
The World Smallest Plants (Wolffia Sp.) as Potential Species for Bioregenerative Life Support Systems in Space



{kind=link}
{kind=link}
Abstract
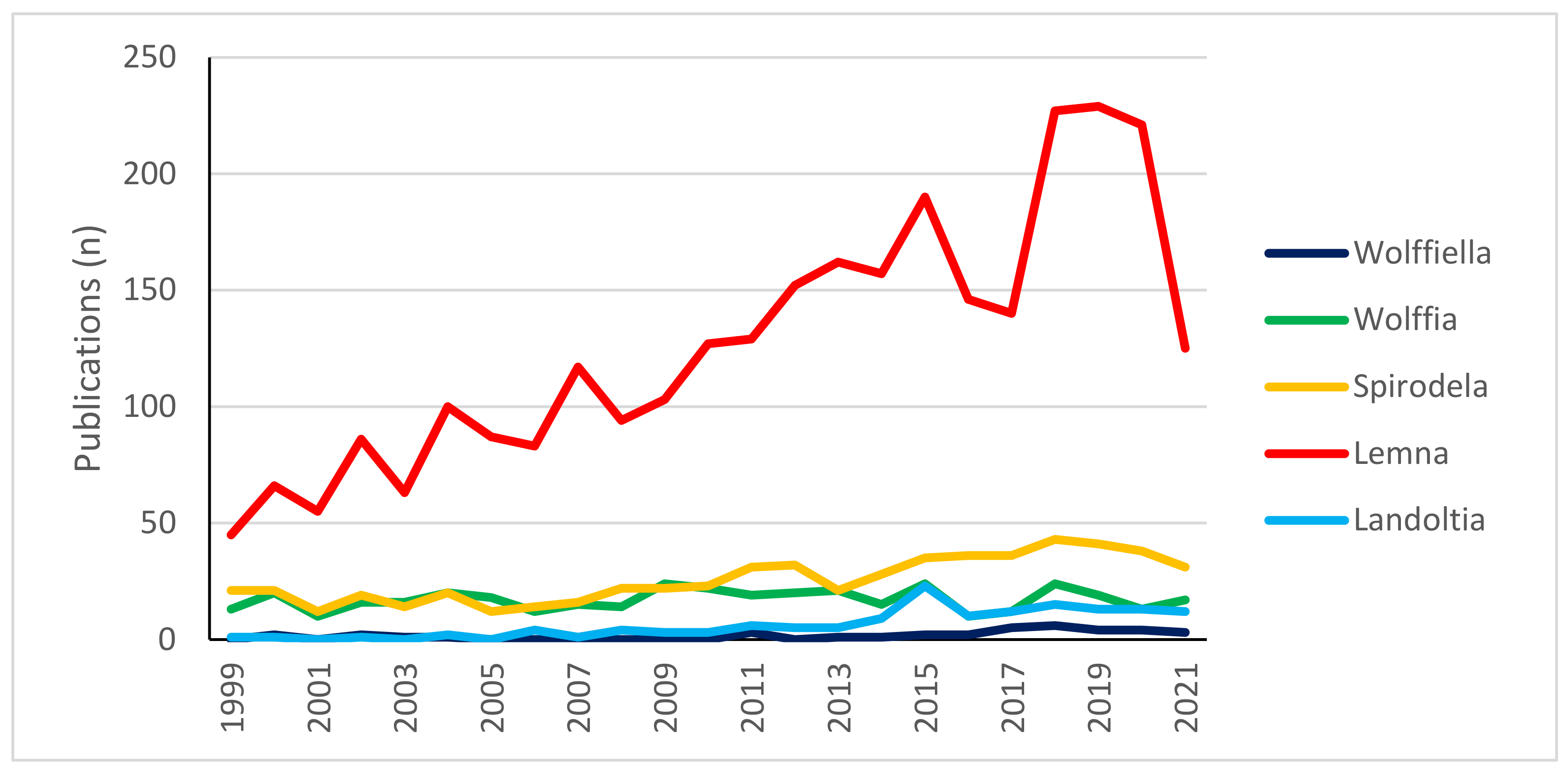
:1. Introduction
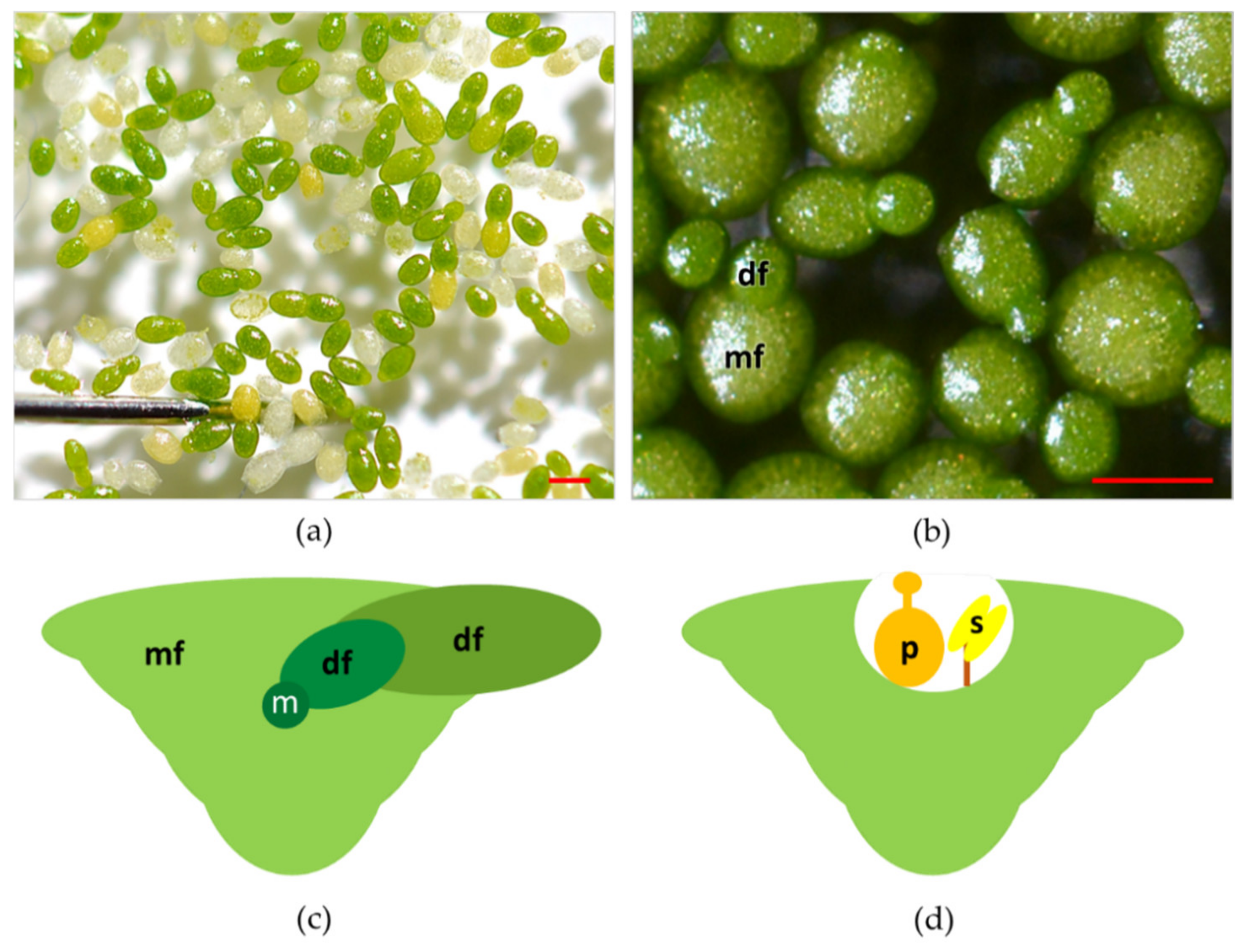
2. Plant Morphological and Reproductive Traits
3. Human Nutrition
4. Biomass Production and Waste Recycling
5. Space Application
6. General Discussion and Critical Insights
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tamponnet, C.; Savage, C. Closed ecological systems. J. Biol. Educ. 1994, 28, 167–174. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G.; Colla, G.; Fortezza, R.; De Pascale, S. Agro-biology for bioregenerative life support systems in long-term space missions: General constraints and the Italian efforts. J. Plant Interactions 2009, 4, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Gòdia, F.; Albiol, J.; Montesinos, J.; Pérez, J.; Creus, N.; Cabello, F.; Mengual, X.; Montras, A.; Lasseur, C. MELISSA: A loop of interconnected bioreactors to develop life support in space. J. Biotechnol. 2002, 99, 319–330. [Google Scholar] [CrossRef]
- De Micco, V.; Buonomo, R.; Paradiso, R.; De Pascale, S.; Aronne, G. Soybean cultivar selection for Bioregenerative Life Support Systems (BLSS)—Theoretical selection. Adv. Space Res. 2012, 49, 1415–1421. [Google Scholar] [CrossRef]
- Paradiso, R.; De Micco, V.; Buonomo, R.; Aronne, G.; Barbieri, G.; De Pascale, S. Soilless cultivation of soybean for Bioregenerative Life-Support Systems: A literature review and the experience of the MELiSSA Project—Food characterisation Phase I. Plant Biol. 2013, 16, 69–78. [Google Scholar] [CrossRef]
- Wheeler, R.M. Agriculture for space: People and places paving the way. Open Agric. 2017, 2, 14–32. [Google Scholar] [CrossRef]
- De Pascale, S.; Arena, C.; Aronne, G.; De Micco, V.; Pannico, A.; Paradiso, R.; Rouphael, Y. Rouphael, biology and crop production in Space environments: Challenges and opportunities. Life Sci. Space Res. 2021, 29, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Dueck, T.; Kempkes, F.; Meinen, E.; Stanghellini, C. Choosing crops for cultivation in space. Contrib. Proc. 2016. [Google Scholar] [CrossRef]
- Tuominen, L.K.; Levine, L.H.; Musgrave, M.E. Plant secondary metabolism in altered gravity. Methods Mol. Biol. 2009, 547, 373–386. [Google Scholar] [CrossRef]
- Fahrion, J.; Mastroleo, F.; Dussap, C.-G.; Leys, N. Use of photobioreactors in regenerative life support systems for human space exploration. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Sree, K.S.; Maheshwari, S.C.; Boka, K.; Khurana, J.P.; Keresztes, Á.; Appenroth, K.J. The duckweed Wolffia microscopica: A unique aquatic monocot. Flora—Morphol. Distrib. Funct. Ecol. Plants 2015, 210, 31–39. [Google Scholar] [CrossRef]
- Landolt, E. Key to the determination of taxa within the family of “Lemnaceae”. Veröffentlichungen Geobot. Inst. Eidg. Tech. Hochsch. Stift Rübel 1980. [Google Scholar] [CrossRef]
- Landolt, E. The Family of Lemnaceae—A Monographic Study. Vol. 1. Biosystematic Investigations in the Family of Duckweeds; Geobotanische Institut ETH: Zurich, Switzerland, 1986. [Google Scholar]
- Bog, M.; Appenroth, K.-J.; Sree, K.S. Duckweed (Lemnaceae): Its molecular taxonomy. Front. Sustain. Food Syst. 2019, 3. [Google Scholar] [CrossRef]
- Bog, M.; Appenroth, K.J.; Sree, K.S. Key to the determination of taxa of Lemnaceae: An update. Nord. J. Bot. 2020, 38. [Google Scholar] [CrossRef]
- Tao, X.; Fang, Y.; Xiao, Y.; Jin, Y.-L.; Ma, X.-R.; Zhao, Y.; He, K.-Z.; Zhao, H.; Wang, H.-Y. Comparative transcriptome analysis to investigate the high starch accumulation of duckweed (Landoltia punctata) under nutrient starvation. Biotechnol. Biofuels 2013, 6, 72. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Appenroth, relative in vitro growth rates of duckweeds (Lemnaceae)—The most rapidly growing higher plants. Plant Biol. 2014, 17, 33–41. [Google Scholar] [CrossRef]
- Bog, M.; Schneider, P.; Hellwig, F.; Sachse, S.; Kochieva, E.; Martyrosian, E.; Landolt, E.; Appenroth, K.-J. Genetic characterisation and barcoding of taxa in the genus Wolffia Horkel ex Schleid. (Lemnaceae) as revealed by two plastidic markers and amplified fragment length polymorphism (AFLP). Planta 2012, 237, 1–13. [Google Scholar] [CrossRef]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.-J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 1–7. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Sree, K.S.; Bog, M.; Ecker, J.; Seeliger, C.; Böhm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Banasch, K.; et al. Nutritional value of the duckweed species of the genus wolffia (Lemnaceae) as human food. Front. Chem. 2018, 6, 483. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Mori, K.; Kodera, T. Nutrient removal and starch production through cultivation of Wolffia arrhiza. J. Biosci. Bioeng. 1999, 87, 194–198. [Google Scholar] [CrossRef]
- Bluem, V.; Paris, F. Novel aquatic modules for bioregenerative life-support systems based on the closed equilibrated biological aquatic system (C.E.B.A.S.). Acta Astronaut. 2002, 50, 775–785. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, F.-J.; Huang, Q.; Williams, P.; Sun, G.; Zhu, Y. Arsenic uptake and speciation in the rootless duckweed Wolffia globosa. New Phytol. 2009, 182, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Prakash, D. Duckweed: An effective tool for phyto-remediation. Toxicol. Environ. Chem. 2013, 95, 1256–1266. [Google Scholar] [CrossRef]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell 2021. [Google Scholar] [CrossRef]
- Landolt, E. Lemna yungensis, a new duckweed species from rocks of the Andean Yungas in Bolivia. Bull. Geobot. Inst. ETH 1998, 64. [Google Scholar] [CrossRef]
- Hillman, W.S. The lemnaceae, or duckweeds. Bot. Rev. 1961, 27, 221–287. [Google Scholar] [CrossRef]
- Anderson, J.L.; Thomson, W.W.; Swader, J.A. Fine structure of Wolffia arrhiza. Can. J. Bot. 1973, 51, 1619–1622. [Google Scholar] [CrossRef]
- Leng, R.A. Duckweed: A Tiny Aquatic Plant with Enormous Potential for Agriculture and Environment; FAO: Roma, Italy, 1999. [Google Scholar]
- Cedergreen, N.; Madsen, T.V. Nitrogen uptake by the floating macrophyte Lemna minor. New Phytol. 2002, 155, 285–292. [Google Scholar] [CrossRef]
- Takai, Y.; Mishima, D.; Kuniki, M.; Sei, K.; Soda, S.; Ike, M. Ethanol production from vegetative fronds and turions of Wolffia arrhiza. Jpn. J. Water Treat. Biol. 2014, 50, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Appenroth, K.J.; Teller, S.; Horn, M. Photophysiology of turion formation and germination in Spirodela polyrhiza. Biol. Plant. 1996, 38, 95–106. [Google Scholar] [CrossRef]
- Fourounjian, P.; Slovin, J.; Messing, J. Flowering and seed production across the lemnaceae. Int. J. Mol. Sci. 2021, 22, 2733. [Google Scholar] [CrossRef]
- Barrett, S.C.; Eckert, C.G.; Husband, B.C. Evolutionary processes in aquatic plant populations. Aquat. Bot. 1993, 44, 105–145. [Google Scholar] [CrossRef]
- Crawford, D.J.; Landolt, E. Allozyme studies in Spirodela (Lemnaceae): Variation among conspecific clones and divergence among the species. Syst. Bot. 1993, 18, 389. [Google Scholar] [CrossRef]
- Park, H.; Park, J.H.; Jeon, H.H.; Woo, D.U.; Lee, Y.; Kang, Y.J. Characterisation of the complete chloroplast genome sequence of Wolffia globosa (Lemnoideae) and its phylogenetic relationships to other Araceae family. Mitochondrial DNA Part B 2020, 5, 1905–1907. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Park, J.H.; Lee, Y.; Woo, D.U.; Jeon, H.H.; Sung, Y.W.; Shim, S.; Kim, S.H.; Lee, K.O.; Kim, J.-Y.; et al. Genome of the world’s smallest flowering plant, Wolffia australiana, helps explain its specialised physiology and unique morphology. Commun. Biol. 2021, 4, 900. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Messing, J. High-throughput sequencing of three lemnoideae (Duckweeds) chloroplast genomes from total DNA. PLoS ONE 2011, 6, e24670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, T.P.; Ernst, E.; Hartwick, N.; Chu, P.; Bryant, D.; Gilbert, S.; Ortleb, S.; Baggs, E.L.; Sree, K.S.; Appenroth, K.J.; et al. Genome and time-of-day transcriptome of Wolffia australiana link morphological minimisation with gene loss and less growth control. Genome Res. 2020, 31, 225–238. [Google Scholar] [CrossRef]
- Boehm, R.; Kruse, C.; Voeste, D.; Barth, S.; Schnabl, H. A transient transformation system for duckweed (Wolffia columbiana) using Agrobacterium-mediated gene transfer. J. Appl. Bot. 2001, 75, 107–111. [Google Scholar]
- Kruse, C.; Boehm, R.; Voeste, D.; Barth, S.; Schnabl, H. Transient transformation of Wolffia columbiana by particle bombardment. Aquat. Bot. 2002, 72, 175–181. [Google Scholar] [CrossRef]
- Pham, T.L.T.; Nguyen, H.A.; Pham, T.H.; Nguyen, T.H.; Le, H.H. Improvement of transformation procedure into duckweed (Wolffia sp.) via Agrobacterium tumefaciens. Tapchi’Congnghe Sinhhoc 2010, 8, 53–60. [Google Scholar]
- Khvatkov, P.; Chernobrovkina, M.; Okuneva, A.; Pushin, A.; Dolgov, S. Transformation of Wolffia arrhiza (L.) Horkel ex Wimm. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 123, 299–307. [Google Scholar] [CrossRef]
- Heenatigala, P.P.M.; Yang, J.; Bishopp, A.; Sun, Z.; Li, G.; Kumar, S.; Hu, S.; Wu, Z.; Lin, W.; Yao, L.; et al. Development of efficient protocols for stable and transient gene transformation for Wolffia globosa using agrobacterium. Front. Chem. 2018, 6, 227. [Google Scholar] [CrossRef] [Green Version]
- Khvatkov, P.; Firsov, A.; Shvedova, A.; Kozlov, O.; Chernobrovkina, M.; Pushin, A.; Shaloiko, L.; Dolgov, S. Wolffia arrhiza as a promising producer of recombinant hirudin. 3 Biotech 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.; Mehta, R.L. Anticoagulation strategies for continuous renal replacement therapies. Crit. Care Nephrol. 2009, 1342–1349. [Google Scholar] [CrossRef]
- Bhanthumnavin, K.; McGarry, M.G. Wolffia arrhiza as a possible source of inexpensive protein. Nature 1971, 232, 495. [Google Scholar] [CrossRef] [PubMed]
- Rusoff, L.L.; Blakeney, E.W.; Culley, D.D. Duckweeds (Lemnaceae family): A potential source of protein and amino acids. J. Agric. Food Chem. 1980, 28, 848–850. [Google Scholar] [CrossRef]
- Skillicorn, P.; Spira, W.; Journey, W. Duckweed Aquaculture: A New Aquatic Farming System for Developing Countries; The World Bank: Washington, DC, USA, 1993. [Google Scholar]
- Appenroth, K.-J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional value of duckweeds (Lemnaceae) as human food. Food Chem. 2017, 217, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Human requirement for N-3 polyunsaturated fatty acids. Poult. Sci. 2000, 79, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Hiemeyer, R.; Amadò, R.; Marti, U. Proteingehalt, aminosäurezusammensetzung und neutralzuckergehalt von lemnaceen: Vorläufige mitteilung. Veröffentlichungen Geobot. Instit. Eidg. Tech. Hochsch. Stift. Rübel Zürich 1980. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Augsten, H.; Liebermann, B.; Feist, H. Feist, effects of light quality on amino acid composition of proteins in Wolffia arrhiza (L.) Wimm. using a specially modified bradford method. Biochem. Physiol. Pflanz. 1982, 177, 251–258. [Google Scholar] [CrossRef]
- Tang, J.; Li, Y.; Ma, J.; Cheng, J.J. Cheng, survey of duckweed diversity in Lake Chao and total fatty acid, triacylglycerol, profiles of representative strains. Plant Biol. 2015, 17, 1066–1072. [Google Scholar] [CrossRef]
- Yan, Y.; Candreva, J.; Shi, H.; Ernst, E.; Martienssen, R.; Schwender, J.; Shanklin, J. Shanklin, survey of the total fatty acid and triacylglycerol composition and content of 30 duckweed species and cloning of a Δ6-desaturase responsible for the production of γ-linolenic and stearidonic acids in Lemna gibba. BMC Plant Biol. 2013, 13, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.; Adams, W.W.I.; Escobar, C.M.; López-Pozo, M.; Demmig-Adams, B. Growth and essential carotenoid micronutrients in Lemna gibba as a function of growth light intensity. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.; Adams, W.; López-Pozo, M.; Garcia, N.D.; McNamara, M.; Escobar, C.; Demmig-Adams, B. Demmig-Adams, features of the duckweed Lemna that support rapid growth under extremes of light intensity. Cells 2021, 10, 1481. [Google Scholar] [CrossRef] [PubMed]
- Pagliuso, D.; Jara, C.E.P.; Grandis, A.; Lam, E.; Ferreira, M.J.P.; Buckeridge, M.S. Flavonoids from duckweeds: Potential applications in the human diet. RSC Adv. 2020, 10, 44981–44988. [Google Scholar] [CrossRef]
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of secondary metabolites in plants: An evolutionary perspective. Trends Plant Sci. 2000, 5, 439–445. [Google Scholar] [CrossRef]
- Plazonić, A.; Bucar, F.; Maleš, Ž.; Mornar, A.; Nigovic, B.; Kujundžić, N. Identification and quantification of flavonoids and phenolic acids in burr parsley (Caucalis platycarpos L.), using high-performance liquid chromatography with diode array detection and electrospray ionisation mass spectrometry. Molecules 2009, 14, 2466–2490. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. Science 2016, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sela, I.; Meir, A.Y.; Brandis, A.; Krajmalnik-Brown, R.; Zeibich, L.; Chang, D.; Dirks, B.; Tsaban, G.; Kaplan, A.; Rinott, E.; et al. Wolffia globosa–Mankai plant-based protein contains bioactive vitamin B12 and is well absorbed in humans. Nutrients 2020, 12, 3067. [Google Scholar] [CrossRef] [PubMed]
- Kotowska, U.; Piotrowska, A.; Isidorova, A.; Bajguz, A.; Isidorov, V. Gas chromatographic-mass spectrometric investigation of the chemical composition of the aquatic plant Wolffia arrhiza (Lemnaceae). Oceanol. Hydrobiol. Stud. 2013, 42, 181–187. [Google Scholar] [CrossRef]
- Kaplan, A.; Zelicha, H.; Tsaban, G.; Meir, A.Y.; Rinott, E.; Kovsan, J.; Novack, L.; Thiery, J.; Ceglarek, U.; Burkhardt, R.; et al. Protein bioavailability of Wolffia globosa duckweed, a novel aquatic plant—A randomised controlled trial. Clin. Nutr. 2018, 38, 2576–2582. [Google Scholar] [CrossRef]
- Kawamata, Y.; Shibui, Y.; Takumi, A.; Seki, T.; Shimada, T.; Hashimoto, M.; Inoue, N.; Kobayashi, H.; Narita, T. Genotoxicity and repeated-dose toxicity evaluation of dried Wolffia globosa Mankai. Toxicol. Rep. 2020, 7, 1233–1241. [Google Scholar] [CrossRef]
- Sree, K.S.; Dahse, H.-M.; Chandran, J.N.; Schneider, B.; Jahreis, G.; Appenroth, K.J. Duckweed for human nutrition: No cytotoxic and no anti-proliferative effects on human cell lines. Plant Foods Hum. Nutr. 2019, 74, 223–224. [Google Scholar] [CrossRef]
- Hejný, S. Ellias Landolt The family of Lemnaceae—A monographic study—Vol.1 biosystematic investigations in the family of Duckweeds (Lemnaceae) (vol. 2). Folia Geobot. Phytotax 1993, 28, 50. [Google Scholar] [CrossRef] [Green Version]
- Soda, S.; Kawahata, Y.; Takai, Y.; Mishima, D.; Fujita, M.; Ike, M. Kinetics of nutrient removal and biomass production by duckweed Wolffia arrhiza in continuous-flow mesocosms. Ecol. Eng. 2013, 57, 210–215. [Google Scholar] [CrossRef]
- Naumann, B.; Eberius, M.; Appenroth, K.-J. Growth rate based dose-response relationships and EC-values of ten heavy metals using the duckweed growth inhibition test (ISO 20079) with Lemna minor L. clone St. J. Plant Physiol. 2007, 164, 1656–1664. [Google Scholar] [CrossRef]
- Sabliy, L.; Konontsev, S.; Grokhovska, J.; Widomski, M.K.; Łagód, G. Nitrogen removal from fish farms water by Lemna minor and Wolffia arrhiza. Proc. ECOpole 2016, 10. [Google Scholar] [CrossRef]
- Fujita, T.; Nakao, E.; Takeuchi, M.; Tanimura, A.; Ando, A.; Kishino, S.; Kikukawa, H.; Shima, J.; Ogawa, J.; Shimizu, S. Characterisation of starch-accumulating duckweeds, Wolffia globosa, as renewable carbon source for bioethanol production. Biocatal. Agric. Biotechnol. 2016, 6, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Karpiscak, M.M.; Gerba, C.P.; Watt, P.M.; Foster, K.E.; Falabi, J.A. Multi-species plant systems for wastewater quality improvements and habitat enhancement. Water Sci. Technol. 1996, 33, 231–236. [Google Scholar] [CrossRef]
- van der Steen, P.; Brenner, A.; Shabtai, Y.; Oron, G. Improved fecal coliform decay in integrated duckweed and algal ponds. Water Sci. Technol. 2000, 42, 363–370. [Google Scholar] [CrossRef]
- Khairina, Y.; Jog, R.; Boonmak, C.; Toyama, T.; Morikawa, M. Indigenous bacteria, an excellent reservoir of functional plant growth promoters for enhancing duckweed biomass yield on site. Chemosphere 2020, 268, 129247. [Google Scholar] [CrossRef]
- Duong, T.P.; Tiedje, J.M. Nitrogen fixation by naturally occurring duckweed–cyanobacterial associations. Can. J. Microbiol. 1985, 31, 327–330. [Google Scholar] [CrossRef]
- Yoneda, Y.; Yamamoto, K.; Makino, A.; Tanaka, Y.; Meng, X.-Y.; Hashimoto, J.; Shin-Ya, K.; Satoh, N.; Fujie, M.; Toyama, T.; et al. Novel plant-associated acidobacteria promotes growth of common floating aquatic plants, duckweeds. Microorganisms 2021, 9, 1133. [Google Scholar] [CrossRef]
- Gale, J.; Smernoff, D.; Macler, B.; MacElroy, R. Carbon balance and productivity of Lemna gibba, a candidate plant for CELSS. Adv. Space Res. 1989, 9, 43–52. [Google Scholar] [CrossRef]
- Kutlakhmedov, L.A.; Sokirko, D.M.; Grodzinskii, A.L.; Mashinshii, G.S.; Nechitailo, N.I. Investigation of the effects of space flight factors on the emergence from the antibiotic state of turions of the great duckweed. Aerosp. Med. Biol. 1980, 12, 49–54. [Google Scholar]
- Kutlakhmedov, L.A.; Sokirko, D.M.; Grodzinskii, G.S. The effects of simulated weightlessness on the reproductive capacity of the great duckweed in the norm and under irradiation. Aerosp. Med. Biol. 1980, 12, 55–58. [Google Scholar]
- Maccallum, T.; Anderson, G.A.; Poynter, J.E.; Stodieck, L.S.; Klaus, D.M. Autonomous Biological System (ABS) experiments. Biol. Sci. Space 1998, 12, 363–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Xu, K. Effects of simulated microgravity on the performance of the duckweeds Lemna aequinoctialis and Wolffia globosa. Aquat. Bot. 2017, 137, 65–71. [Google Scholar] [CrossRef]
- Facius, R.; Scherer, K.; Strauch, W.; Nevzgodina, L.; Maximova, E.; Akatov, Y. Mortality and morphological anomalies related to the passage of cosmic heavy ions through the smallest flowering aquatic plant Wolffia arrhiza. Adv. Space Res. 1996, 18, 195–204. [Google Scholar] [CrossRef]
- Van Hoeck, A.; Horemans, N.; Nauts, R.; Van Hees, M.; Vandenhove, H.; Blust, R. Lemna minor plants chronically exposed to ionising radiation: RNA-seq analysis indicates a dose rate dependent shift from acclimation to survival strategies. Plant Sci. 2017, 257, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B. The maximisation of the productivity of aquatic plants for use in controlled ecological life support systems (CELSS). Acta Astronaut. 1989, 19, 269–273. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, L.E.; Aronne, G. The World Smallest Plants (Wolffia Sp.) as Potential Species for Bioregenerative Life Support Systems in Space. Plants 2021, 10, 1896. https://doi.org/10.3390/plants10091896
Romano LE, Aronne G. The World Smallest Plants (Wolffia Sp.) as Potential Species for Bioregenerative Life Support Systems in Space. Plants. 2021; 10(9):1896. https://doi.org/10.3390/plants10091896
Chicago/Turabian StyleRomano, Leone Ermes, and Giovanna Aronne. 2021. "The World Smallest Plants (Wolffia Sp.) as Potential Species for Bioregenerative Life Support Systems in Space" Plants 10, no. 9: 1896. https://doi.org/10.3390/plants10091896