
Influence of the Nitrate-N to Ammonium-N Ratio on Relative Growth Rate and Crude Protein Content in the Duckweeds Lemna minor and Wolffiella hyalina

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Analytical Methods
2.3. Statistics
3. Results
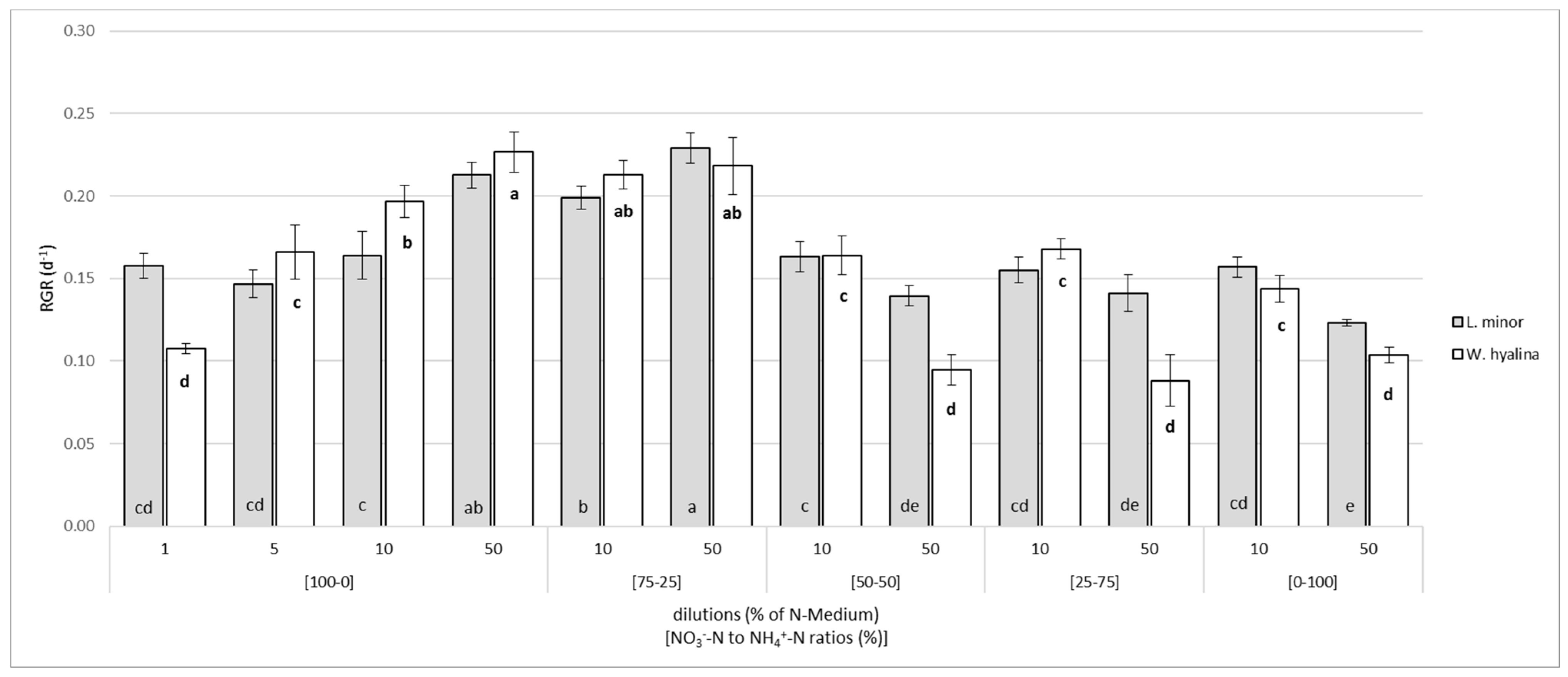
3.1. Growth
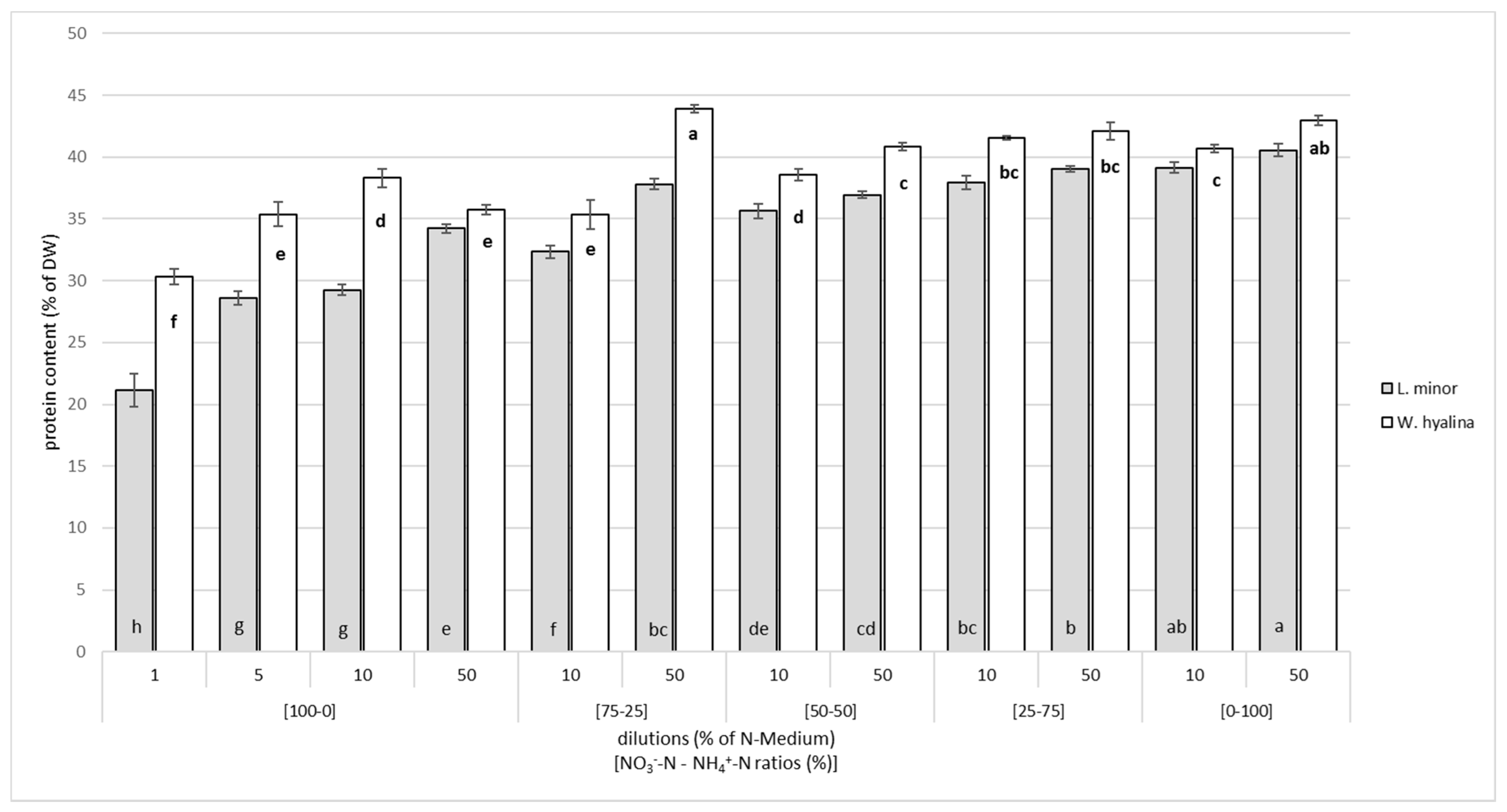
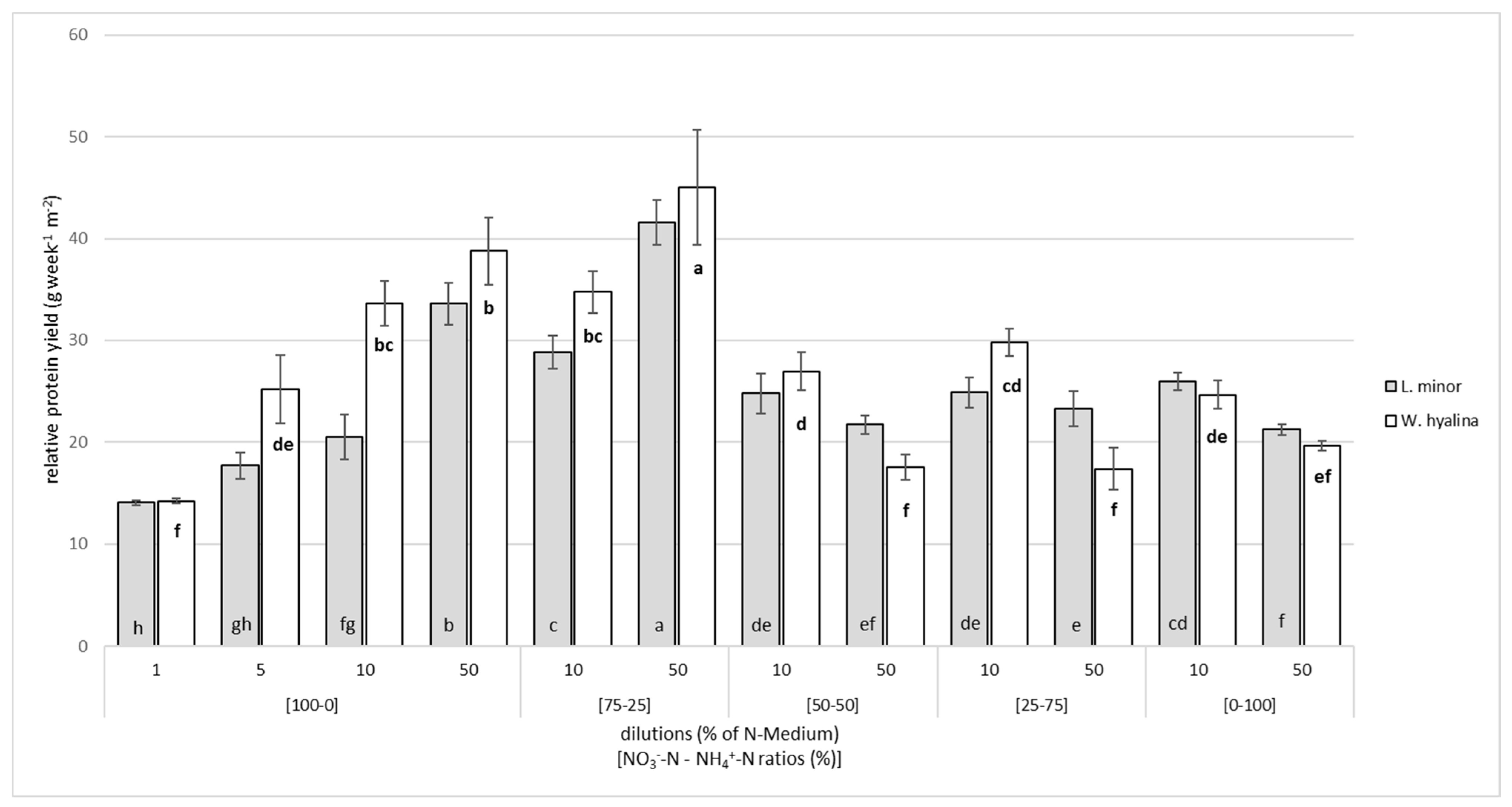
3.2. Crude Protein Content and Protein Yield
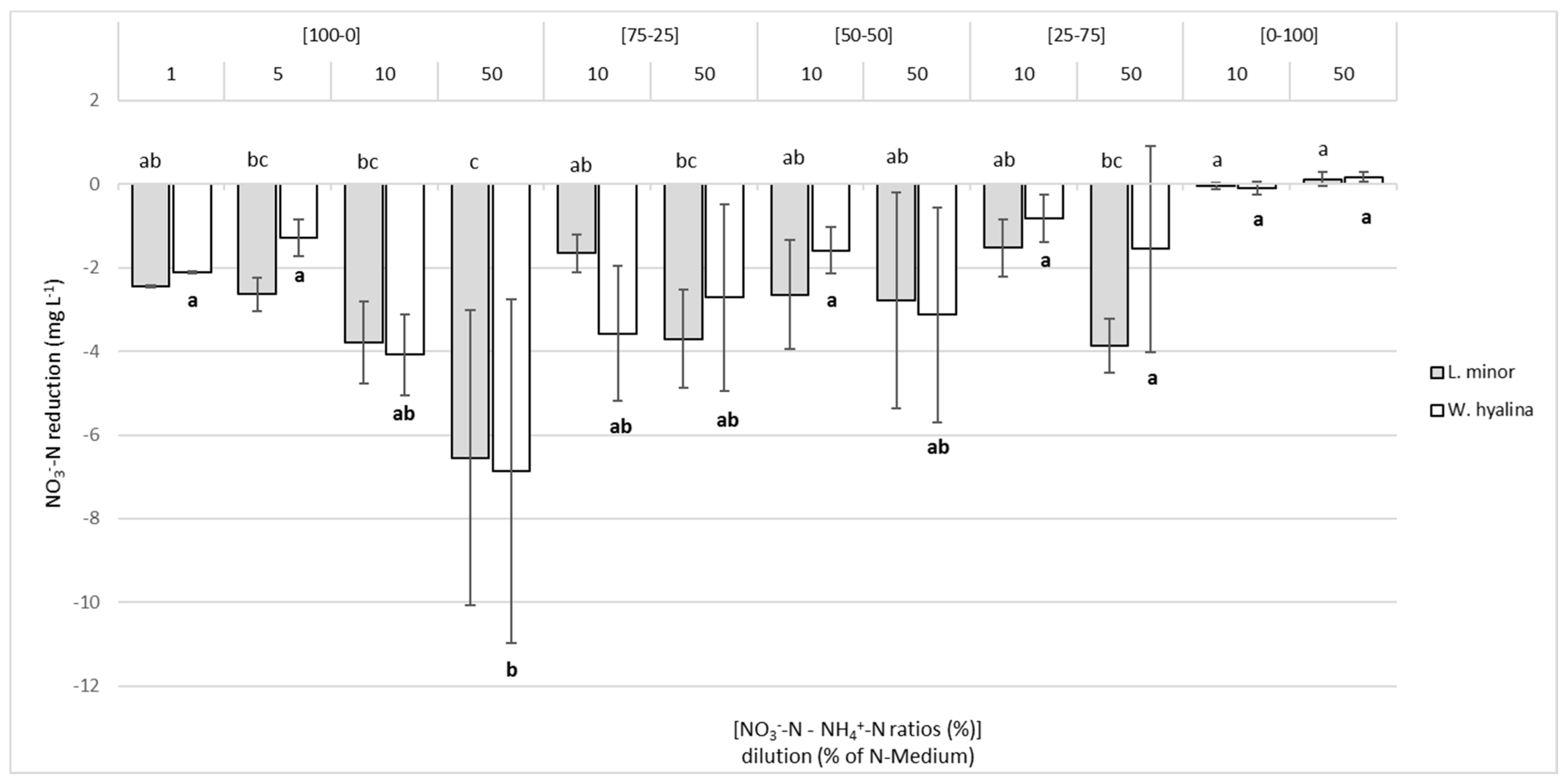
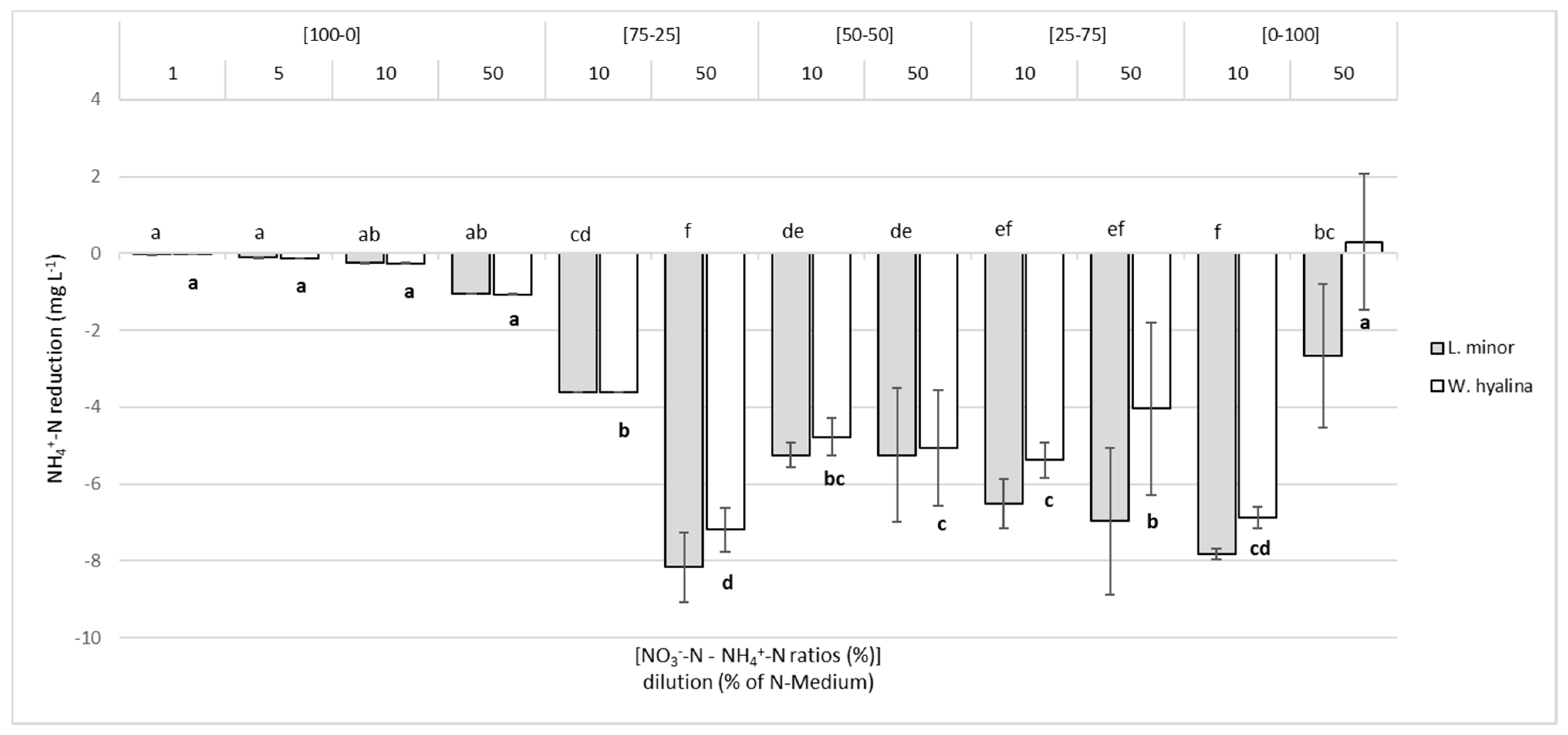
3.3. NO3−-N and NH4+-N Reduction in the Media
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Appenroth, K.J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional value of duckweeds (Lemnaceae) as human food. Food Chem. 2017, 217, 266–273. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sree, K.S.; Bog, M.; Ecker, J.; Seeliger, C.; Böhm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Banasch, K.; et al. Nutritional value of the duckweed species of the genus Wolffia (Lemnaceae) as human food. Front. Chem. 2018, 6, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, R.; Clark, W.D.; Sharma, J.G.; Goswami, R.K.; Shrivastav, A.K.; Tocher, D.R. Mass production of Lemna minor and its amino acid and fatty acid profiles. Front. Chem. 2018, 6, 479. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.J. Relative in vitro growth rates of duckweeds (Lemnaceae)—The most rapidly growing higher plants. Plant Biol. 2015, 17, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant 2015, 37, 204. [Google Scholar] [CrossRef]
- Xu, J.; Cui, W.; Cheng, J.J.; Stomp, A.M. Production of high-starch duckweed and its conversion to bioethanol. Biosyst. Eng. 2011, 110, 67–72. [Google Scholar] [CrossRef]
- Xu, J.; Cheng, J.; Stomp, A.M. Growing Spirodela polyrhiza in swine wastewater for the production of animal feed and fuel ethanol: A Pilot Study. CLEAN Soil Air Water 2012, 40, 760–765. [Google Scholar] [CrossRef]
- Landolt, E.; Kandeler, R. Biosystematic Investigations in the Family of Duckweeds (Lemnaceae)—Vol. 4: The Family of Lemnaceae—A Monographic Study; Geobotanisches Institut ETH: Zürich, Germany, 1987. [Google Scholar]
- Sońta, M.; Rekiel, A.; Batorska, M. Use of duckweed (Lemna L.) in sustainable livestock production and aquaculture—A review. Ann. Anim. Sci. 2019, 19, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Gwaze, F.R.; Mwale, M. The Prospect of Duckweed in Pig Nutrition: A Review. J. Agric. Sci. 2015, 7, 189–199. [Google Scholar] [CrossRef]
- Moss, M.E. Economics and Feed Value of Integrating Duckweed Production with a Swine Operation. Master’s Thesis, Texas Tech University, Lubbock, TX, USA, 1999. [Google Scholar]
- Nguyen, T.K.K.; Ogle, B. Effects of replacing roasted soya beans by broken rice and duckweed on performance of growing Tau Vang chickens confined on-station and scavenging on-farm. Livest. Res. Rural Dev. 2004, 16, 56. [Google Scholar]
- Appenroth, K.J. Media for in vitro-cultivation of duckweed. Duckweed Forum 2015, 3, 180–186. [Google Scholar]
- Appenroth, K.J.; Teller, S.; Horn, M. Photophysiology of turion formation and germination in Spirodela polyrhiza. Biol. Plant 1996, 38, 95–106. [Google Scholar] [CrossRef]
- Fang, Y.Y.; Babourina, O.; Rengel, Z.; Yang, X.E.; Pu, P.M. Ammonium and nitrate uptake by the floating plant Landoltia punctata. Ann. Bot. 2007, 99, 365–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Yang, C.; Tang, X.; Gu, X.; Zhu, Q.; Pan, K.; Hu, Q.; Ma, D. Effects of high ammonium level on biomass accumulation of common duckweed Lemna minor L. Environ. Sci. Pollut. Res. Int. 2014, 21, 14202–14210. [Google Scholar] [CrossRef] [PubMed]
- Caicedo, J.; van der Steen, N.P.; Arce, O.; Gijzen, H.J. Effect of total ammonia nitrogen concentration and pH on growth rates of duckweed (Spirodela polyrhiza). Water Res. 2000, 34, 3829–3835. [Google Scholar] [CrossRef]
- Cedergreen, N.; Madsen, T.V. Nitrogen uptake by the floating macrophyte Lemna minor. New Phytol. 2002, 155, 285–292. [Google Scholar] [CrossRef]
- Simonne, A.H.; Simonne, E.H.; Eitenmiller, R.R.; Mills, H.A.; Cresman, C.P. Could the Dumas method replace the Kjeldahl digestion for nitrogen and crude protein determinations in foods? J. Sci. Food Agric. 1997, 73, 39–45. [Google Scholar] [CrossRef]
- Casal, J.A.; Vermaat, J.E.; Wiegman, F. A test of two methods for plant protein determination using duckweed. Aquat. Bot. 2000, 67, 61–67. [Google Scholar] [CrossRef]
- VDLUFA. Methodenbuch Band 1: Die Untersuchung von Böden, Methode A 6.1.4.1—Bestimmung von Mineralischem Stickstoff (Nitrat und Ammonium) in Bodenprofilen (Nmin-Labormethode); VDLUFA-Verlag: Darmstadt, Germany, 2012. [Google Scholar]
- VDLUFA. Methodenbuch Band 1: Die Untersuchung der Böden, Methode A 6.1.1.1—Bestimmung von Nitrat-Stickstoff durch UV-Absorption; VDLUFA-Verlag: Darmstadt, Germany, 2012. [Google Scholar]
- Iatrou, E.I.; Kora, E.; Stasinakis, A.S. Investigation of biomass production, crude protein and starch content in laboratory wastewater treatment systems planted with Lemna minor and Lemna gibba. Environ. Technol. 2019, 40, 2649–2656. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, Y.-P.; Zhang, T.-T.; Zhao, Y.; Shen, Y.; Huang, L.; Gao, X.; Guo, J.-S. The logistic growth of duckweed (Lemna minor) and kinetics of ammonium uptake. Environ. Technol. 2014, 35, 562–567. [Google Scholar] [CrossRef]
- Mehrer, I.; Mohr, H. Ammonium toxicity: Description of the syndrome in Sinapis alba and the search for its causation. Physiol. Plant. 1989, 77, 545–554. [Google Scholar] [CrossRef]
- Hecht, U.; Mohr, H. Factors controlling nitrate and ammonium accumulation in mustard (Sinapis alba) seedlings. Physiol. Plant. 1990, 78, 379–387. [Google Scholar] [CrossRef]
- Tian, X.; Fang, Y.; Jin, Y.; Yi, Z.; Li, J.; Du, A.; He, K.; Huang, Y.; Zhao, H. Ammonium detoxification mechanism of ammonium-tolerant duckweed (Landoltia punctata) revealed by carbon and nitrogen metabolism under ammonium stress. Environ. Pollut. 2021, 277, 116834. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Islam, R.; Kandwal, P.; Khanam, S.; Proshad, R.; Kormoker, T.; Tusher, T.R. Nitrate transport and assimilation in plants: A potential review. Arch. Agron. Soil Sci. 2020. [Google Scholar] [CrossRef]
- Hao, D.-L.; Zhou, J.-Y.; Yang, S.-Y.; Qi, W.; Yang, K.-J.; Su, Y.-H. Function and Regulation of Ammonium Transporters in Plants. Int. J. Mol. Sci. 2020, 21, 3557. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Augsten, H.; Mattner, A.; Teller, S.; Döhler, G. Effect of UVB irradiation on enzymes of nitrogen metabolism in turions of Spirodela polyrhiza (L.) Schleiden. J. Photochem. Photobiol. B 1993, 18, 215–220. [Google Scholar] [CrossRef]
- Emerson, K.; Russo, R.C.; Lund, R.E.; Thurston, R.V. Aqueous Ammonia Equilibrium Calculations: Effect of pH and Temperature. J. Fish. Res. Board Can. 1975, 32, 2379–2383. [Google Scholar] [CrossRef]
- Liu, C.; Dai, Z.; Sun, H. Potential of duckweed (Lemna minor) for removal of nitrogen and phosphorus from water under salt stress. J. Environ. Manag. 2017, 187, 497–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shammout, M.W.; Zakaria, H. Water lentils (duckweed) in Jordan irrigation ponds as a natural water bioremediation agent and protein source for broilers. J. Ecol. Eng. 2015, 83, 71–77. [Google Scholar] [CrossRef]
- Mohedano, R.A.; Costa, R.H.R.; Tavares, F.A.; Belli Filho, P. High nutrient removal rate from swine wastes and protein biomass production by full-scale duckweed ponds. Bioresour. Technol. 2012, 112, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairina, Y.; Jog, R.; Boonmak, C.; Toyama, T.; Oyama, T.; Morikawa, M. Indigenous bacteria, an excellent reservoir of functional plant growth promoters for enhancing duckweed biomass yield on site. Chemosphere 2021, 268, 129247. [Google Scholar] [CrossRef]
- Hutabarat, R.C.S.M.; Indradewa, D. Effects of water flow rate and surface cover plant density on the growth of duckweed (Lemna minor L.). Ilmu Pertan. Agric. Sci. 2020, 5, 98–109. [Google Scholar] [CrossRef]
- Verma, R.; Suthar, S. Impact of density loads on performance of duckweed bioreactor: A potential system for synchronized wastewater treatment and energy biomass production. Environ. Prog. Sustain. Energy 2015, 34, 1596–1604. [Google Scholar] [CrossRef]
- Färber, E.; Königshofer, H.; Kandeler, R. Ethylene Production and Overcrowding in Lemnaceae. J. Plant Physiol. 1986, 124, 379–384. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stock Solution | Product Name | Main Components | [100-0] (g L−1) | [75-25] (g L−1) | [50-50] (g L−1) | [25-75] (g L−1) | 0-100 (g L−1) |
|---|---|---|---|---|---|---|---|
| 1 | Calcinit | NO3−-N, NH4+-N, Ca+ | 47.2 | 35.4 | 23.6 | 11.8 | 0 |
| 1 | Krista K Plus | NO3−-N, K+ | 161.8 | 121.3 | 80.9 | 40.4 | 0 |
| 2 | NH4Cl | NH4+-N, Cl− | 0 | 0 | 26.7 | 53.5 | 80.2 |
| 3 | OCI Granular 2 | NH4+-N, SO42− | 0 | 33 | 33 | 33 | 33 |
| 4 | KCl | K+, Cl− | 0 | 29.8 | 59.6 | 89.5 | 119.3 |
| 4 | CaCl2 · 2 H20 | Ca+, Cl− | 0 | 7.4 | 14.7 | 22.1 | 29.4 |
| 5 | Krista MKP | PO43−, K+ | 27.2 | 27.2 | 27.2 | 27.2 | 27.2 |
| 6 | Epso Combitop | Mg2+, SO42−, Mn2+, Zn2+ | 49.3 | 49.3 | 49.3 | 49.3 | 49.3 |
| 6 | Borax | BO33− | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| 6 | Mangaan | Mn2+, SO42− | 0.44 | 0.44 | 0.44 | 0.44 | 0.44 |
| 6 | MoNa2O4 · 2 H20 | MoO42−, Na+ | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| 7 | Ferty 72 | Fe3+ | 2.2 | 2.2 | 2.2 | 2.2 | 2.2 |
| NO3−-N to NH4+-N Ratio Substance | [100-0] (mM) | [75-25] (mM) | [50-50] (mM) | [25-75] (mM) | 0-100 (mM) |
|---|---|---|---|---|---|
| NO3−-N | 10.1 | 7.6 | 5.1 | 2.6 | 0.1 |
| NH4+-N | 0 | 2.5 | 5 | 7.5 | 10 |
| PO43− | 1 | 1 | 1 | 1 | 1 |
| K+ | 9.1 | 9.1 | 9.1 | 9.1 | 9.1 |
| Mg2+ | 1.3 | 1.3 | 1.3 | 1.3 | 1.3 |
| SO42− | 2.0 | 3.2 | 3.2 | 3.2 | 3.2 |
| Ca+ | 2.2 | 2.2 | 2.2 | 2.2 | 2.2 |
| Cl− | 0.9 | 3.4 | 8.4 | 13.4 | 18.4 |
| Fe3+ | 0.025 | 0.025 | 0.025 | 0.025 | 0.025 |
| BO33− | 0.005 | 0.005 | 0.005 | 0.005 | 0.005 |
| Mn2+ | 0.013 | 0.013 | 0.013 | 0.013 | 0.013 |
| Zn2+ | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| MoO42− | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 |
| Na+ | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| Ratio | [100-0] | [75-25] | [50-50] | [25-75] | 0-100 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dilution (%) | 1 | 5 | 10 | 50 | 10 | 50 | 10 | 50 | 10 | 50 | 10 | 50 |
| NO3−-N (mg L−1) | 2.7 | 8.8 | 15.3 | 71.2 | 12.1 | 56.7 | 10.1 | 35.3 | 5.2 | 16.6 | 1.1 | 1.4 |
| NH4+-N (mg L−1) | 0.06 | 0.17 | 0.29 | 1.1 | 3.6 | 17.3 | 7.1 | 32.5 | 10.7 | 51.3 | 14.4 | 64.4 |
| EC (mS cm−1) | 0.43 | 0.46 | 0.53 | 1.15 | 0.6 | 1.33 | 0.66 | 1.58 | 0.65 | 1.64 | 0.64 | 1.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petersen, F.; Demann, J.; Restemeyer, D.; Ulbrich, A.; Olfs, H.-W.; Westendarp, H.; Appenroth, K.-J. Influence of the Nitrate-N to Ammonium-N Ratio on Relative Growth Rate and Crude Protein Content in the Duckweeds Lemna minor and Wolffiella hyalina. Plants 2021, 10, 1741. https://doi.org/10.3390/plants10081741
Petersen F, Demann J, Restemeyer D, Ulbrich A, Olfs H-W, Westendarp H, Appenroth K-J. Influence of the Nitrate-N to Ammonium-N Ratio on Relative Growth Rate and Crude Protein Content in the Duckweeds Lemna minor and Wolffiella hyalina. Plants. 2021; 10(8):1741. https://doi.org/10.3390/plants10081741
Chicago/Turabian StylePetersen, Finn, Johannes Demann, Dina Restemeyer, Andreas Ulbrich, Hans-Werner Olfs, Heiner Westendarp, and Klaus-Jürgen Appenroth. 2021. "Influence of the Nitrate-N to Ammonium-N Ratio on Relative Growth Rate and Crude Protein Content in the Duckweeds Lemna minor and Wolffiella hyalina" Plants 10, no. 8: 1741. https://doi.org/10.3390/plants10081741