
Chlorophyll Fluorescence Imaging-Based Duckweed Phenotyping to Assess Acute Phytotoxic Effects


Abstract
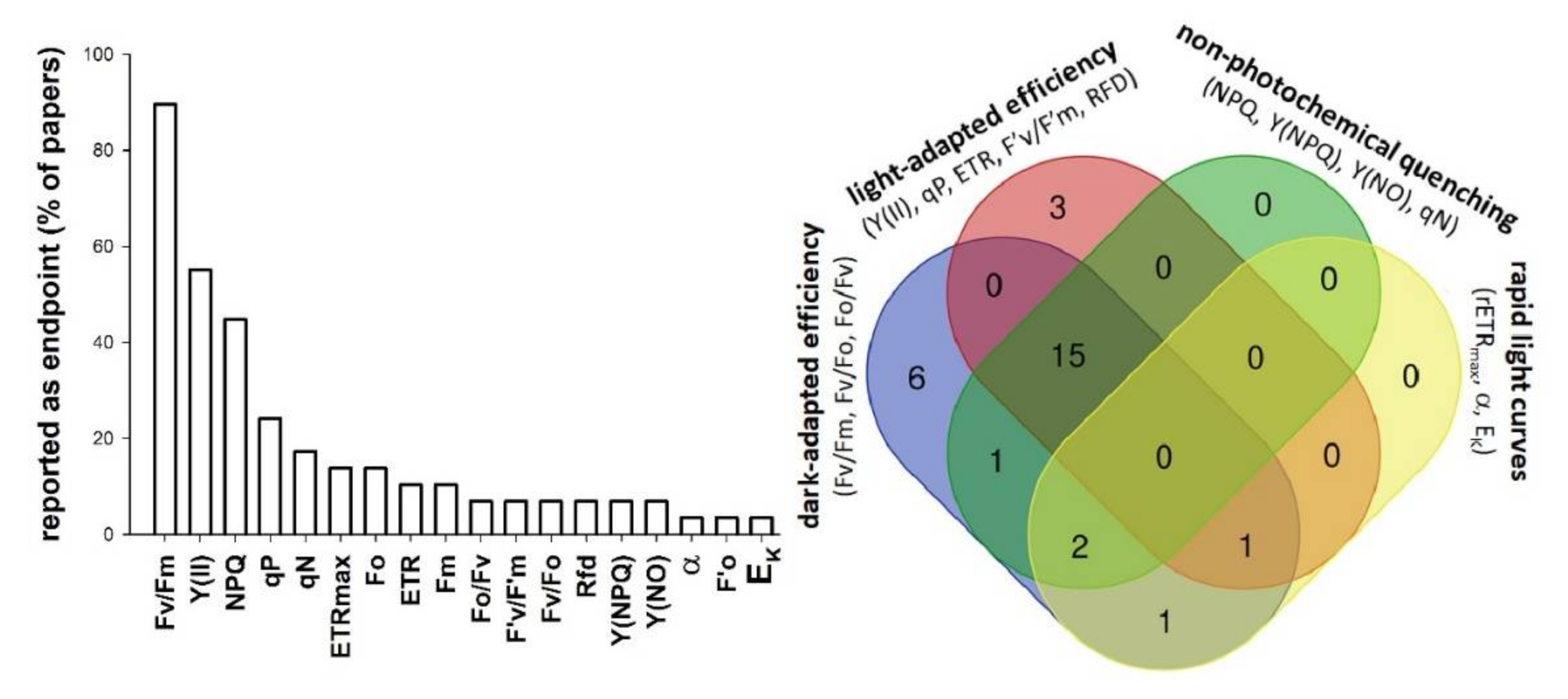
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Chlorophyll Fluorescence Imaging
- First, the test plants in plate were dark-adapted for 20 min in order to reach a fully oxidized state of PSII photochemistry (all PSII reaction centers were “open”).
- In dark-adapted state ground fluorescence yield (Fo) of plants was determined using weak, non-inductive measuring light (blue, 450 nm, intensity: 2, frequency: 1).
- A single saturating pulse (blue, 450 nm, intensity: 9, i.e., ~4000 µE m−2 s−1 with the applied instrumental setup, length 720 ms) was used to saturate PSII photochemistry (all PSII reaction centers became “closed”) and to determine maximal fluorescence yield (Fm) of plants.
- After the saturation pulse, a continuous actinic irradiation (blue, 450 nm) with similar intensity (intensity: 5, equivalent to ~77 µE m−2 s−1 in the applied instrumental setup) to that of the plants’ ambient light environment was switched on for 10 min to induce a light-adapted state of plants (Supplementary Figure S1). After 10 min, steady-state chlorophyll fluorescence yield (Fs) of plants under actinic illumination was determined and a second saturating pulse—with the same settings as for determining Fm—was applied to measure their light-adapted maximal fluorescence yield (F’m). Due to technical reasons, F’o, which is the ground fluorescence yield in a light-acclimated state, was not directly measured by the instrument, but calculated using the formula [30]:
2.3. Data Processing and Statistical Analyses
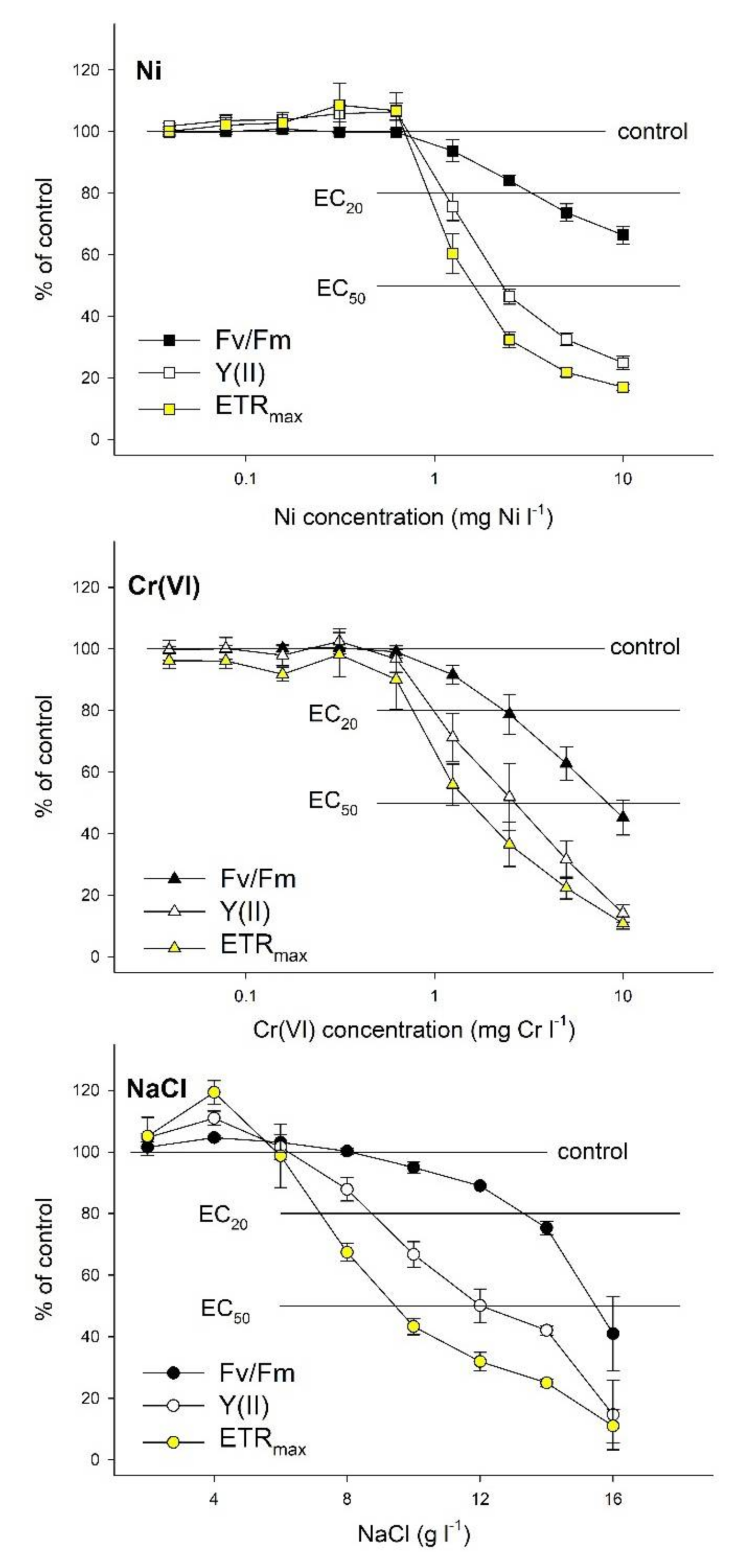
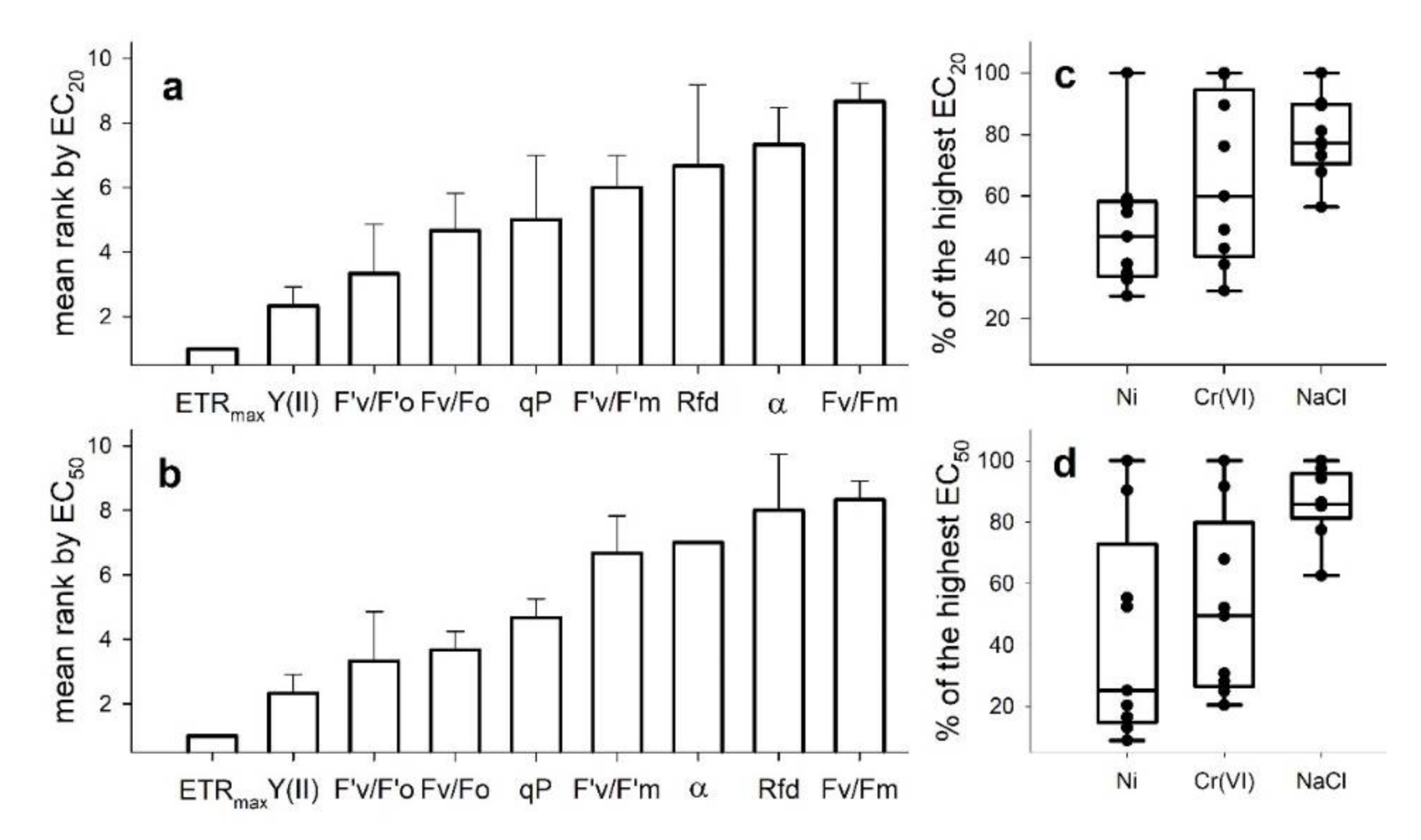
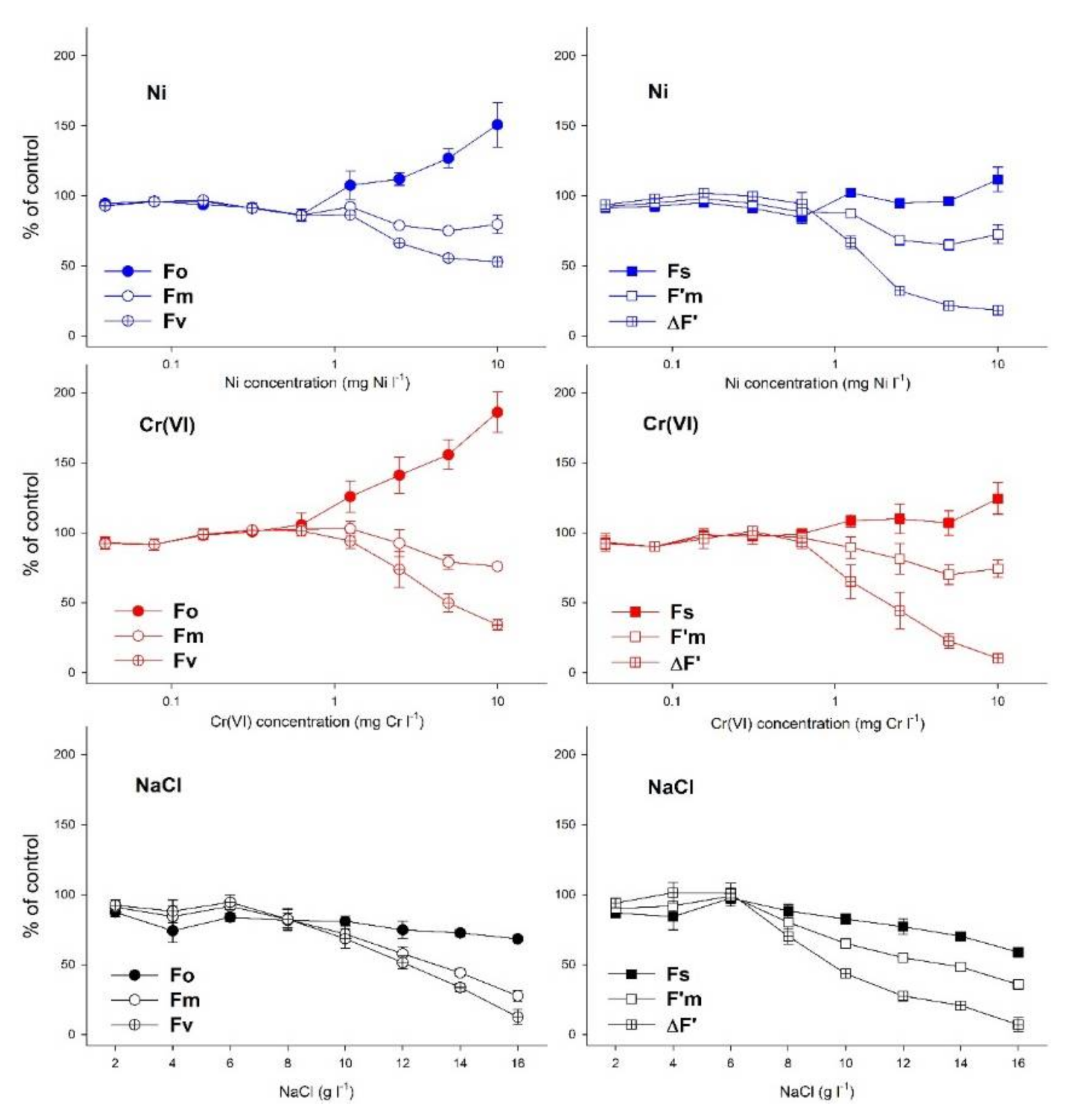
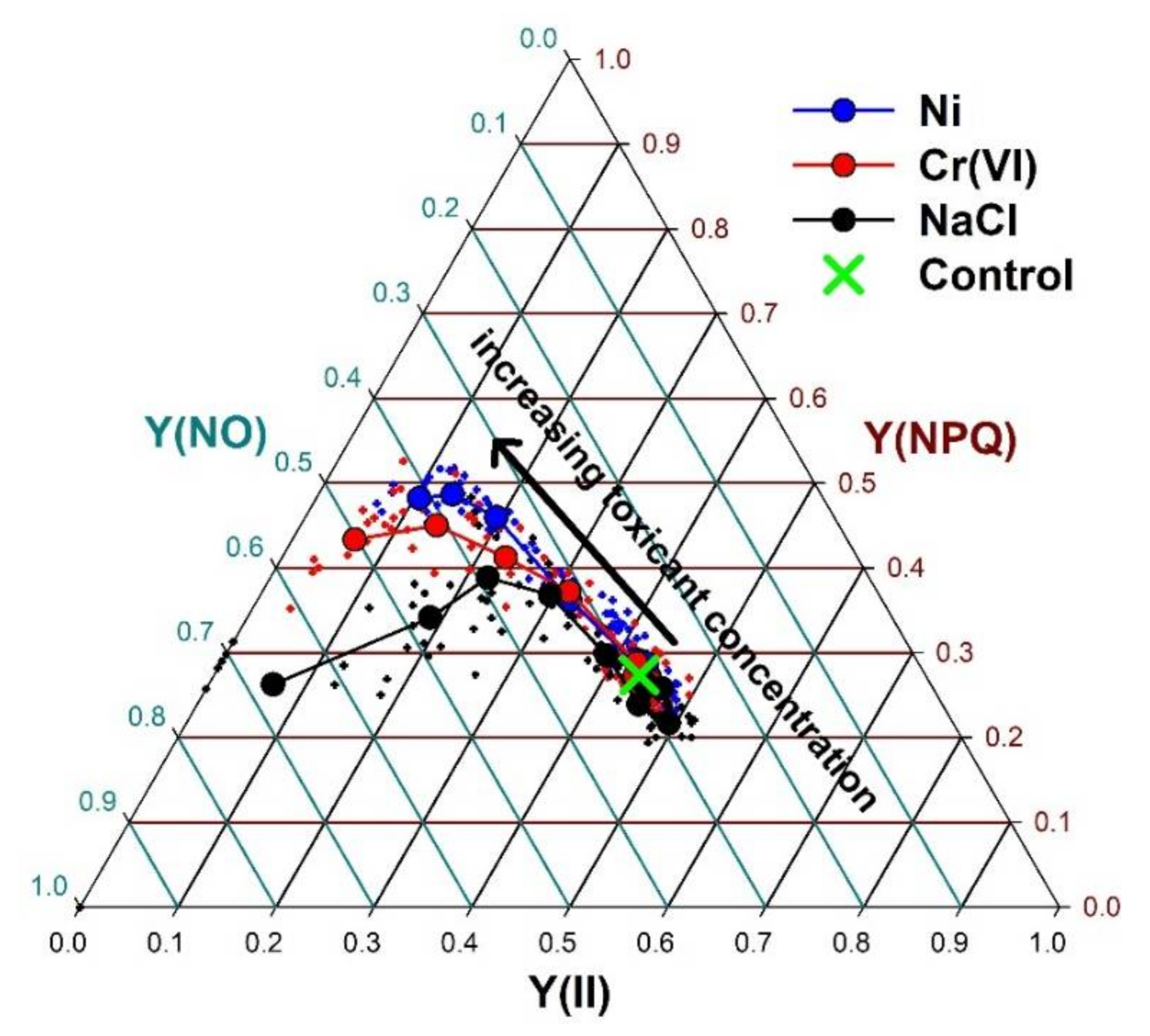
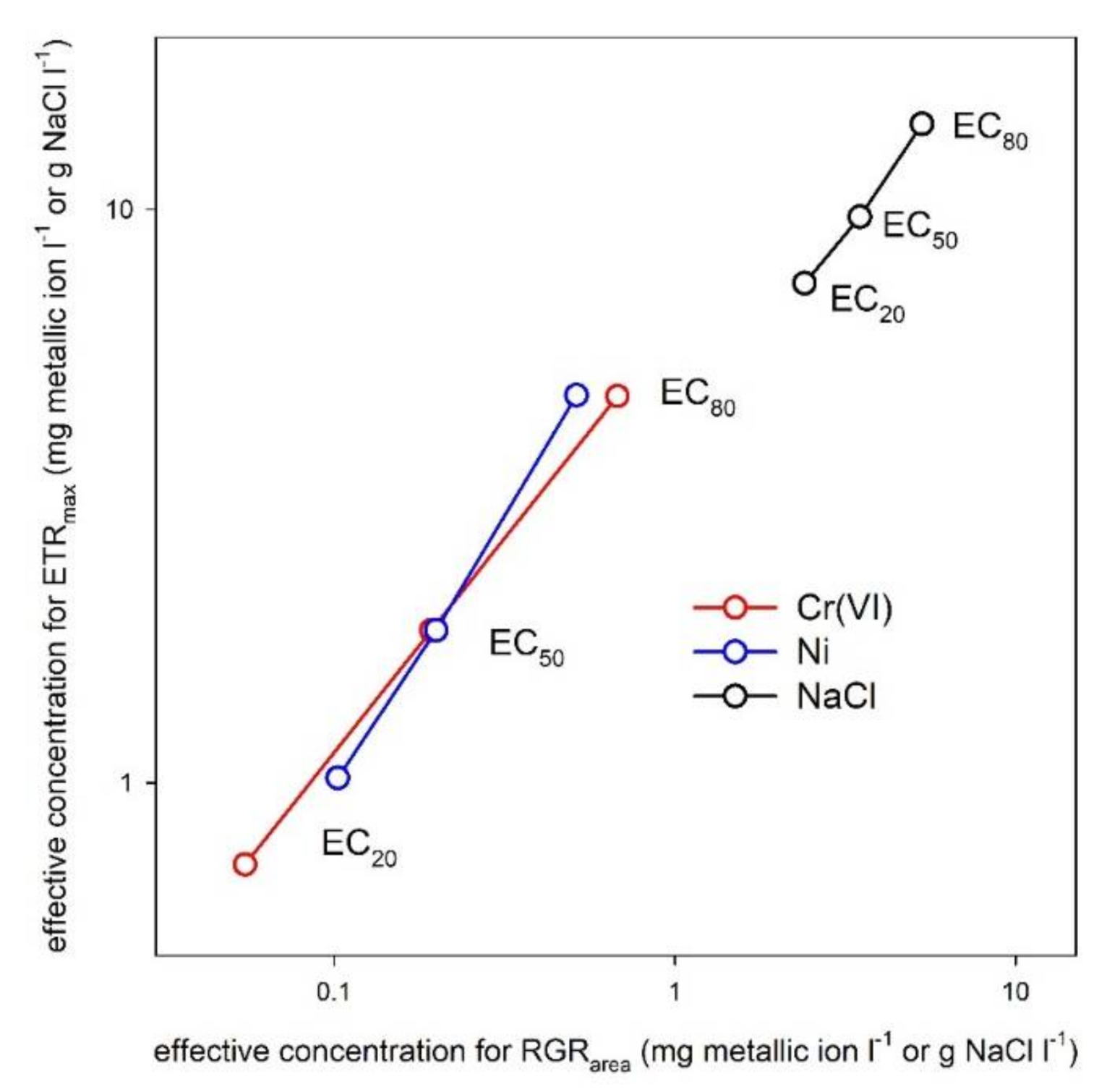
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Environment Canada Biological Test Method-Test for Measuring the Inhibition of Growth Using the Freshwater Macrophyte Lemna minor, 2nd ed.; Environmental Protection Service Report EPS 1/RM/37; Method Development and Applications Section, Environmental Technology Centre, Environment Canada: Ottawa, ON, Canada, 2007; ISBN 978-0-662-43340-8.
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a Model Plant System in the Genomics and Postgenomics Era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Relative in Vitro Growth Rates of Duckweeds (Lemnaceae)-the Most Rapidly Growing Higher Plants. Plant Biol. J. 2015, 17, 33–41. [Google Scholar] [CrossRef] [PubMed]
- ISO 20079:2005. Water Quality-Determination of Toxic Effect of Water Constituents and Waste Water to Duckweed (Lemna minor)—Duckweed Growth Inhibition Test, 1st ed.; International Organization for Standardization: Geneva, Switzerland, 2005. [Google Scholar]
- OECD Guidelines for the Testing of Chemicals, Revised Proposal for a New Guideline 221, Lemna Sp. Growth Inhibition Test; OECD: Paris, France, 2006.
- Mkandawire, M.; Teixeira da Silva, J.A.; Dudel, E.G. The Lemna Bioassay: Contemporary Issues as the Most Standardized Plant Bioassay for Aquatic Ecotoxicology. Crit. Rev. Environ. Sci. Technol. 2014, 44, 154–197. [Google Scholar] [CrossRef]
- Henke, R.; Eberius, M.; Appenroth, K.-J. Induction of Frond Abscission by Metals and Other Toxic Compounds in Lemna minor. Aquat. Toxicol. 2011, 101, 261–265. [Google Scholar] [CrossRef]
- Park, A.; Kim, Y.-J.; Choi, E.-M.; Brown, M.T.; Han, T. A Novel Bioassay Using Root Re-Growth in Lemna. Aquat. Toxicol. 2013, 140–141, 415–424. [Google Scholar] [CrossRef]
- Brain, R.A.; Cedergreen, N. Biomarkers in Aquatic Plants: Selection and Utility. In Reviews of Environmental Contamination and Toxicology; Springer: New York, NY, USA, 2009; Volume 198, pp. 1–61. ISBN 978-0-387-09646-9. [Google Scholar]
- Ralph, P.J.; Smith, R.A.; Macinnis-Ng, C.M.O.; Seery, C.R. Use of Fluorescence-Based Ecotoxicological Bioassays in Monitoring Toxicants and Pollution in Aquatic Systems: Review. Toxicol. Environ. Chem. 2007, 89, 589–607. [Google Scholar] [CrossRef]
- Ziegler, P.; Sree, K.S.; Appenroth, K.-J. Duckweed Biomarkers for Identifying Toxic Water Contaminants? Environ. Sci Pollut. Res. 2019, 26, 14797–14822. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Quayle, P.; Schmidt, S.; Escher, B.I.; Mueller, J.F. Methodology and Evaluation of a Highly Sensitive Algae Toxicity Test Based on Multiwell Chlorophyll Fluorescence Imaging. Biosens. Bioelectron. 2007, 22, 2554–2563. [Google Scholar] [CrossRef]
- Muller, R.; Schreiber, U.; Escher, B.I.; Quayle, P.; Bengtson Nash, S.M.; Mueller, J.F. Rapid Exposure Assessment of PSII Herbicides in Surface Water Using a Novel Chlorophyll a Fluorescence Imaging Assay. Sci. Total Environ. 2008, 401, 51–59. [Google Scholar] [CrossRef]
- Ralph, P.J.; Macinnis-Ng, C.M.O.; Frankart, C. Fluorescence Imaging Application: Effect of Leaf Age on Seagrass Photokinetics. Aquat. Bot. 2005, 81, 69–84. [Google Scholar] [CrossRef]
- Wilkinson, A.D.; Collier, C.J.; Flores, F.; Mercurio, P.; O’Brien, J.; Ralph, P.J.; Negri, A.P. A Miniature Bioassay for Testing the Acute Phytotoxicity of Photosystem II Herbicides on Seagrass. PLoS ONE 2015, 10, e0117541. [Google Scholar] [CrossRef]
- Chen, Y.-E.; Wu, N.; Zhang, Z.-W.; Yuan, M.; Yuan, S. Perspective of Monitoring Heavy Metals by Moss Visible Chlorophyll Fluorescence Parameters. Front. Plant Sci. 2019, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- DalCorso, G. Heavy Metal Toxicity in Plants. In Plants and Heavy Metals; Furini, A., Ed.; Springer Briefs in Molecular Science; Springer: Dordrecht, The Netherlands, 2012; pp. 1–25. ISBN 978-94-007-4440-0. [Google Scholar]
- Appenroth, K.-J.; Krech, K.; Keresztes, Á.; Fischer, W.; Koloczek, H. Effects of Nickel on the Chloroplasts of the Duckweeds Spirodela Polyrhiza and Lemna minor and Their Possible Use in Biomonitoring and Phytoremediation. Chemosphere 2010, 78, 216–223. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Hayat, S.; Ahmad, A. Nickel: An Overview of Uptake, Essentiality and Toxicity in Plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17. [Google Scholar] [CrossRef]
- Andresen, E.; Opitz, J.; Thomas, G.; Stärk, H.-J.; Dienemann, H.; Jenemann, K.; Dickinson, B.C.; Küpper, H. Effects of Cd & Ni Toxicity to Ceratophyllum Demersum under Environmentally Relevant Conditions in Soft & Hard Water Including a German Lake. Aquat. Toxicol. 2013, 142–143, 387–402. [Google Scholar] [CrossRef]
- Naumann, B.; Eberius, M.; Appenroth, K.-J. Growth Rate Based Dose–Response Relationships and EC-Values of Ten Heavy Metals Using the Duckweed Growth Inhibition Test (ISO 20079) with Lemna minor L. Clone St. J. Plant Physiol. 2007, 164, 1656–1664. [Google Scholar] [CrossRef]
- Oláh, V.; Hepp, A.; Mészáros, I. Comparative Study on the Sensitivity of Turions and Active Fronds of Giant Duckweed (Spirodela Polyrhiza (L.) Schleiden) to Heavy Metal Treatments. Chemosphere 2015, 132, 40–46. [Google Scholar] [CrossRef]
- Cervantes, C.; Loza-Tavera, H.; Torres-Guzman, J.C.; Moreno-Sanchez, R. Interactions of Chromium with Microorganisms and Plants. FEMS Microbiol. Rev. 2001, 13, 335–347. [Google Scholar] [CrossRef]
- Shanker, A.; Cervantes, C.; Lozatavera, H.; Avudainayagam, S. Chromium Toxicity in Plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant Response to Salt Stress and Role of Exogenous Protectants to Mitigate Salt-Induced Damages. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 25–87. ISBN 978-1-4614-4746-7. [Google Scholar]
- Jajoo, A. Changes in Photosystem II in Response to Salt Stress. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 149–168. ISBN 978-1-4614-4746-7. [Google Scholar]
- Sree, K.S.; Adelmann, K.; Garcia, C.; Lam, E.; Appenroth, K.-J. Natural Variance in Salt Tolerance and Induction of Starch Accumulation in Duckweeds. Planta 2015, 241, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Oláh, V.; Hepp, A.; Gaibor Vaca, N.Y.; Tamás, M.; Mészáros, I. Retrospective Analyses of Archive Phytotoxicity Test Data Can Help in Assessing Internal Dynamics and Stability of Growth in Laboratory Duckweed Cultures. Aquat. Toxicol. 2018, 201, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Walz GmbH IMAGING-PAM M-Series Chlorophyll Fluorometer: Instrument Description and Information for Users, 5th ed.; Heinz Walz GmbH: Effeltrich, Germany, 2019.
- Roháček, K. Chlorophyll Fluorescence Parameters: The Definitions, Photosynthetic Meaning, and Mutual Relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II Quantum Yields Calculated from Simple Fluorescence Parameters Measured by PAM Fluorometry and the Saturation Pulse Method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to Correctly Determine the Different Chlorophyll Fluorescence Parameters and the Chlorophyll Fluorescence Decrease Ratio RFd of Leaves with the PAM Fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Park, J.; Brown, M.T.; Depuydt, S.; Kim, J.K.; Won, D.-S.; Han, T. Comparing the Acute Sensitivity of Growth and Photosynthetic Endpoints in Three Lemna Species Exposed to Four Herbicides. Environ. Pollut. 2017, 220, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently Asked Questions about Chlorophyll Fluorescence, the Sequel. Photosynth. Res. 2017, 132, 13–66. [Google Scholar] [CrossRef] [Green Version]
- Ritz, C.; Streibig, J.C. Bioassay Analysis Using R. J. Stat. Softw. 2005, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Brain, P.; Cousens, R. An Equation to Describe Dose Responses Where There Is Stimulation of Growth at Low Doses. Weed Res. 1989, 29, 93–96. [Google Scholar] [CrossRef]
- Cedergreen, N.; Ritz, C.; Streibig, J.C. Improved Empirical Models Describing Hormesis. Environ. Toxicol Chem 2005, 24, 3166. [Google Scholar] [CrossRef]
- Babani, F.; Lichtenthaler, H.K. Light-Induced and Age-Dependent Development of Chloroplasts in Etiolated Barley Leaves as Visualized by Determination of Photosynthetic Pigments, C02 Assimilation Rates and Different Kinds of Chlorophyll Fluorescence Ratios. J. Plant Physiol. 1996, 148, 555–566. [Google Scholar] [CrossRef]
- Láposi, R.; Veres, S.; Lakatos, G.; Oláh, V.; Fieldsend, A.; Mészáros, I. Responses of Leaf Traits of European Beech (Fagus sylvatica L.) Saplings to Supplemental UV-B Radiation and UV-B Exclusion. Agric. For. Meteorol. 2009, 149, 745–755. [Google Scholar] [CrossRef]
- Oláh, V.; Lakatos, G.; Bertók, C.; Kanalas, P.; Szőllősi, E.; Kis, J.; Mészáros, I. Short-Term Chromium(VI) Stress Induces Different Photosynthetic Responses in Two Duckweed Species, Lemna gibba L. and Lemna minor L. Photosynthetica 2010, 48, 513–520. [Google Scholar] [CrossRef]
- Pietrini, F.; Bianconi, D.; Massacci, A.; Iannelli, M.A. Combined Effects of Elevated CO2 and Cd-Contaminated Water on Growth, Photosynthetic Response, Cd Accumulation and Thiolic Components Status in Lemna minor L. J. Hazard. Mater. 2016, 309, 77–86. [Google Scholar] [CrossRef]
- Kumar, K.S.; Han, T. Physiological Response of Lemna Species to Herbicides and Its Probable Use in Toxicity Testing. Toxicol. Environ. Health Sci. 2010, 2, 39–49. [Google Scholar] [CrossRef]
- Perreault, F.; Oukarroum, A.; Pirastru, L.; Sirois, L.; Gerson Matias, W.; Popovic, R. Evaluation of Copper Oxide Nanoparticles Toxicity Using Chlorophyll a Fluorescence Imaging in Lemna gibba. J. Bot. 2010, 2010, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lahive, E. Frond Development Gradients Are a Determinant of the Impact of Zinc on Photosynthesis in Three Species of Lemnaceae. Aquat. Bot. 2012, 101, 55–63. [Google Scholar] [CrossRef]
- Flores, F.; Collier, C.J.; Mercurio, P.; Negri, A.P. Phytotoxicity of Four Photosystem II Herbicides to Tropical Seagrasses. PLoS ONE 2013, 8, e75798. [Google Scholar] [CrossRef] [Green Version]
- Dewez, D.; Goltsev, V.; Kalaji, H.M.; Oukarroum, A. Inhibitory Effects of Silver Nanoparticles on Photosystem II Performance in Lemna gibba Probed by Chlorophyll Fluorescence. Curr. Plant Biol. 2018, 16, 15–21. [Google Scholar] [CrossRef]
- Pietrini, F.; Passatore, L.; Fischetti, E.; Carloni, S.; Ferrario, C.; Polesello, S.; Zacchini, M. Evaluation of Morpho-Physiological Traits and Contaminant Accumulation Ability in Lemna minor L. Treated with Increasing Perfluorooctanoic Acid (PFOA) Concentrations under Laboratory Conditions. Sci. Total Environ. 2019, 695, 133828. [Google Scholar] [CrossRef]
- Park, J.; Lee, H.; Han, T. Comparative Paraquat Sensitivity of Newly Germinated and Mature Fronds of the Aquatic Macrophyte Spirodela Polyrhiza. AJPS 2020, 11, 1008–1024. [Google Scholar] [CrossRef]
- Lee, H.; Depuydt, S.; Shin, K.; Choi, S.; Kim, G.; Lee, Y.H.; Park, J.T.; Han, T.; Park, J. Assessment of Various Toxicity Endpoints in Duckweed (Lemna minor) at the Physiological, Biochemical, and Molecular Levels as a Measure of Diuron Stress. Biology 2021, 10, 684. [Google Scholar] [CrossRef]
- Park, J.-S.; Brown, M.T.; Han, T. Phenol Toxicity to the Aquatic Macrophyte Lemna Paucicostata. Aquat. Toxicol. 2012, 106–107, 182–188. [Google Scholar] [CrossRef]
- Kummerová, M.; Zezulka, Š.; Babula, P.; Tříska, J. Possible Ecological Risk of Two Pharmaceuticals Diclofenac and Paracetamol Demonstrated on a Model Plant Lemna minor. J. Hazard. Mat. 2016, 302, 351–361. [Google Scholar] [CrossRef]
- Hepp, A.; Oláh, V.; Sipos, O.; Adorján, B.; Mészáros, I. Békalencse fitotoxikológiai tesztek eredményét befolyásoló tényezők vizsgálata (Assessment of factors affecting the results of duckweed phytotoxicity tests). In Proceedings of the XIV Environmental Scientific Conference of the Carpathian Basin, Gödöllő, Hungary, 5–7 April 2018; pp. 127–131. [Google Scholar]
- Oláh, V.; Hepp, A.; Lakatos, G.; Mészáros, I. Cadmium-Induced Turion Formation of Spirodela Polyrhiza (L.) Schleiden. Acta Biol. Szeged. 2014; 58, 103–108. [Google Scholar]
- Alkimin, G.D.; Daniel, D.; Frankenbach, S.; Serôdio, J.; Soares, A.M.V.M.; Barata, C.; Nunes, B. Evaluation of Pharmaceutical Toxic Effects of Non-Standard Endpoints on the Macrophyte Species Lemna minor and Lemna gibba. Sci. Total Environ. 2019, 657, 926–937. [Google Scholar] [CrossRef]
- Christensen, M.G.; Teicher, H.B.; Streibig, J.C. Linking Fluorescence Induction Curve and Biomass in Herbicide Screening. Pest. Manag. Sci. 2003, 59, 1303–1310. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Blain, R.B. Hormesis and Plant Biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef]
- Agathokleous, E. Environmental Hormesis, a Fundamental Non-Monotonic Biological Phenomenon with Implications in Ecotoxicology and Environmental Safety. Ecotoxicol. Environ. Saf. 2018, 148, 1042–1053. [Google Scholar] [CrossRef] [Green Version]
- Cedergreen, N.; Streibig, J.C.; Kudsk, P.; Mathiassen, S.K.; Duke, S.O. The Occurrence of Hormesis in Plants and Algae. Dose-Response 2007, 5, 150–162. [Google Scholar] [CrossRef]
- Belz, R.G.; Cedergreen, N.; Sørensen, H. Hormesis in Mixtures—Can It Be Predicted? Sci. Total Environ. 2008, 404, 77–87. [Google Scholar] [CrossRef]
- Agathokleous, E.; Mouzaki-Paxinou, A.-C.; Saitanis, C.J.; Paoletti, E.; Manning, W.J. The First Toxicological Study of the Antiozonant and Research Tool Ethylene Diurea (EDU) Using a Lemna minor L. Bioassay: Hints to Its Mode of Action. Environ. Pollut. 2016, 213, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Li, Y.; Cheng, J.J. Effects of Selenite on Chlorophyll Fluorescence, Starch Content and Fatty Acid in the Duckweed Landoltia Punctata. J. Plant Res. 2016, 129, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E. The Rise and Fall of Photosynthesis: Hormetic Dose Response in Plants. J. For. Res. 2021, 32, 889–898. [Google Scholar] [CrossRef]
- Hulsen, K.; Minne, V.; Lootens, P.; Vandecasteele, P.; Hofte, M. A Chlorophyll a Fluorescence-Based Lemna minor Bioassay to Monitor Microbial Degradation of Nanomolar to Micromolar Concentrations of Linuron. Environ. Microbiol. 2002, 4, 327–337. [Google Scholar] [CrossRef]
- Küster, A.; Pohl, K.; Altenburger, R. A Fluorescence-Based Bioassay for Aquatic Macrophytes and Its Suitability for Effect Analysis of Non-Photosystem II Inhibitors. Environ. Sci. Pollut. Res. Int. 2007, 14, 377–383. [Google Scholar] [CrossRef]
- Juhel, G.; Batisse, E.; Hugues, Q.; Daly, D.; van Pelt, F.N.A.M.; O’Halloran, J.; Jansen, M.A.K. Alumina Nanoparticles Enhance Growth of Lemna minor. Aquat. Toxicol. 2011, 105, 328–336. [Google Scholar] [CrossRef]
- Lahive, E.; O’ Halloran, J.; Jansen, M.A.K. Differential Sensitivity of Four Lemnaceae Species to Zinc Sulphate. Environ. Exp. Bot. 2011, 71, 25–33. [Google Scholar] [CrossRef]
- Zezulka, Š.; Kummerová, M.; Babula, P.; Váňová, L. Lemna minor Exposed to Fluoranthene: Growth, Biochemical, Physiological and Histochemical Changes. Aquat. Toxicol. 2013, 140–141, 37–47. [Google Scholar] [CrossRef]
- Senavirathna, M.D.H.J.; Takashi, A.; Kimura, Y. Short-Duration Exposure to Radiofrequency Electromagnetic Radiation Alters the Chlorophyll Fluorescence of Duckweeds (Lemna minor). Electromagn. Biol. Med. 2014, 33, 327–334. [Google Scholar] [CrossRef]
- Gomes, M.P.; Juneau, P. Oxidative Stress in Duckweed (Lemna minor L.) Induced by Glyphosate: Is the Mitochondrial Electron Transport Chain a Target of This Herbicide? Environ. Pollut. 2016, 218, 402–409. [Google Scholar] [CrossRef]
- Di Baccio, D.; Pietrini, F.; Bertolotto, P.; Pérez, S.; Barcelò, D.; Zacchini, M.; Donati, E. Response of Lemna gibba L. to High and Environmentally Relevant Concentrations of Ibuprofen: Removal, Metabolism and Morpho-Physiological Traits for Biomonitoring of Emerging Contaminants. Sci. Total Environ. 2017, 584–585, 363–373. [Google Scholar] [CrossRef]
- Paolacci, S. The Invasive Duckweed Lemna Minuta Kunth Displays a Different Light Utilisation Strategy than Native Lemna minor Linnaeus. Aquat. Bot. 2018, 7, 8–14. [Google Scholar] [CrossRef]
- Chen, X.; O’Halloran, J.; Jansen, M.A.K. Orthophosphate Modulates the Phytotoxicity of Nano-ZnO to Lemna minor (L.). Environ. Technol. 2019, 40, 2446–2454. [Google Scholar] [CrossRef]
- Grenni, P.; Patrolecco, L.; Rauseo, J.; Spataro, F.; Di Lenola, M.; Aimola, G.; Zacchini, M.; Pietrini, F.; Di Baccio, D.; Stanton, I.C.; et al. Sulfamethoxazole Persistence in a River Water Ecosystem and Its Effects on the Natural Microbial Community and Lemna minor Plant. Microchem. J. 2019, 149, 103999. [Google Scholar] [CrossRef]
- Mateos-Cárdenas, A.; Scott, D.T.; Seitmaganbetova, G.; Frank, N.A.M.v.P.; John, O.; Marcel, A.K.J. Polyethylene Microplastics Adhere to Lemna minor (L.), yet Have No Effects on Plant Growth or Feeding by Gammarus duebeni (Lillj.). Sci. Total Environ. 2019, 689, 413–421. [Google Scholar] [CrossRef]
- Nunes, B.; Veiga, V.; Frankenbach, S.; Serôdio, J.; Pinto, G. Evaluation of Physiological Changes Induced by the Fluoroquinolone Antibiotic Ciprofloxacin in the Freshwater Macrophyte Species Lemna minor and Lemna gibba. Environ. Toxicol. Pharmacol. 2019, 72, 103242. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, H.; Wang, Y.; Tang, X.; He, G.; Wang, S.; Ma, Y.; Kong, Y.; Yu, C.; Zhou, G. A Submerged Duckweed Mutant with Abundant Starch Accumulation for Bioethanol Production. GCB Bioenergy 2020, 12, 1078–1091. [Google Scholar] [CrossRef]
- Shi, H.; Duan, M.; Li, C.; Zhang, Q.; Liu, C.; Liang, S.; Guan, Y.; Kang, X.; Zhao, Z.; Xiao, G. The Change of Accumulation of Heavy Metal Drive Interspecific Facilitation under Copper and Cold Stress. Aquat. Toxicol. 2020, 225, 105550. [Google Scholar] [CrossRef]
- Walsh, É.; Kuehnhold, H.; O’Brien, S.; Coughlan, N.E.; Jansen, M.A.K. Light Intensity Alters the Phytoremediation Potential of Lemna minor. Environ. Sci. Pollut. Res. 2021, 28, 16394–16407. [Google Scholar] [CrossRef]
- Walsh, É.; Coughlan, N.E.; O’Brien, S.; Jansen, M.A.K.; Kuehnhold, H. Density Dependence Influences the Efficacy of Wastewater Remediation by Lemna minor. Plants 2021, 10, 1366. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Calculation | Description | Reference |
|---|---|---|---|
| Fv/Fm | (Fm − Fo)/Fm | Maximum quantum yield of PSII photochemistry | [31] |
| Fv/Fo | (Fm − Fo)/Fo | Maximum ratio of quantum yields of photochemical and concurrent non-photochemical processes in PSII in dark-adapted state | [31] |
| Y(II) | ∆F/F’m | The effective quantum yield of photochemical energy conversion in PSII | [31] |
| Y(NPQ) | (Fs/F’m) − (Fs/Fm) | Quantum yield of regulated non-photochemical energy loss in PSII | [32] |
| Y(NO) | Fs/Fm | Quantum yield of non-regulated heat dissipation and fluorescence emission | [32] |
| qP | 1 − ((Fs − F’o)/F’v) = ∆F/F’v = (F’m − Fs)/(F’m − F’o) | Photochemical quenching of variable ChlF, i.e., the fraction of open PSII reaction centers in light-adapted state | [31] |
| F’v/F’m | (F’m − F’o)/F’m | Effective quantum yield of PSII photochemistry in light-adapted state | [31] |
| F’v/F’o | (F’m − F’o)/F’o | Effective ratio of quantum yields of photochemical and concurrent non-photochemical processes in PSII related to light-adapted state | [31] |
| Rfd | (Fm − Fs)/Fs | Chlorophyll fluorescence decrease ratio proportional to net photosynthesis; a.k.a. “vitality index” | [33] |
| Parameter | Model | DF | RSS | F-Value | Pseudo-R2 | Max | ECmax | NOAEC (95% CI) | EC20 (95% CI) | EC50 (95% CI) |
|---|---|---|---|---|---|---|---|---|---|---|
| Fv/Fm | LL.3 | 27 | 323.56 | 1.7206 | 0.9286 | NA | NA | NA | 3.72 (2.87–4.57) | 18.6 * (13.8–23.4) |
| Fv/Fo | BC.4 | 26 | 914.79 | 0.5439 | 0.9522 | 102.5 | 0.20 | 0.52 (0.16–0.88) | 1.41 (1.10–1.72) | 4.20 (3.36–5.03) |
| Y(II) | BC.4 | 26 | 679 | 3.6103 | 0.9775 | 108.3 | 0.26 | 0.67 (0.49–0.84) | 1.22 (1.05–1.38) | 2.68 (2.35–3.01) |
| qP | CRS.4c | 26 | 1652 | 1.0886 | 0.9151 | 105.8 | 0.13 | 0.61 (0.09–1.14) | 1.74 (1.14–2.35) | 5.19 (3.81–6.56) |
| F’v/F’m | BC.4 | 26 | 404.32 | 1.4081 | 0.9600 | 102.6 | 0.21 | 0.61 (0.30–0.91) | 2.03 (1.65–2.41) | 10.8 * (7.55–14.0) |
| F’v/F’o | BC.4 | 26 | 1051.19 | 1.3835 | 0.9554 | 105.6 | 0.23 | 0.64 (0.38–0.89) | 1.3 (1.04–1.56) | 3.40 (2.74–4.07) |
| Rfd | BC.4 | 26 | 302.72 | 3.8371 | 0.9602 | 103.4 | 0.13 | 0.38 (0.05–0.71) | 2.13 (1.69–2.57) | 20.59 * (12.0–29.2) |
| α | BC.4 | 26 | 747.14 | 0.3612 | 0.9267 | 103.3 | 0.23 | 0.68 (0.25–1.11) | 2.20 (1.66–2.75) | 11.4 * (6.70–16.1) |
| ETRmax | BC.4 | 26 | 1671.24 | 2.5494 | 0.9600 | 110.0 | 0.28 | 0.66 (0.47–0.85) | 1.02 (0.85–1.19) | 1.84 (1.56–2.13) |
| Parameter | Model | DF | RSS | F-Value | Pseudo-R2 | Max | ECmax | NOAEC (95% CI) | EC20 (95% CI) | EC50 (95% CI) |
|---|---|---|---|---|---|---|---|---|---|---|
| Fv/Fm | BC.4 | 26 | 727.5 | 0.0317 | 0.9336 | 100.5 | 0.18 | 0.45 (−0.58–1.47) | 2.46 (1.89–3.03) | 8.26 (6.25–10.3) |
| Fv/Fo | BC.4 | 26 | 1783.9 | 0.0571 | 0.9382 | 102.1 | 0.21 | 0.49 (−0.02–1.00) | 1.21 (0.89–1.53) | 2.77 (2.24–3.30) |
| Y(II) | CRS.4b | 26 | 1873.8 | 0.2508 | 0.9395 | 102.2 | 0.24 | 0.4 (−0.03–0.83) | 1.06 (0.72–1.40) | 2.54 (2.00–3.09) |
| qP | LL.3 | 27 | 5191.1 | 0.3166 | 0.8252 | NA | NA | NA | 2.21 (1.13–3.23) | 4.47 (3.26–5.68) |
| F’v/F’m | BC.4 | 26 | 984.35 | 0.066 | 0.9426 | 100.3 | 0.13 | 0.32 (−0.36–1.00) | 1.48 (1.09–1.87) | 4.70 (3.74–5.67) |
| F’v/F’o | BC.4 | 26 | 2127.9 | 0.1227 | 0.9307 | 100.9 | 0.14 | 0.33 (−0.16–0.82) | 0.93 (0.63–1.23) | 2.25 (1.75–2.74) |
| Rfd | BC.4 | 26 | 1537.2 | 0.2505 | 0.8784 | 105.3 | 0.34 | 0.98 (0.37–1.58) | 2.47 (1.77–3.17) | 9.02 (5.12–12.9) |
| α | LL.3 | 27 | 918.75 | 0.2719 | 0.9357 | NA | NA | NA | 1.88 (1.34–2.43) | 6.13 (5.11–7.15) |
| ETRmax | LL.3 | 27 | 2303.1 | 1.0996 | 0.9344 | NA | NA | NA | 0.72 (0.47–0.98) | 1.85 (1.45–2.24) |
| Parameter | Model | Df | Rss | F-Value | Pseudo-R2 | Max | ECmax | NOAEC (95% CI) | EC20 (95% CI) | EC50 (95% CI) |
|---|---|---|---|---|---|---|---|---|---|---|
| Fv/Fm | LL.3 | 24 | 1109.59 | 0.5767 | 0.8996 | NA | NA | NA | 13.2 (12.4–14.0) | 15.5 (15.0–15.9) |
| Fv/Fo | CRS.4a | 23 | 1273.78 | 2.8564 | 0.9554 | 113.2 | 4.55 | 8.78 (7.47–10.1) | 10.7 (9.96–11.5) | 13.2 (12.6–13.8) |
| Y(II) | CRS.4a | 23 | 1771.4 | 1.3314 | 0.9381 | 108.2 | 3.20 | 6.48 (4.34–8.61) | 8.95 (7.87–10.0) | 12.0 (11.1–12.8) |
| qP | LL.3 | 24 | 2319.2 | 1.4362 | 0.8816 | NA | NA | NA | 9.65 (8.13–11.2) | 13.3 (12.3–14.3) |
| F’v/F’m | LL.3 | 24 | 1546.45 | 1.983 | 0.8844 | NA | NA | NA | 11.8 (10.7–13.0) | 15.1 (14.4–15.9) |
| F’v/F’o | BC.4 | 23 | 2031 | 3.5422 | 0.9333 | 124.6 | 4.63 | 8.57 (7.52–9.61) | 10.2 (9.37–11.1) | 13.3 (12.2–14.3) |
| Rfd | LL.3 | 24 | 1206.4 | 1.4771 | 0.9374 | NA | NA | NA | 10.1 (9.09–11.2) | 13.4 (12.8–14.0) |
| α | LL.3 | 24 | 1479 | 0.8038 | 0.9053 | NA | NA | NA | 11.9 (10.9–12.9) | 14.6 (14.1–15.2) |
| ETRmax | BC.4 | 23 | 1632.4 | 0.9605 | 0.9595 | 115.8 | 3.45 | 6.06 (5.12–7.00) | 7.44 (6.76–8.11) | 9.71 (9.06–10.4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oláh, V.; Hepp, A.; Irfan, M.; Mészáros, I. Chlorophyll Fluorescence Imaging-Based Duckweed Phenotyping to Assess Acute Phytotoxic Effects. Plants 2021, 10, 2763. https://doi.org/10.3390/plants10122763
Oláh V, Hepp A, Irfan M, Mészáros I. Chlorophyll Fluorescence Imaging-Based Duckweed Phenotyping to Assess Acute Phytotoxic Effects. Plants. 2021; 10(12):2763. https://doi.org/10.3390/plants10122763
Chicago/Turabian StyleOláh, Viktor, Anna Hepp, Muhammad Irfan, and Ilona Mészáros. 2021. "Chlorophyll Fluorescence Imaging-Based Duckweed Phenotyping to Assess Acute Phytotoxic Effects" Plants 10, no. 12: 2763. https://doi.org/10.3390/plants10122763