
Lemnaceae and Orontiaceae Are Phylogenetically and Morphologically Distinct from Araceae

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
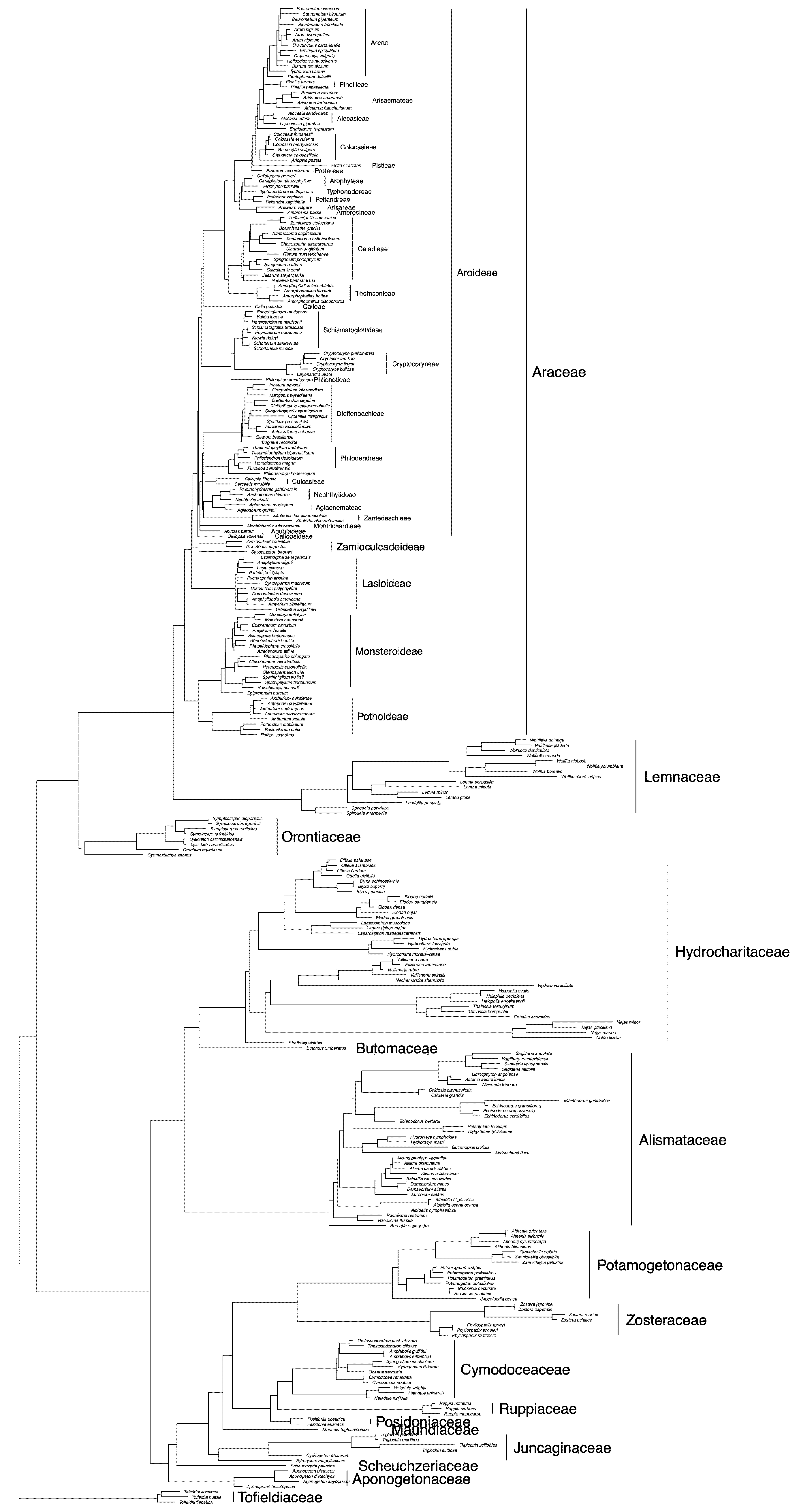
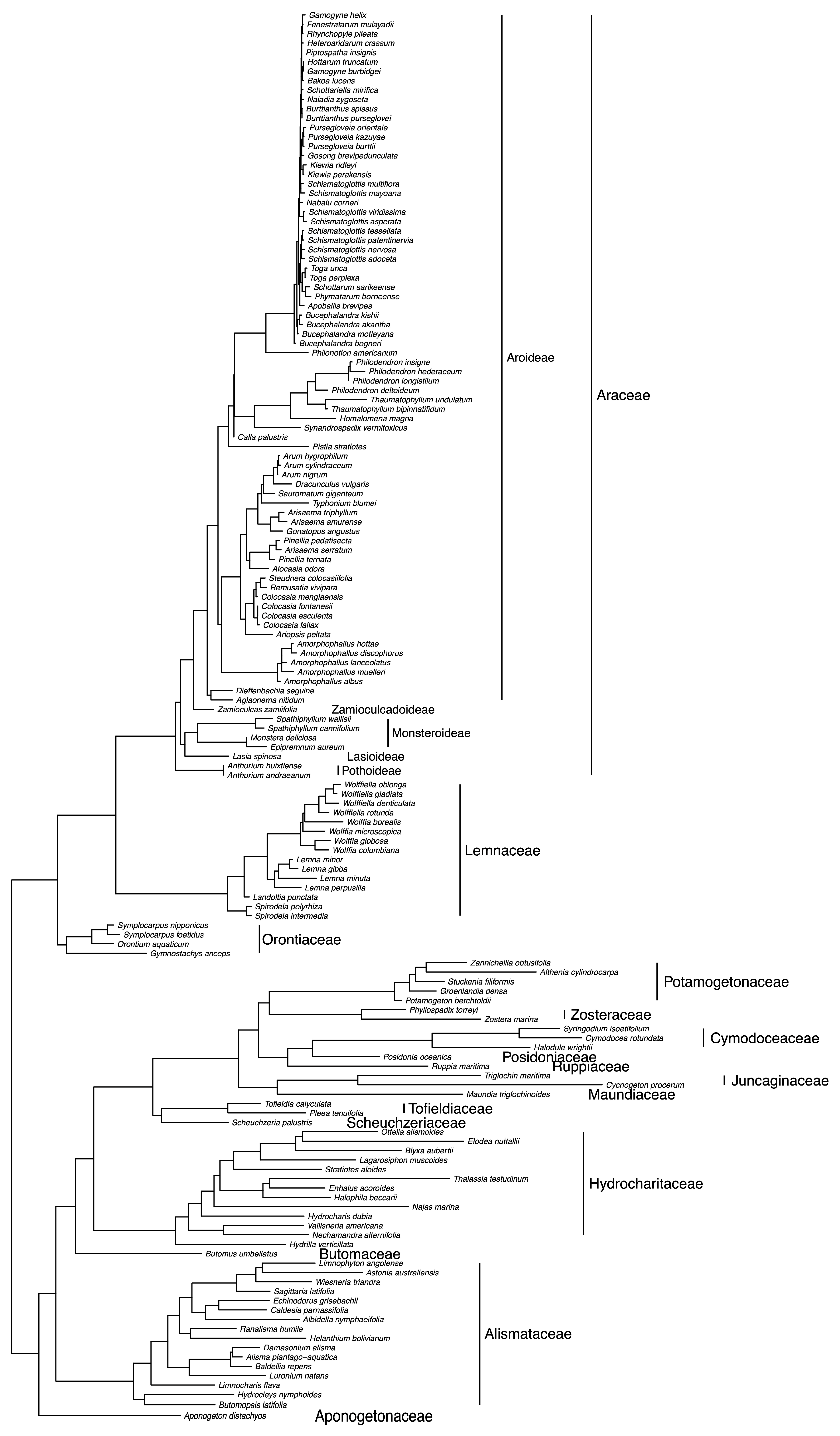
1.1. Taxonomic History of Araceae
1.2. Orontiaceae
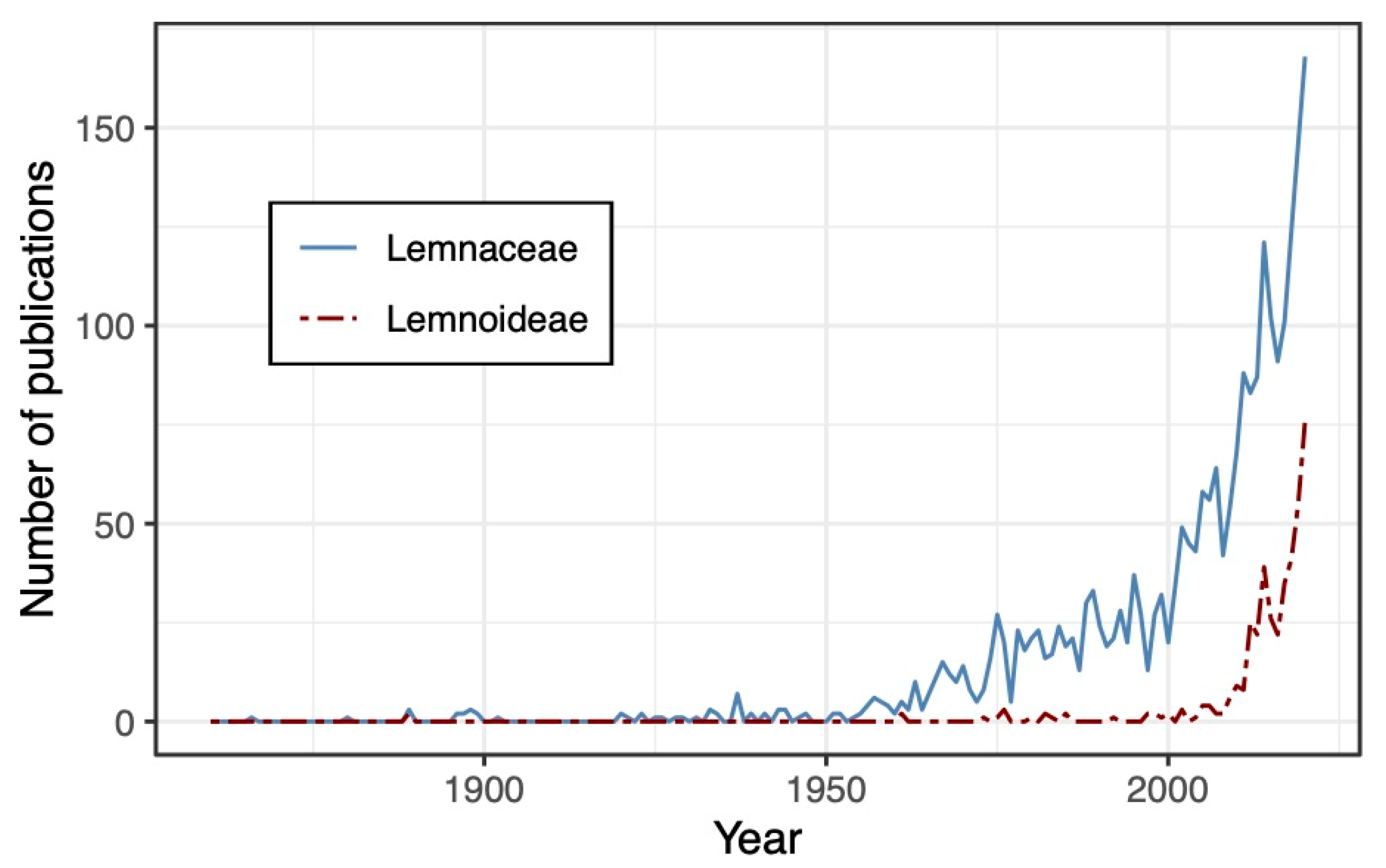
1.3. Lemnaceae
1.4. Related Lineages and Ordinal Classification
1.5. Objectives
2. Molecular Phylogenetic Evidence
2.1. Plastid Molecular Data
2.2. Nuclear Molecular Data
2.3. Mitochondrial Molecular Data
3. Morphological Data
4. Chromosome Number Evolution
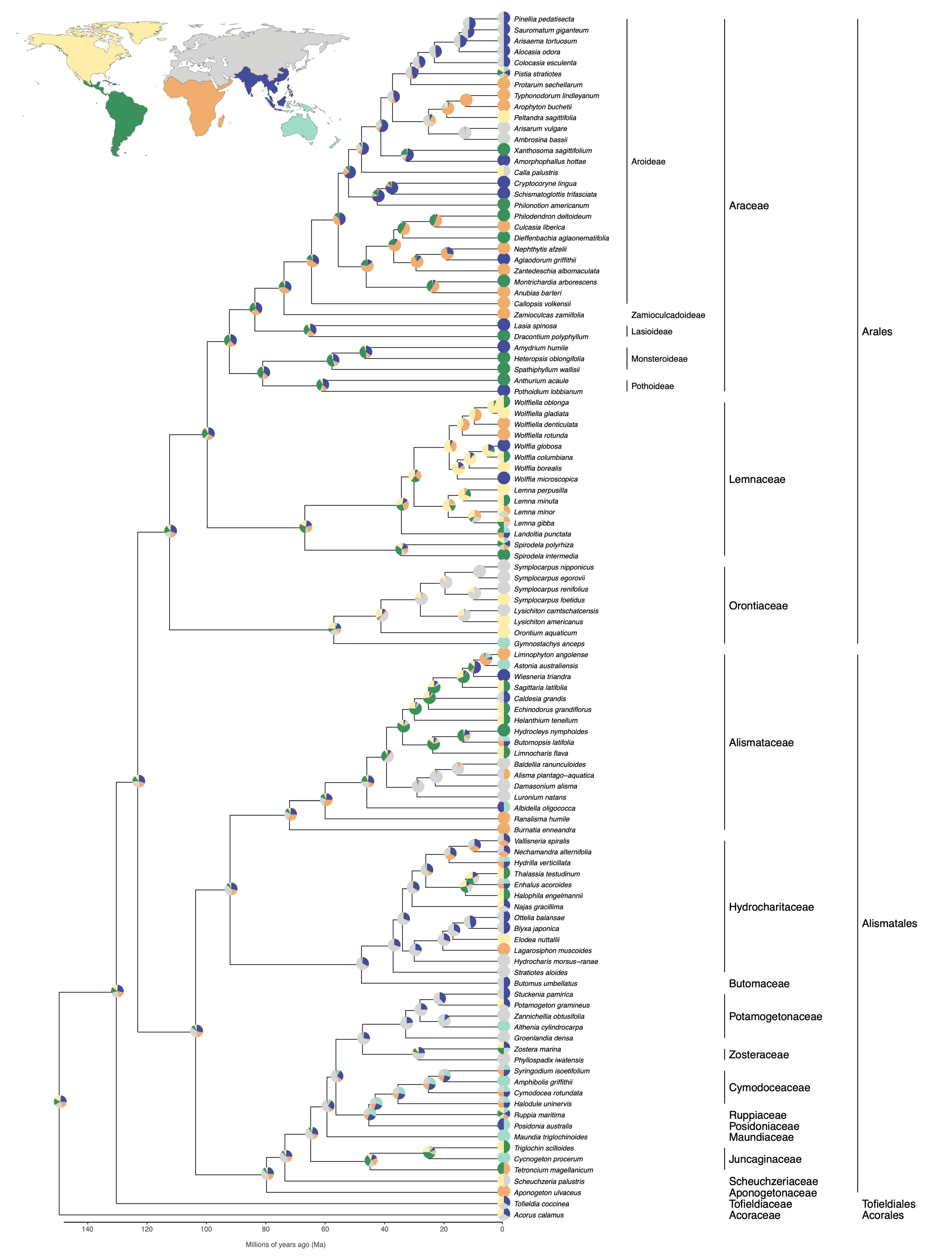
5. Biogeography
6. Discussion
6.1. Nomenclatural Stability and Utility
6.2. The Nature of a Plant Family
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mayo, S.J.; Bogner, J.; Boyce, P.C. The Genera of Araceae; Royal Botanic Gardens: Kew, UK, 1997. [Google Scholar]
- Boyce, P.C.; Croat, T.B. The Überlist of Araceae, Totals for Published and Estimated Number of Species in Aroid Genera. 2018. Available online: http://www.aroid.org/genera/180211uberlist.pdf (accessed on 14 April 2020).
- French, J.C.; Chung, M.G.; Hur, Y.K. Chloroplast DNA phylogeny of the Ariflorae. In Monocotyledons: Systematics and Evolution; Rudall, P.J., Cribb, P.J., Cutler, D.F., Humphries, C.J., Eds.; Royal Botanic Gardens: Kew, UK, 1995; Volume 1, pp. 255–275. [Google Scholar]
- Cabrera, L.I.; Salazar, G.A.; Chase, M.W.; Mayo, S.J.; Bogner, J.; Dávila, P. Phylogenetic relationships of aroids and duckweeds (Araceae) inferred from coding and noncoding plastid DNA. Am. J. Bot. 2008, 95, 1153–1165. [Google Scholar] [CrossRef]
- Cusimano, N.; Bogner, J.; Mayo, S.J.; Boyce, P.C.; Wong, S.Y.; Hesse, M.; Hetterscheid, W.L.; Keating, R.C.; French, J.C. Relationships within the Araceae: Comparison of morphological patterns with molecular phylogenies. Am. J. Bot. 2011, 98, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Nauheimer, L.; Metzler, D.; Renner, S.S. Global history of the ancient monocot family Araceae inferred with models accounting for past continental positions and previous ranges based on fossils. New Phytol. 2012, 195, 938–950. [Google Scholar] [CrossRef]
- Martinov, I. Techno-Botanical Dictionary (Технo-Бoтанический Слoварь); Pechashano v Imperatorskoĭ Tipografii: Saint Petersburg, Russia, 1820. [Google Scholar]
- International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code); Wiersema, J.H.; Turland, N.J.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Association for Plant Taxonomy: Bratislava, Slovakia, 2018; Appendices I–VII. [Google Scholar]
- Bogner, J. The free-floating Aroids (Araceae)–living and fossil. Zitteliana A 2009, 48, 113–128. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group (APG). An ordinal classification for the families of flowering plants. Ann. Missouri Bot. Gard. 1998, 85, 531–553. [Google Scholar] [CrossRef] [Green Version]
- Landolt, E. The family of Lemnaceae—A monographic study. Ver. Geobot. Inst. ETH Stift. Rübel 1986, 71, 1–563. [Google Scholar]
- Les, D.H.; Landolt, E.; Crawford, D.J. Systematics of the Lemnaceae (duckweeds): Inferences from micromolecular and morphological data. Plant Syst. Evol. 1997, 204, 161–177. [Google Scholar] [CrossRef]
- Cusimano, N.; Sousa, A.; Renner, S.S. Maximum likelihood inference implies a high, not a low, ancestral haploid chromosome number in Araceae, with a critique of the bias introduced by ‘x’. Ann. Bot. 2012, 109, 681–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iles, W.J.D. The Phylogeny and Evolution of Two Ancient Lineages of Aquatic Plants. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2013. [Google Scholar]
- Tippery, N.P.; Les, D.H. Tiny plants with enormous potential: Phylogeny and evolution of duckweeds. In The Duckweed Genomes. Compendium of Plant Genomes; Cao, X., Fourounjian, P., Wang, W., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2020; pp. 19–38. [Google Scholar]
- Stevens, P.F. Angiosperm Phylogeny Website, 2001; Version 14. July 2017, Last Updated 24 February 2020. Available online: http://www.mobot.org/MOBOT/research/APweb (accessed on 8 April 2020).
- Les, D.H. Aquatic Monocotyledons of North America: Ecology, Life History, and Systematics; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Magallón, S.; Gómez-Acevedo, S.; Sánchez-Reyes, L.L.; Hernández-Hernández, T. A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 2015, 207, 437–453. [Google Scholar] [CrossRef]
- Jussieu, A.L. Genera Plantarum Secundum Ordines Naturales Disposita juxta Methodum in Horto Regio Parisiensi Exaratam, Anno 1774; Herissant: Paris, France, 1789. [Google Scholar]
- Brown, R. Prodromus Florae Novae Hollandiae; Johnson & Soc.: London, UK, 1810; Volume 1. [Google Scholar]
- Schott, H.W. Aroideae. Österr. Bot. Wochenbl. 1857, 7, 61–62. [Google Scholar] [CrossRef]
- Barton, W.P.C. Vegetable Materia Medica of the United States; H. C. Carey & I. Lea: Philadelphia, PA, USA, 1817. [Google Scholar]
- Schott, H.W. Genera Aroidearum; Carl Ueberreuter: Vienna, Austria, 1858. [Google Scholar]
- Engler, A. Das Pflanzenreich. Araceae–Pars Generalis et Index Familiae Generalis; Wilhelm Engelmann: Leipzig, Germany, 1920; Volume IV.23A. [Google Scholar]
- Hutchinson, J. The Families of Flowering Plants; Clarendon Press: London, UK, 1973. [Google Scholar]
- Grayum, M.H. Evolution and phylogeny of the Araceae. Ann. Missouri Bot. Gard. 1990, 77, 628–697. [Google Scholar] [CrossRef]
- Bartling, F.G. Ordines Naturales Plantarum; Sumptibus Dieterichianis: Göttingen, Germany, 1830. [Google Scholar]
- Grayum, M.H. A summary of evidence and arguments supporting the removal of Acorus from the Araceae. Taxon 1987, 36, 723–729. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, S.-H.; Lee, S.; Maki, M.; Otsuka, K.; Kozhevnikov, A.E.; Kozhevnikova, Z.V.; Wen, J.; Kim, S.-C. New insights into the phylogeny and biogeography of subfamily Orontioideae (Araceae). J. Syst. Evol. 2019, 57, 616–632. [Google Scholar] [CrossRef] [Green Version]
- Ray, T.S. Survey of shoot organization in the Araceae. Am. J. Bot. 1988, 75, 56–84. [Google Scholar] [CrossRef]
- Buzgo, M.; Endress, P.K. Floral structure and development of Acoraceae and its systematic relationships with basal angiosperms. Int. J. Plant Sci. 2000, 161, 23–41. [Google Scholar] [CrossRef]
- Williams, C.A.; Harborne, J.B.; Mayo, S.J. Anthocyanin pigments and leaf flavonoids in the family Araceae. Phytochemistry 1981, 20, 217–234. [Google Scholar] [CrossRef]
- Iwashina, T. Sebuah tinjauan: Senyawa-senyawa flavonoid pada suku Araceae. Bul. Kebun Raya 2020, 23, 1–24. [Google Scholar] [CrossRef]
- Tillich, H.-J. Seedling diversity in Araceae and its systematic implications. Feddes Repert. 2003, 114, 454–487. [Google Scholar] [CrossRef]
- Buzgo, M. Flower structure and development of Araceae compared with alismatids and Acoraceae. Bot. J. Linn. Soc. 2001, 136, 393–425. [Google Scholar] [CrossRef]
- Weber, M.; Halbritter, H.; Hesse, M. The basic pollen wall types in Araceae. Int. J. Plant Sci. 1999, 160, 415–423. [Google Scholar] [CrossRef]
- Hesse, M. Pollen wall ultrastructure of Araceae and Lemnaceae in relation to molecular classifications. Aliso 2006, 22, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Grayum, M.H. Systematic embryology of the Araceae. Bot. Rev. 1991, 57, 167–203. [Google Scholar] [CrossRef]
- Carlquist, S.; Schneider, E.L. Origins and nature of vessels in Monocotyledons. 14. Vessellessness in Orontioideae (Araceae): Adaptation or relictualism? Nord. J. Bot. 2014, 32, 493–502. [Google Scholar] [CrossRef]
- Mayo, S.J.; Bogner, J.; Boyce, P.C. Araceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 4, pp. 26–74. [Google Scholar]
- Nie, Z.L.; Sun, H.; Li, H.; Wen, J. Intercontinental biogeography of subfamily Orontioideae (Symplocarpus, Lysichiton, and Orontium) of Araceae in eastern Asia and North America. Mol. Phyl. Evol. 2006, 40, 155–165. [Google Scholar] [CrossRef]
- Bog, M.; Appenroth, K.J.; Sree, K.S. Duckweed (Lemnaceae): Its molecular taxonomy. Front. Sustain. Food Syst. 2019, 3, 117. [Google Scholar] [CrossRef]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef]
- Maheshwari, S.C. Spirodela polyrrhiza: The link between the aroids and the duckweeds. Nature 1958, 181, 1745–1746. [Google Scholar] [CrossRef]
- Sree, K.S.; Maheshwari, S.C.; Boka, K.; Khurana, J.P.; Keresztes, Á.; Appenroth, K.J. The duckweed Wolffia microscopica: A unique aquatic monocot. Flora 2015, 210, 31–39. [Google Scholar] [CrossRef]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 204. [Google Scholar] [CrossRef]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.J. Relative in vitro growth rates of duckweeds (Lemnaceae), the most rapidly growing higher plants. Plant Biol. 2015, 17 (Suppl. S1), 33–41. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, N.E.; Kelly, T.C.; Jansen, M.A. “Step by step”: High frequency short-distance epizoochorous dispersal of aquatic macrophytes. Biol. Invasions 2017, 19, 625–634. [Google Scholar] [CrossRef]
- Dumortier, B.C.J. Florula Belgica; J. Casterman: Tournai, Belgium, 1827. [Google Scholar]
- Schleiden, M.J. Prodromous monographiae Lemnacearum. Linnaea 1839, 13, 385–392. [Google Scholar]
- Hegelmaier, F. Die Lemnaceen. Eine Monographische Untersuchung; Wilhelm Engelmann: Leipzig, Germany, 1868. [Google Scholar]
- Grayum, M.H. Comparative external pollen ultrastructure of the Araceae and putatively related taxa. Monogr. Syst. Bot. Missouri Bot. Gard. 1992, 43, 1–167. [Google Scholar]
- Bog, M.; Appenroth, K.J.; Sree, K.S. Key to the determination of taxa within the family of Lemnaceae: An update. Nord. J. Bot. 2020, 38, e02658. [Google Scholar] [CrossRef]
- Bog, M.; Sree, K.S.; Fuchs, J.; Hoang, P.T.N.; Schubert, I.; Kuever, J.; Rabenstein, A.; Paolacci, S.; Jansen, M.A.K.; Appenroth, K.J. A taxonomic revision of Lemna sect. Uninerves (Lemnaceae). Taxon 2020, 69, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.F. A Natural Arrangement of British Plants; Baldwin, Cradock, and Joy: London, UK, 1821; Volume 2. [Google Scholar]
- Les, D.H.; Crawford, D.J.; Landolt, E.; Gabel, J.D.; Kimball, R.T. Phylogeny and systematics of Lemnaceae, the duckweed family. Syst. Bot. 2002, 27, 221–240. [Google Scholar] [CrossRef]
- Tippery, N.P.; Les, D.H.; Crawford, D.J. Evaluation of phylogenetic relationships in Lemnaceae using nuclear ribosomal data. Plant Biol. 2015, 17 (Suppl. S1), 50–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Haberer, G.; Gundlach, H.; Gläßer, C.; Nussbaumer, T.; Luo, M.-C.; Lomsadze, A.; Borodovsky, M.; Kerstetter, R.A.; Shanklin, J.; et al. The genome of the primordial monocotyledonous Spirodela polyrhiza: Neotenous reduction, fast growth, and aquatic lifestyle. Nat. Commun. 2014, 5, 3311. [Google Scholar] [CrossRef]
- Van Hoeck, A.; Horemans, N.; Monsieurs, P.; Cao, H.X.; Vandenhove, H.; Blust, R. The first draft genome of the aquatic model plant Lemna minor opens the route for future stress physiology research and biotechnological applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.X.; Vu, G.T.; Wang, W.; Appenroth, K.J.; Messing, J.; Schubert, I. The map-based genome sequence of Spirodela polyrhiza aligned with its chromosomes, a reference for karyotype evolution. New Phytol. 2016, 209, 354–363. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Li, C.S.; Zhou, Y.; Wu, Y.R.; Wang, W.Q. Genomes and transcriptomes of duckweeds. Front. Chem. 2018, 6, 230. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Michael, T.P.; Gilbert, S.; Chu, P.; Motley, S.T.; Appenroth, K.J.; Schubert, I.; Lam, E. Generating a high-confidence reference genome map of the Greater Duckweed by integration of cytogenomic, optical mapping, and Oxford Nanopore technologies. Plant J. 2018, 96, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Les, D.H.; Tippery, N.P. In time and with water… the systematics of alismatid monocotyledons. In Early Events in Monocot Evolution; Wilkin, P., Ed.; Cambridge University Press: Cambridge, UK, 2013; pp. 118–164. [Google Scholar]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberly, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Kubitzki, K.; Rudall, P.J.; Chase, M.C. Systematics and Evolution. In Flowering Plants Monocotyledons. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 3. [Google Scholar] [CrossRef]
- Eguchi, S.; Tamura, M.N. Evolutionary timescale of monocots determined by the fossilized birth-death model using a large number of fossil records. Evolution 2016, 70, 1136–1144. [Google Scholar] [CrossRef]
- Reveal, J.L.; Zomlefer, W.B. Two New Orders for Monocotyledonous Plants. Novon 1998, 8, 176–177. [Google Scholar] [CrossRef]
- Haynes, R.R.; Les, D.H. Alismatales (Water Plantains). In eLS; Wiley: Hoboken, NJ, USA, 2005. [Google Scholar] [CrossRef]
- Takhtajan, A. Diversity and Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1997. [Google Scholar]
- Cronquist, A. An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Tamura, M.N.; Yamashita, J.; Fuse, S.; Haraguchi, M. Molecular phylogeny of monocotyledons inferred from combined analysis of plastid matK and rbcL gene sequences. J. Plant. Res. 2004, 117, 109–120. [Google Scholar] [CrossRef]
- Janssen, T.; Bremer, K. The age of major monocot groups inferred from 800+rbcL sequences. Bot. J. Linn. Soc. 2004, 146, 385–398. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Messing, J. High-Throughput Sequencing of Three Lemnoideae (Duckweeds) Chloroplast Genomes from Total DNA. PLoS ONE 2011, 6, e24670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriquez, C.L.; Arias, T.; Pires, J.C.; Croat, T.B.; Schaal, B.A. Phylogenomics of the plant family Araceae. Mol. Phylogenet. Evol. 2014, 75, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Ruhfel, B.R.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E.; Burleigh, J.G. From algae to angiosperms–inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes. BMC Evol. Biol. 2014, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.T.; Yi, T.S.; Gao, L.M.; Ma, P.F.; Zhang, T.; Yang, J.B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef]
- Li, H.T.; Yi, T.S.; Gao, L.M.; Ma, P.F.; Zhang, T.; Yang, J.B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Data from: Origin of angiosperms and the puzzle of the Jurassic gap. Dryad Dataset 2019, 5, 461–470. [Google Scholar] [CrossRef]
- Ross, T.G.; Barrett, C.F.; Soto Gomez, M.; Lam, V.K.; Henriquez, C.L.; Les, D.H.; Davis, J.I.; Cuenca, A.; Petersen, G.; Seberg, O.; et al. Plastid phylogenomics and molecular evolution of Alismatales. Cladistics 2016, 32, 160–178. [Google Scholar] [CrossRef]
- Choi, K.S.; Park, K.T.; Park, S. The Chloroplast Genome of Symplocarpus renifolius: A Comparison of Chloroplast Genome Structure in Araceae. Genes 2017, 8, 324. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Yang, J.; Park, J.; Yamada, T.; Maki, M.; Kim, S.-C. Comparison of Whole Plastome Sequences between Thermogenic Skunk Cabbage Symplocarpus renifolius and Nonthermogenic S. nipponicus (Orontioideae; Araceae) in East Asia. Int. J. Mol. Sci. 2019, 20, 4678. [Google Scholar] [CrossRef] [Green Version]
- Henriquez, C.L.; Ahmed, I.; Carlsen, M.M.; Zuluaga, A.; Croat, T.B.; McKain, M.R. Evolutionary dynamics of chloroplast genomes in subfamily Aroideae (Araceae). Genomics 2020, 112, 2349–2360. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, C.L.; Mehmood, F.; Shahzadi, I.; Ali, Z.; Waheed, M.T.; Croat, T.B.; Poczai, P.; Ahmed, I. Comparison of Chloroplast Genomes among Species of Unisexual and Bisexual Clades of the Monocot Family Araceae. Plants 2020, 9, 737. [Google Scholar] [CrossRef]
- Henriquez, C.L.; Mehmood, F.; Carlsen, M.M.; Islam, M.; Waheed, M.T.; Poczai, P.; Croat, T.B.; Ahmed, I. Complete Chloroplast Genomes of Anthurium huixtlense and Pothos scandens (Pothoideae, Araceae): Unique Inverted Repeat Expansion and Contraction Affect Rate of Evolution. J. Mol. Evol. 2020, 88, 562–574. [Google Scholar] [CrossRef]
- Henriquez, C.L.; Croat, T.B.; Poczai, P.; Ahmed, I. Mutational Dynamics of Aroid Chloroplast Genomes II. Front. Genet. 2021, 11, 610838. [Google Scholar] [CrossRef]
- Sanderson, M.J. Estimating Absolute Rates of Molecular Evolution and Divergence Times: A Penalized Likelihood Approach. Mol. Biol. Evol. 2002, 19, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS Region of Nuclear Ribosomal DNA: A Valuable Source of Evidence on Angiosperm Phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Soltis, D.E.; Soltis, P.S.; Nickrent, D.L.; Johnson, L.A.; Hahn, W.J.; Hoot, S.B.; Sweere, J.A.; Kuzoff, R.K.; Kron, K.A.; Chase, M.W.; et al. Angiosperm Phylogeny Inferred from 18S Ribosomal DNA Sequences. Ann. Mo. Bot. Gard. 1997, 84, 1–49. [Google Scholar] [CrossRef]
- Maia, V.H.; Gitzendanner, M.A.; Soltis, P.S.; Wong, G.K.S.; Soltis, D.E. Angiosperm Phylogeny Based on 18S/26S rDNA Sequence Data: Constructing a Large Data Set Using Next-Generation Sequence Data. Int. J. Plant Sci. 2014, 175, 613–650. [Google Scholar] [CrossRef]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenet. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef] [Green Version]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Blattner, F.R. Direct Amplification of the Entire ITS Region from Poorly Preserved Plant Material Using Recombinant PCR. BioTechniques 1999, 27, 1180–1186. [Google Scholar] [CrossRef]
- Shoup, S.; Lewis, L.A. Polyphyletic origin of parallel basal bodies in swimming cells of chlorophycean green algae (Chlorophyta). J. Phycol. 2003, 39, 789–796. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, C.M.; Neiman, M.; Taylor, D.R. Inheritance and recombination of mitochondrial genomes in plants, fungi and animals. New Phytol. 2005, 168, 39–50. [Google Scholar] [CrossRef]
- Pentinsaari, M.; Salmela, H.; Mutanen, M.; Roslin, T. Molecular evolution of a widely-adopted taxonomic marker (COI) across the animal tree of life. Sci. Rep. 2016, 6, 35275. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Palmer, J.D. Multiple acquisitions via horizontal transfer of a group I intron in the mitochondrial cox1 gene during evolution of the Araceae family. Mol. Biol. Evol. 1999, 16, 1155–1165. [Google Scholar] [CrossRef] [Green Version]
- Cusimano, N.; Zhang, L.B.; Renner, S.S. Reevaluation of the cox1 Group I Intron in Araceae and Angiosperms Indicates a History Dominated by Loss rather than Horizontal Transfer. Mol. Biol. Evol. 2008, 25, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Li, L.; Hendry, T.A.; Li, R.; Taylor, D.W.; Issa, M.J.; Ronen, A.J.; Vekaria, M.L.; White, A.M. Reconstructing the basal angiosperm phylogeny: Evaluating information content of mitochondrial genes. Taxon 2006, 55, 837–856. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.L.; Li, L.; Wang, B.; Xue, J.Y.; Hendry, T.A.; Li, R.Q.; Brown, J.W.; Liu, Y.; Hudson, G.T.; Chen, Z.D. Angiosperm phylogeny inferred from sequences of four mitochondrial genes. J. Syst. Evol. 2010, 48, 391–425. [Google Scholar] [CrossRef]
- Petersen, G.; Seberg, O.; Davis, J.I.; Stevenson, D.W. RNA editing and phylogenetic reconstruction in two monocot mitochondrial genes. Taxon 2006, 55, 871–886. [Google Scholar] [CrossRef]
- Cuenca, A.; Petersen, G.; Seberg, O.; Davis, J.I.; Stevenson, D.W. Are substitution rates and RNA editing correlated? BMC Evol. Biol. 2010, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Petersen, G.; Cuenca, A.; Zervas, A.; Ross, G.T.; Graham, S.W.; Barrett, C.F.; Davis, J.I.; Seberg, O. Mitochondrial genome evolution in Alismatales: Size reduction and extensive loss of ribosomal protein genes. PLoS ONE 2017, 12, e0177606. [Google Scholar] [CrossRef] [Green Version]
- Mower, J.P. Variation in protein gene and intron content among land plant mitogenomes. Mitochondrion 2020, 53, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, Y.; Messing, J. The Mitochondrial Genome of an Aquatic Plant, Spirodela polyrhiza. PLoS ONE 2012, 7, e46747. [Google Scholar] [CrossRef] [Green Version]
- Thompson, S.A. Araceae. In Flora of North America Editorial Committee. Flora of North America, North of Mexico. Magnoliophyta: Alismatidae, Arecidae, Commelinidae (in Part), and Zingiberidae; Oxford University Press: New York, NY, USA, 2000; Volume 22. [Google Scholar]
- Haynes, R.R. Hydrocharitaceae. In Flora of North America Editorial Committee. Flora of North America, North of Mexico. Magnoliophyta: Alismatidae, Arecidae, Commelinidae (in Part), and Zingiberidae; Oxford University Press: New York, NY, USA, 2000; Volume 22. [Google Scholar]
- Keating, R.C. Acoraceae and Araceae. In Anatomy of the Monocotyledons; Gregory, M., Cutler, D.F., Eds.; Oxford University Press: Oxford, UK, 2002; Volume 9. [Google Scholar]
- Sender, L.M.; Doyle, J.A.; Upchurch, G.R., Jr.; Villanueva-Amadoz, U.; Diez, J.B. Leaf and inflorescence evidence for near-basal Araceae and an unexpected diversity of other monocots from the late Early Cretaceous of Spain. J. Syst. Palaeontol. 2019, 17, 1313–1346. [Google Scholar] [CrossRef]
- Stockey, R.A.; Hoffman, G.L.; Rothwell, G.W. Fossil evidence for Paleocene diversification of Araceae: Bognerospadix gen. nov. and Orontiophyllum grandifolium comb. nov. Am. J. Bot. 2021, 108, 1417–1440. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 3234. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Schubert, I. Reconstruction of chromosome rearrangements between the two most ancestral duckweed species Spirodela polyrhiza and S. intermedia. Chromosoma 2017, 126, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Jiang, L.; Wang, L.; Han, X.; Guo, W.; Li, C.; Zhou, Y.; Denton, M.; Zhang, P. A high-quality genome of taro (Colocasia esculenta (L.) Schott), one of the world’s oldest crops. Mol. Ecol. Resour. 2021, 21, 68–77. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org (accessed on 15 October 2021).
- Brummitt, R.K. World Geographical Scheme for Recording Plant Distributions; For International Working Group on Taxonomic Databases for Plant Sciences (TDWG) 153; Hunt Institute for Botanical Documentation; Carnegie Mellon University: Pittsburgh, PA, USA, 2001. [Google Scholar]
- Smith, S.A.; Brown, J.W. Constructing a broadly inclusive seed plant phylogeny. Am. J. Bot. 2018, 105, 302–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmstead, R.G.; de Pamphilis, C.W.; Wolfe, A.D.; Young, N.D.; Elisons, W.J.; Reeves, P.A. Disintegration of the Scrophulariaceae. Am. J. Bot. 2001, 88, 348–361. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tippery, N.P.; Les, D.H.; Appenroth, K.J.; Sree, K.S.; Crawford, D.J.; Bog, M. Lemnaceae and Orontiaceae Are Phylogenetically and Morphologically Distinct from Araceae. Plants 2021, 10, 2639. https://doi.org/10.3390/plants10122639
Tippery NP, Les DH, Appenroth KJ, Sree KS, Crawford DJ, Bog M. Lemnaceae and Orontiaceae Are Phylogenetically and Morphologically Distinct from Araceae. Plants. 2021; 10(12):2639. https://doi.org/10.3390/plants10122639
Chicago/Turabian StyleTippery, Nicholas P., Donald H. Les, Klaus J. Appenroth, K. Sowjanya Sree, Daniel J. Crawford, and Manuela Bog. 2021. "Lemnaceae and Orontiaceae Are Phylogenetically and Morphologically Distinct from Araceae" Plants 10, no. 12: 2639. https://doi.org/10.3390/plants10122639