shahzad, Z.; Amtmann, A. Food for thought: How nutrients regulate root system architecture. Curr. Opin. Plant Biol. 2017, 39, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Stotz, G.C.; Salgado-Luarte, C.; Escobedo, V.M.; Valladares, F. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef]
- Asaah, E.K.; Wanduku, T.N.; Tchoundjeu, Z.; Kouodiekong, L.; Damme, P.V. Do propagation methods affect the fine root architecture of African plum (Dacryodes edulis). Trees 2012, 26, 1461–1469. [Google Scholar] [CrossRef]
- Albrecht, U.; Bordas, M.; Lamb, B.; Meyering, B. Influence of propagation method on root architecture and other traits of young Citrus rootstock plants. HortScience 2017, 52, 1569–1576. [Google Scholar] [CrossRef]
- Hassan, A.; Balachandran, P.; Khamis, K.R. Early root development of Eucalyptus pellita F. Muell. seedlings from seed and stem cutting propagation methods at nursery stage. J. For. Res. 2021, 2021, 6624266. [Google Scholar] [CrossRef]
- Amtmann, A.; Bennett, M.J.; Henry, A. Root phenotypes for the future. Plant Cell Environ. 2022, 45, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Yaffar, D.; Cabugao, K.G.; Meier, I.C. Representing root physiological traits in the root economic space framework. New Phytol. 2022, 234, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.H.; Yan, W.; Rashid, M.H.U.; Tigabu, M.; Gilani, M.M.; Zou, X.H.; Wu, P.F. Chinese fir (Cunninghamia Lanceolata) a green gold of China with continues decline in its productivity over the successive rotations: A review. Appl. Ecol. Environ. Res. 2019, 17, 11055–11067. [Google Scholar] [CrossRef]
- Huang, Z.J.; Selvalakshmi, S.; Vasu, D.; Liu, Q.Q.; Cheng, H.; Guo, F.T.; Ma, X.Q. Identification of indicators for evaluating and monitoring the effects of Chinese fir monoculture plantations on soil quality. Ecol. Indic. 2018, 93, 547–554. [Google Scholar]
- Farooq, T.H.; Ma, X.Q.; Rashid, M.H.U.; Wu, W.J.; Xu, J.J.; Tarin, M.W.K.; He, Z.; Wu, P.F. Impact of stand density on soil quality in Chinese Fir (Cunninghamia Lanceolata) monoculture. Appl. Ecol. Environ. Res. 2019, 17, 3553–3566. [Google Scholar] [CrossRef]
- Wang, P.; Diao, F.W.; Yin, L.M.; Huo, C.F. Absorptive roots trait plasticity explains the variation of root foraging strategies in Cunninghamia lanceolata. Environ. Exp. Bot. 2016, 129, 127–135. [Google Scholar] [CrossRef]
- Zou, X.H.; Wu, P.F.; Chen, N.L.; Wang, P.; Ma, X.Q. Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil. Can. J. For. Res. 2015, 45, 402–410. [Google Scholar] [CrossRef]
- Farooq, T.H.; Wu, W.J.; Tigabu, M.; Ma, X.Q.; He, Z.M.; Rashid, M.H.U.; Gilani, M.M.; Wu, P.F. Growth, biomass production and root development of Chinese fir in relation to initial planting density. Forests 2019, 10, 236. [Google Scholar] [CrossRef]
- Cao, J.L.; Xie, L.; Zheng, Y.X.; Yang, Y.S. Drought intensify the effects of warming on root-colonizing arbuscular mycorrhizal fungal community in subtropical Chinese fir plantation. For. Ecol. Manag. 2020, 464, 118078. [Google Scholar] [CrossRef]
- Wu, P.F.; Ma, X.Q.; Tigabu, M.; Wang, C.; Liu, A.Q.; Odén, P.C. Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress. Can. J. For. Res. 2011, 41, 228–234. [Google Scholar] [CrossRef]
- Xiong, Q.; Huang, X.Y. Speed Tree-Based Forest Simulation System. In Proceedings of the 2010 International Conference on Electrical and Control Engineering, Wuhan, China, 25–27 June 2010. [Google Scholar]
- Walk, T.C.; Erp, E.V.; Lynch, J.P. Modelling applicability of fractal analysis to efficiency of soil exploration by roots. Ann. Bot. 2004, 94, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitter, A.H. An architectural approach to the comparative ecology of plant root systems. New Phytol. 1987, 106, 61–77. [Google Scholar] [CrossRef]
- Bouma, T.J.; Nielsen, K.L.; Hal, J.V.; Koutstaal, B. Root system topology and diameter distribution of species from habitats differing in inundation frequency. Funct. Ecol. 2001, 15, 360–369. [Google Scholar] [CrossRef]
- Tiwari, M.; Kumar, R.; Min, D.; Krishna Jagadish, S.V. Genetic and molecular mechanisms underlying root architecture and function under heat stress—A hidden story. Plant Cell Environ. 2022, 45, 771–788. [Google Scholar] [CrossRef]
- Ma, Z.Q.; Guo, D.L.; Xu, X.L.; Lu, M.Z.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef]
- Kong, D.L.; Ma, C.G.; Zhang, Q.; Li, L.; Chen, X.Y.; Zeng, H.; Guo, D.L. Leading dimensions in absorptive root trait variation across 96 subtropical forest species. New Phytol. 2014, 203, 863–872. [Google Scholar] [CrossRef]
- Napsucialy-mendivil, S.; Dubrovsky, J.G. Genetic and phenotypic analysis of lateral root development in Arabidopsis thaliana. Methods Mol. Biol. 2018, 1761, 47–75. [Google Scholar]
- Van der Bom, F.J.T.; Williams, A.; Bell, M.J. Root architecture for improved resource capture: Tradeoffs in complex environments. J. Exp. Bot. 2020, 71, 5752–5763. [Google Scholar] [CrossRef]
- Mccarthy, M.C.; Enquist, B.J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Li, S.W. Molecular bases for the regulation of adventitious root generation in plants. Front. Plant Sci. 2021, 12, 614072. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Kajala, K. Convergent evolution of gene regulatory networks underlying plant adaptations to dry environments. Plant Cell Environ. 2021, 44, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Eliyahu, A.; Duman, Z.; Sherf, S.; Genin, O.; Cinnamon, Y.; Abu-Abied, M.; Weinstain, R.; Dag, A.; Sadot, E. Vegetative propagation of elite eucalyptus clones as food source for honeybees (Apis mellifera); adventitious roots versus callus formation. Israel J. Plant Sci. 2020, 67, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Zhao, X.Y.; Zhao, H.L.; Huang, Y.X.; Zou, X.A. Linking root morphology, longevity and function to root branch order: A case study in three shrubs. Plant Soil 2010, 336, 197–208. [Google Scholar] [CrossRef]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root structure and functioning for efficient acquisition of phosphorus: Matching morphological and physiological traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef]
- Awotedu, B.F.; Omolola, T.O.; Akala, A.O.; Awotedu, O.L.; Olaoti-Laaro, S.O. Vegetative propagation: A unique technique of improving plants growth. World News Nat. Sci. 2021, 35, 83–101. [Google Scholar]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef]
- Waring, B.G.; Powers, J.S. Overlooking what is underground: Root: Shoot ratios and coarse root allometric equations for tropical forests. For. Ecol. Manag. 2017, 385, 10–15. [Google Scholar] [CrossRef]
- Paz, H.; Pineda-García, F.; Pinzón-Pérez, L.F. Root depth and morphology in response to soil drought: Comparing ecological groups along the secondary succession in a tropical dry forest. Oecologia 2015, 179, 551–561. [Google Scholar] [CrossRef]
- Bontpart, T.; Concha, C.; Giuffrida, M.V.; Robertson, L.; Admkie, K.; Degefu, T.; Girma, N.; Tesfaye, K.; Haileselassie, T.; Fikre, A.; et al. Affordable and robust phenotyping framework to analyse root system architecture of soil-grown plants. Plant J. 2020, 103, 2330–2343. [Google Scholar] [CrossRef]
- Farooq, T.H.; Yan, W.; Chen, X.; Shakoor, A.; Rashid, M.H.U.; Gilani, M.M.; He, Z.; Wu, P. Dynamics of canopy development of Cunninghamia lanceolata mid-age plantation in relation to foliar nitrogen and soil quality influenced by stand density. Glob. Ecol. Conserv. 2020, 24, e01209. [Google Scholar] [CrossRef]
- Poorter, H.; Jagodzinski, A.M.; Ruiz-Peinado, R.; Kuyah, S.; Luo, Y.J.; Oleksyn, J.; Usoltsev, V.A.; Buckley, T.N.; Reich, P.B.; Sack, L. How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continents. New Phytol. 2015, 208, 736–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Propagation Methods | Height/cm | Diameter at the Root Collar/cm |
|---|---|---|
| Seed germination | 25.92 ± 2.84b | 0.40 ± 0.05b |
| Tissue culture | 29.20 ± 3.73a | 0.49 ± 0.08a |
| Rooted cutting | 23.10 ± 2.66b | 0.38 ± 0.06b |
| p-value | <0.001 | <0.001 |
| Propagation Methods | Root Depth/cm | Root Width/cm | Root Width/Root Depth |
|---|---|---|---|
| Seed germination | 17.33 ± 1.70a | 10.87 ± 0.80b | 0.72 ± 0.09b |
| Tissue culture | 12.41 ± 1.31b | 17.05 ± 1.33a | 1.59 ± 0.25a |
| Rooted cutting | 15.20 ± 1.57ab | 13.57 ± 0.82b | 1.06 ± 0.16b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Deng, X.; Zhang, T.; Tian, Y.; Ma, X.; Wu, P. Propagation Methods Decide Root Architecture of Chinese Fir: Evidence from Tissue Culturing, Rooted Cutting and Seed Germination. Plants 2022, 11, 2472. https://doi.org/10.3390/plants11192472
Li L, Deng X, Zhang T, Tian Y, Ma X, Wu P. Propagation Methods Decide Root Architecture of Chinese Fir: Evidence from Tissue Culturing, Rooted Cutting and Seed Germination. Plants. 2022; 11(19):2472. https://doi.org/10.3390/plants11192472
Chicago/Turabian StyleLi, Linxin, Xianhua Deng, Ting Zhang, Yunlong Tian, Xiangqing Ma, and Pengfei Wu. 2022. "Propagation Methods Decide Root Architecture of Chinese Fir: Evidence from Tissue Culturing, Rooted Cutting and Seed Germination" Plants 11, no. 19: 2472. https://doi.org/10.3390/plants11192472