
Pre- and/or Postharvest Silicon Application Prolongs the Vase Life and Enhances the Quality of Cut Peony (Paeonia lactiflora Pall.) Flowers




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Silicon Treatments and Experimental Design
2.3. Growth Attribute Measurements
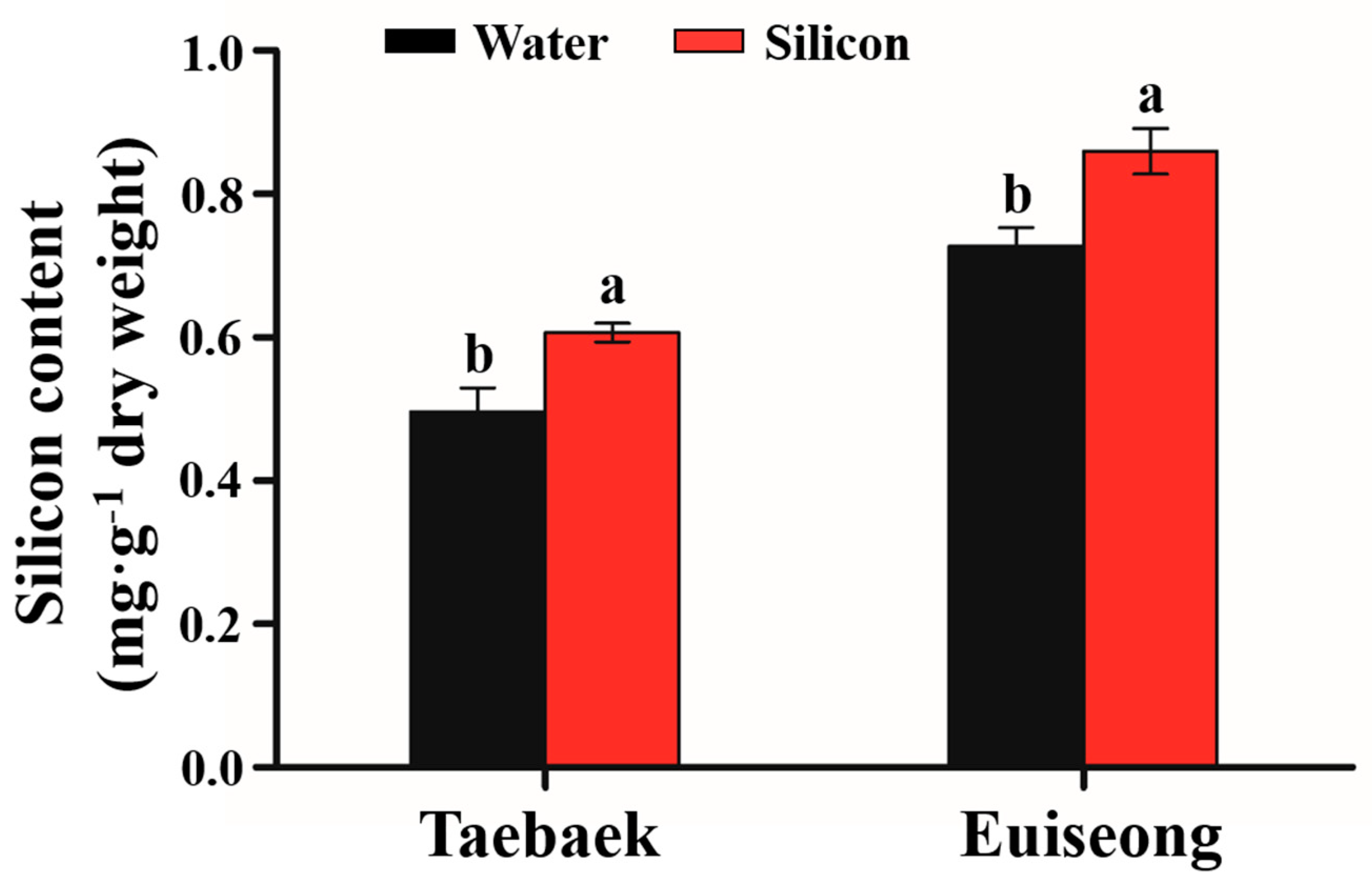
2.4. Determination of the Si Contents
2.5. Definitions of Postharvest Flower Opening Stages for ‘Taebaek’ and ‘Euiseong’
2.6. Measurements of Flowering Stem Fresh Weight during the Vase Life
2.7. Quantification of the Antioxidant Enzyme Activities
2.8. Stem Strength Determination
2.9. Statistical Analysis
3. Results
3.1. Growth and Development of Peony Plants as Affected by Si
3.2. Si Contents in the Harvested Peony
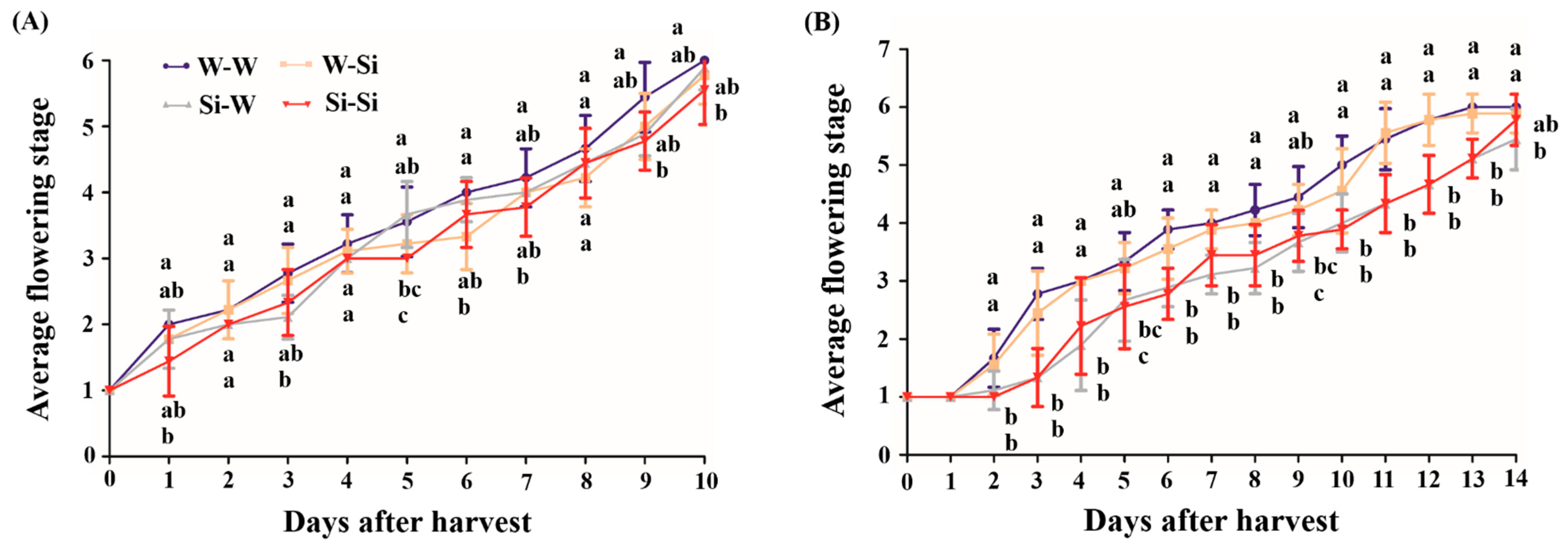
3.3. Vase Life of Postharvest Peony
3.4. Fresh Weight of Postharvest Peony
3.5. Antioxidant Enzyme Activities
3.6. MOE and MOR of Postharvest Peony
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, D.; Hao, Z.; Tao, J. Effects of shade on plant growth and flower quality in the herbaceous peony (Paeonia lactiflora Pall.). Plant Physiol. Biochem. 2012, 61, 187–196. [Google Scholar] [CrossRef]
- Holloway, P.S.; Buchholz, K. The State of the Alaska Peony Industry 2012; Agricultural and Forestry Experiment Station, School of Agriculture and Land Resources Management, University of Alaska Fairbanks: Fairbanks, AK, USA, 2013; Volume 3, pp. 1–8. [Google Scholar]
- Rhie, Y.H.; Jung, H.H.; Kim, K.S. Chilling requirement for breaking dormancy and flowering in Paeonia lactiflora ‘Taebaek’ and ‘Mulsurae’. Hortic. Environ. Biotechnol. 2012, 53, 277–282. [Google Scholar] [CrossRef]
- Zhao, D.; Cheng, M.; Tang, W.; Liu, D.; Zhou, S.; Meng, J.; Tao, J. Nano-silver modifies the vase life of cut herbaceous peony (Paeonia lactiflora Pall.) flowers. Protoplasma 2018, 255, 1001–1013. [Google Scholar] [CrossRef]
- Walton, E.; Boldingh, H.; McLaren, G.; Williams, M.; Jackman, R. The dynamics of starch and sugar utilisation in cut peony (Paeonia lactiflora Pall.) stems during storage and vase life. Postharvest Biol. Technol. 2010, 58, 142–146. [Google Scholar] [CrossRef]
- Van Doorn, W.G.; Abadie, P.; Belde, P. Alkylethoxylate surfactants for rehydration of roses and Bouvardia flowers. Postharvest Biol. Technol. 2001, 24, 327–333. [Google Scholar] [CrossRef]
- Dole, J.; Stamps, B.; Carlson, A.; Ahmad, I.; Greer, L.; Laushman, J. Postharvest Handling of Cut Flowers and Greens; ASCFG Press: Oberlin, OH, USA, 2017. [Google Scholar]
- Ahmad, I.; Dole, J.M.; Amjad, A.; Ahmad, S. Dry storage effects on postharvest performance of selected cut flowers. HortTechnology 2012, 22, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Hajizadeh, H.S.; Farokhzad, A.; Chelan, V. Using of preservative solutions to improve postharvest life of Rosa Hybrid cv. Black Magic. J. Agric. Technol. 2012, 8, 1817–1826. [Google Scholar]
- Pospíšilová, J.; Teixeira da Silva, J.A. Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues. Volumes I–IV. Biol. Plant. 2007, 51, 390. [Google Scholar] [CrossRef]
- Anjum, M.A.; Naveed, F.; Shakeel, F.; Amin, S. Effect of some chemicals on keeping quality and vase-life of tuberose (Polianthes tuberosa L.) cut flowers. Life 2001, 12, 23–65. [Google Scholar]
- Pozza, E.A.; Pozza, A.A.A.; Botelho, D.M.d.S. Silicon in plant disease control. Rev. Ceres 2015, 62, 323–331. [Google Scholar] [CrossRef]
- Campos, C.N.S.; Silva Júnior, G.B.D.; Prado, R.D.M.; David, C.H.O.D.; Souza Junior, J.P.D.; Teodoro, P.E. Silicon mitigates ammonium toxicity in plants. Agron. J. 2020, 112, 635–647. [Google Scholar] [CrossRef]
- Agarie, S.; Uchida, H.; Agata, W.; Kubota, F.; Kaufman, P.B. Effects of silicon on transpiration and leaf conductance in rice plants (Oryza sativa L.). Plant Prod. Sci. 1998, 1, 89–95. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, J.; Hu, J.; Jeong, B.R. Method of silicon application affects quality of strawberry daughter plants during cutting propagation in hydroponic substrate system. Agronomy 2020, 10, 1753. [Google Scholar] [CrossRef]
- Feng, J.; Shi, Q.; Wang, X.; Wei, M.; Yang, F.; Xu, H. Silicon supplementation ameliorated the inhibition of photosynthesis and nitrate metabolism by cadmium (Cd) toxicity in Cucumis sativus L. Sci. Hortic. 2010, 123, 521–530. [Google Scholar] [CrossRef]
- Chérif, M.; Benhamou, N.; Menzies, J.G.; Bélanger, R. Silicon induced resistance in cucumber plants against Pythium ultimum. Physiol. Mol. Plant Pathol. 1992, 41, 411–425. [Google Scholar] [CrossRef]
- Fortunato, A.A.; Rodrigues, F.Á.; Baroni, J.C.P.; Soares, G.C.B.; Rodriguez, M.A.D.; Pereira, O.L. Silicon suppresses Fusarium wilt development in banana plants. J. Phytopathol. 2012, 160, 674–679. [Google Scholar] [CrossRef]
- Eason, J.; Pinkney, T.; Heyes, J.; Brash, D.; Bycroft, B. Effect of storage temperature and harvest bud maturity on bud opening and vase life of Paeonia lactiflora cultivars. N. Z. J. Crop Hortic. 2002, 30, 61–67. [Google Scholar] [CrossRef]
- Park, Y.G.; Muneer, S.; Kim, S.; Hwang, S.J.; Jeong, B.R. Foliar or subirrigational silicon supply modulates salt stress in strawberry during vegetative propagation. Hortic. Environ. Biotechnol. 2018, 59, 11–18. [Google Scholar] [CrossRef]
- Zhang, H.; Dotson, P. The use of microwave muffle furnace for dry ashing plant tissue samples. Commun. Soil Sci. Plan. 1994, 25, 1321–1327. [Google Scholar] [CrossRef]
- Rabiza-Świder, J.; Skutnik, E.; Jędrzejuk, A.; Łukaszewska, A. Postharvest treatments improve quality of cut peony flowers. Agronomy 2020, 10, 1583. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Arum, L.S.; Ko, C.H.; Muneer, S.; Jeong, B.R. Silicon-mediated enhancement of physiological and biochemical characteristics of Zinnia elegans ‘Dreamland Yellow’ grown under salinity stress. Hortic. Environ. Biotechnol. 2015, 56, 721–731. [Google Scholar] [CrossRef]
- Al-Zube, L.A.; Robertson, D.J.; Edwards, J.N.; Sun, W.; Cook, D.D. Measuring the compressive modulus of elasticity of pith-filled plant stems. Plant Methods 2017, 13, 1–9. [Google Scholar] [CrossRef]
- Onoda, Y.; Richards, A.E.; Westoby, M. The relationship between stem biomechanics and wood density is modified by rainfall in 32 Australian woody plant species. New Phytol. 2010, 185, 493–501. [Google Scholar] [CrossRef]
- Gaur, S.; Kumar, J.; Kumar, D.; Chauhan, D.K.; Prasad, S.M.; Srivastava, P.K. Fascinating impact of silicon and silicon transporters in plants: A review. Ecotoxicol. Environ. Saf. 2020, 202, 110885. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Khan, A.L.; Lee, I.J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef]
- Cherif, M.; Belanger, R. Use of potassium silicate amendments in recirculating nutrient solutions to suppress Pythium ultimum on long English cucumber. Plant Dis. 1992, 76, 1008–1011. [Google Scholar] [CrossRef]
- Zhao, D.; Hao, Z.; Tao, J.; Han, C. Silicon application enhances the mechanical strength of inflorescence stem in herbaceous peony (Paeonia lactiflora Pall.). Sci. Hortic. 2013, 151, 165–172. [Google Scholar] [CrossRef]
- Mar, R.J.; Hassani, A.; Abdollahi, A.; Hanafi, S. Improvement of the vase life of cut gladiolus flowers by essential oils, salicylic acid and silver thiosulfate. J. Med. Plants Res. 2011, 5, 5039–5043. [Google Scholar]
- Kamenidou, S.; Cavins, T.J.; Marek, S. Silicon supplements affect horticultural traits of greenhouse-produced ornafomental sunflowers. HortScience 2008, 43, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Khan, P.; Shahrin, S.; Taufique, T.; Mehraj, H.; Jamal Uddin, A. Prolonging vase life of cut rose (Rosa hybrida L. cv. Red Pearl) through chemical preservatives. J. Biosci. Agric. Res. 2015, 5, 10–15. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Li, Y.; Jeong, B.R. Silicon Alleviates Temperature Stresses in Poinsettia by Regulating Stomata, Photosynthesis, and Oxidative Damages. Agronomy 2020, 10, 1419. [Google Scholar] [CrossRef]
- Gupta, J.; Dubey, R. Factors affecting postharvest life of flower crops. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 548–557. [Google Scholar] [CrossRef]
- Skutnik, E.; Rabiza-Świder, J.; Jędrzejuk, A.; Łukaszewska, A. The effect of the long-term cold storage and preservatives on senescence of cut herbaceous peony flowers. Agronomy 2020, 10, 1631. [Google Scholar] [CrossRef]
- Heuser, C.W.; Evensen, K.B. Cut flower longevity of peony. J. Am. Soc. Hortic. Sci. 1986, 111, 896–899. [Google Scholar]
- Gast, K.; McLaren, J.; Kampjes, R. Identification of bud maturity indicators for fresh-cut peony flowers. Acta Hortic. 2001, 543, 317–325. [Google Scholar] [CrossRef]
- Jaiganesh, V.; Devi Shanthini, V.; Kannan, C.; Darwin Christdhas Henry, L. Role of Silicon Nutrient Mediated Plant Disease Resistance. Curr. Res. Innov. Plant Pathol. 2019, 3, 63–81. [Google Scholar]
- Song, J.N.; Liu, X.H.; Wang, Y.Q.; Yang, H.B. Transcriptome analysis reveals salinity responses in four Tartary buckwheat cultivars. J. Plant Biochem. Biotechnol. 2021, 30, 564–578. [Google Scholar] [CrossRef]
- Markovich, O.; Steiner, E.; Kouřil, Š.; Tarkowski, P.; Aharoni, A.; Elbaum, R. Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plant Cell Environ. 2017, 40, 1189–1196. [Google Scholar] [CrossRef]
- Tiryaki, S.; Hamzaçebi, C. Predicting modulus of rupture (MOR) and modulus of elasticity (MOE) of heat treated woods by artificial neural networks. Measurement 2014, 49, 266–274. [Google Scholar] [CrossRef]
- Hussain, S.; Shuxian, L.; Mumtaz, M.; Shafiq, I.; Iqbal, N.; Brestic, M.; Shoaib, M.; Sisi, Q.; Li, W.; Mei, X. Foliar application of silicon improves stem strength under low light stress by regulating lignin biosynthesis genes in soybean (Glycine max (L.) Merr.). J. Hazard. Mater. 2021, 401, 123256. [Google Scholar] [CrossRef]
- Babalar, M.; Edrisi, B.; Naderi, R. Evaluation of the mechanical strength of gerbera flower stem in response to silicon and salicylic acid application. J. Ornam. Plants 2016, 6, 163–171. [Google Scholar]
- Hwang, S.J.; Park, H.M.; Jeong, B.R. Effects of potassium silicate on the growth of miniature rose ’Pinocchio’ grown on rockwool and its cut flower quality. J. Jpn. Soc. Hortic. Sci. 2005, 74, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Karimian, N.; Nazari, F.; Samadi, S. Morphological and Biochemical Properties, Leaf Nutrient Content, and Vase Life of Tuberose (Polianthes tuberosa L.) Affected by Root or Foliar Applications of Silicon (Si) and Silicon Nanoparticles (SiNPs). J. Plant Growth Regul. 2020, 1–15. [Google Scholar] [CrossRef]
- Geerdink, G.M.; Orsi, B.; Tezotto-Uliana, J.V.; Pessoa, C.O.; Sasaki, F.F.; Kluge, R.A. Pre-harvest silicon treatment improves quality of cut rose stems and maintains postharvest vase life. J. Plant Nutr. 2020, 43, 1418–1426. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar (A) | Treatment (B) | Shoot Length (cm) | Stem Diameter (mm) | Leaf Length (cm) | Leaf Width (cm) | Bud Diameter (mm) |
|---|---|---|---|---|---|---|
| ‘Taebaek’ | Water | 74.13 b z | 3.29 b | 9.58 b | 3.25 b | 23.71 b |
| Si | 83.42 a | 4.41 a | 11.82 a | 4.13 a | 28.62 a | |
| ‘Euiseong’ | Water | 69.42 b | 3.53 a | 8.85 b | 3.22 a | 26.26 a |
| Si | 81.25 a | 3.96 a | 10.52 a | 3.53 a | 27.72 a | |
| F-test | A | * y | NS | ** | NS | NS |
| B | *** | *** | *** | ** | *** | |
| A × B | NS | * | NS | NS | ** |
| (A) MOE and MOR values of the peony ‘Taebaek’ subjected to different treatments | |||||
| Treatment | Modulus of elasticity (MOE, MPa) | Modulus of rupture (MOR, MPa) | |||
| Pre-harvest | Postharvest | Neck | Base | Neck | Base |
| Water | Water | 186.0 d Z | 183.0 d | 8.34 b | 12.13 ab |
| Water | Si | 296.5 bc | 351.1 c | 11.92 ab | 12.03 ab |
| Si | Water | 333.8 ab | 487.7 ab | 12.07 a | 11.95 ab |
| Si | Si | 357.9 a | 454.6 ab | 12.24 a | 13.61 a |
| (B) MOE and MOR values of the peony ‘Euiseong’ subjected to different treatments | |||||
| Treatment | Modulus of elasticity (MOE, MPa) | Modulus of rupture (MOR, MPa) | |||
| Pre-harvest | Postharvest | Neck | Base | Neck | Base |
| Water | Water | 112.1 b | 152.0 c | 7.1 c | 11.3 ab |
| Water | Si | 189.1 a | 334.8 a | 18.6 ab | 15.5 a |
| Si | Water | 138.5 b | 232.2 ab | 11.2 bc | 8.8 b |
| Si | Si | 218.4 a | 331.1 a | 22.1 a | 13.1 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Li, Y.; Hu, J.; Lee, J.; Jeong, B.R. Pre- and/or Postharvest Silicon Application Prolongs the Vase Life and Enhances the Quality of Cut Peony (Paeonia lactiflora Pall.) Flowers. Plants 2021, 10, 1742. https://doi.org/10.3390/plants10081742
Song J, Li Y, Hu J, Lee J, Jeong BR. Pre- and/or Postharvest Silicon Application Prolongs the Vase Life and Enhances the Quality of Cut Peony (Paeonia lactiflora Pall.) Flowers. Plants. 2021; 10(8):1742. https://doi.org/10.3390/plants10081742
Chicago/Turabian StyleSong, Jinnan, Yali Li, Jiangtao Hu, Jaehyeok Lee, and Byoung Ryong Jeong. 2021. "Pre- and/or Postharvest Silicon Application Prolongs the Vase Life and Enhances the Quality of Cut Peony (Paeonia lactiflora Pall.) Flowers" Plants 10, no. 8: 1742. https://doi.org/10.3390/plants10081742