
New Insights into Interspecific Hybridization in Lemna L. Sect. Lemna (Lemnaceae Martinov)

 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. TBP Profiling of Duckweed Clones in Lemna Sect. Lemna
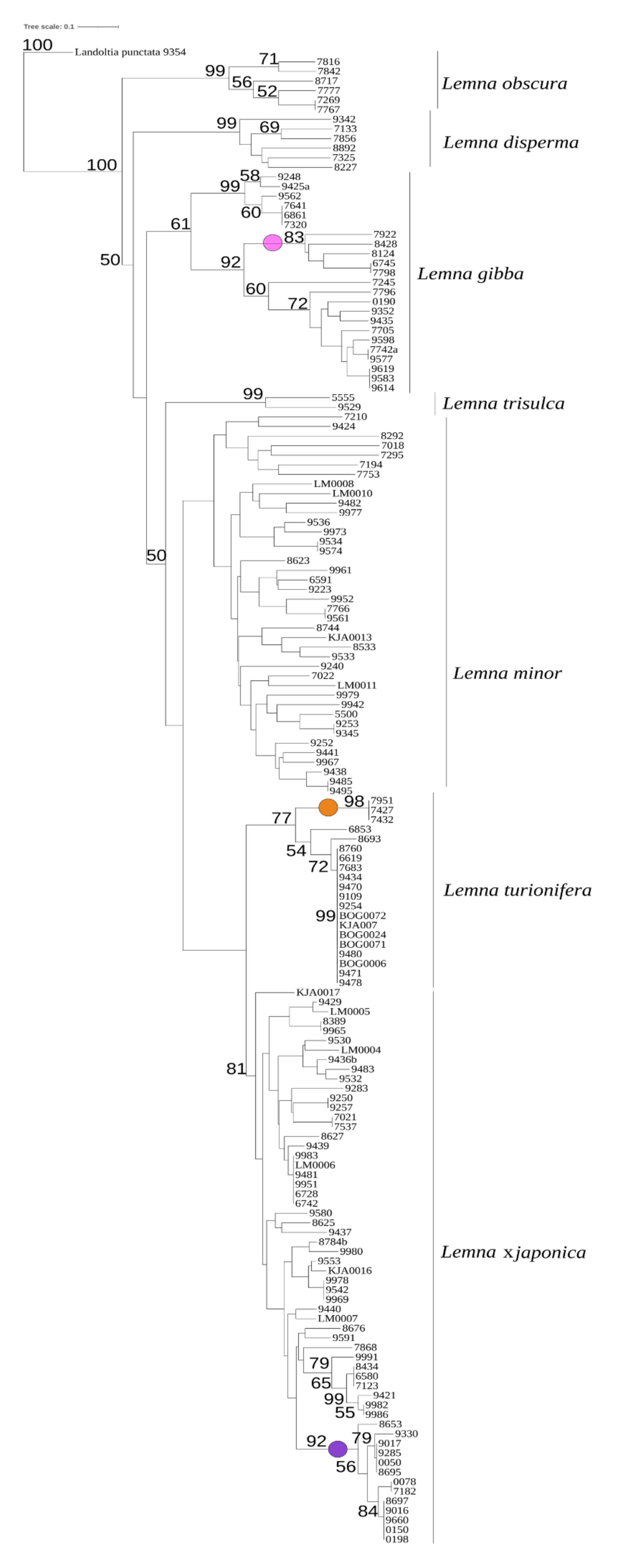
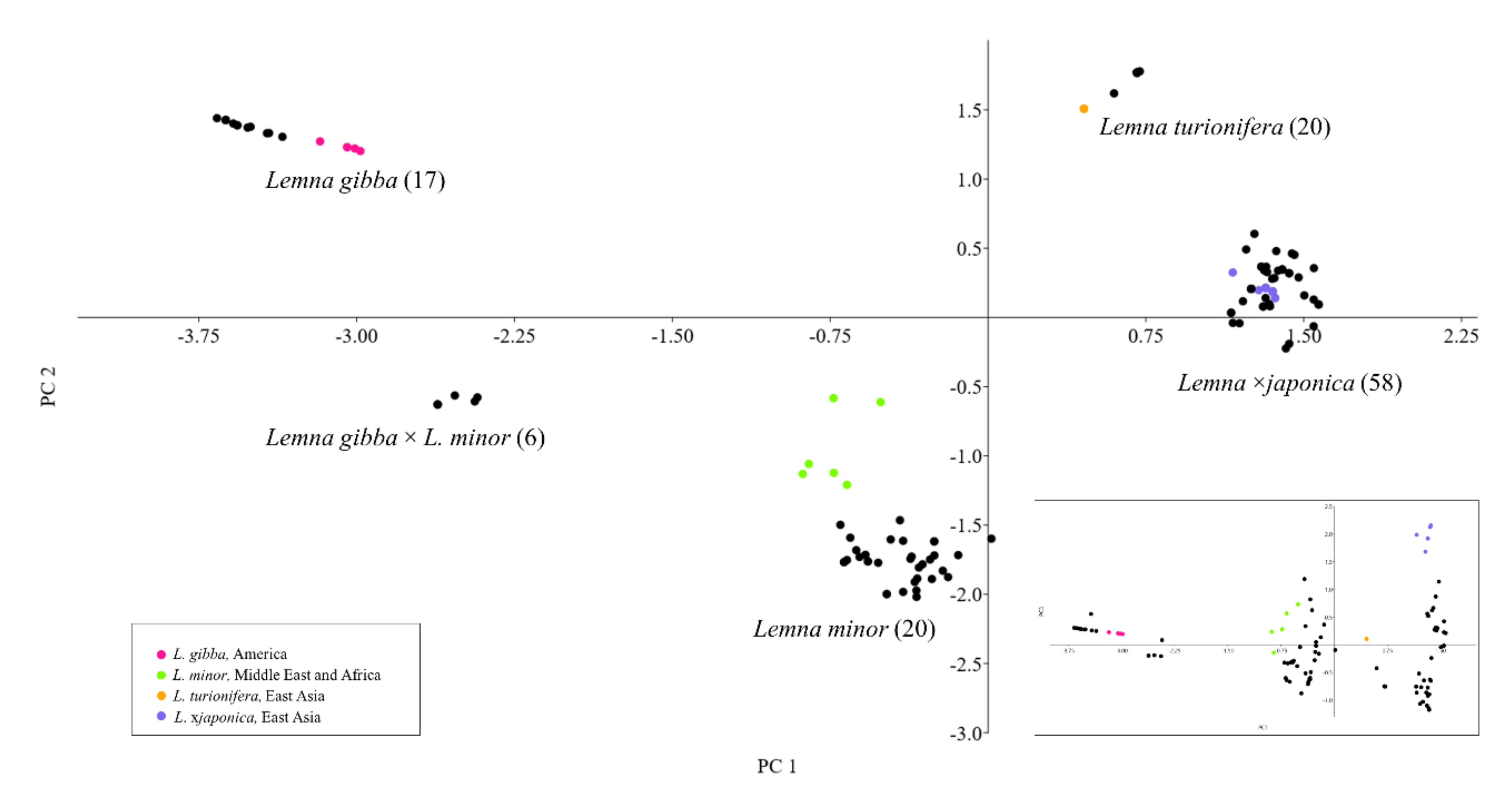
2.2. Reclassification of Clones by TBP
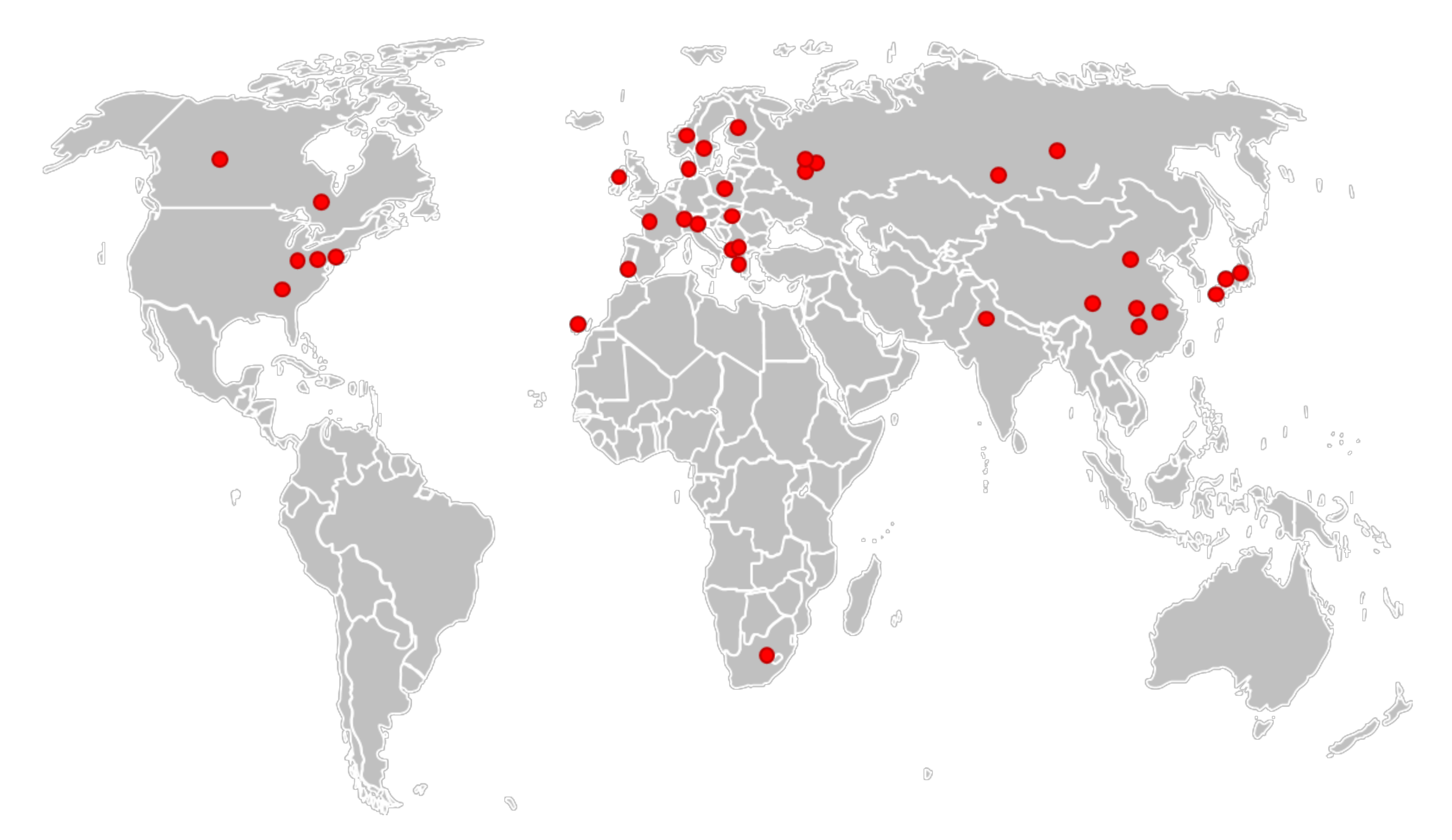
- The abundance of L. ×japonica populations was highly underestimated, as well as its geographical distribution, which is not limited to Japan, Korea and the east coast of China, as reported by Landolt for L. japonica [16]. The actual distribution, deduced from investigated clones and shown in Figure 3, covers all the temperate regions from Eastern Asia to Central Asia, Europe and North America, although their invasive origin in the different regions remains to be elucidated. One clone was even found in South Africa (Figure 3).
- At least part of the huge variability observed in L. minor, e.g., in genome size, ploidy or physiological parameters etc. could be due to erroneous classification of clones.
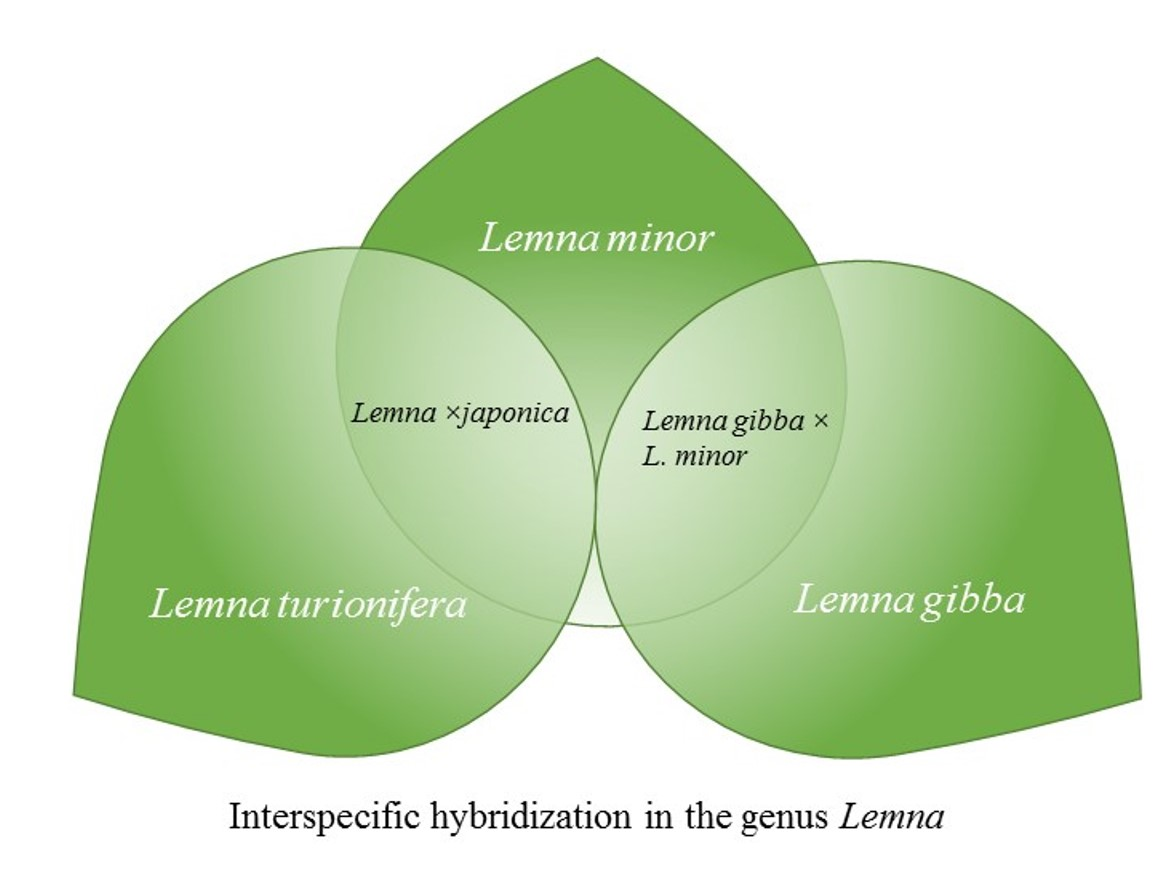
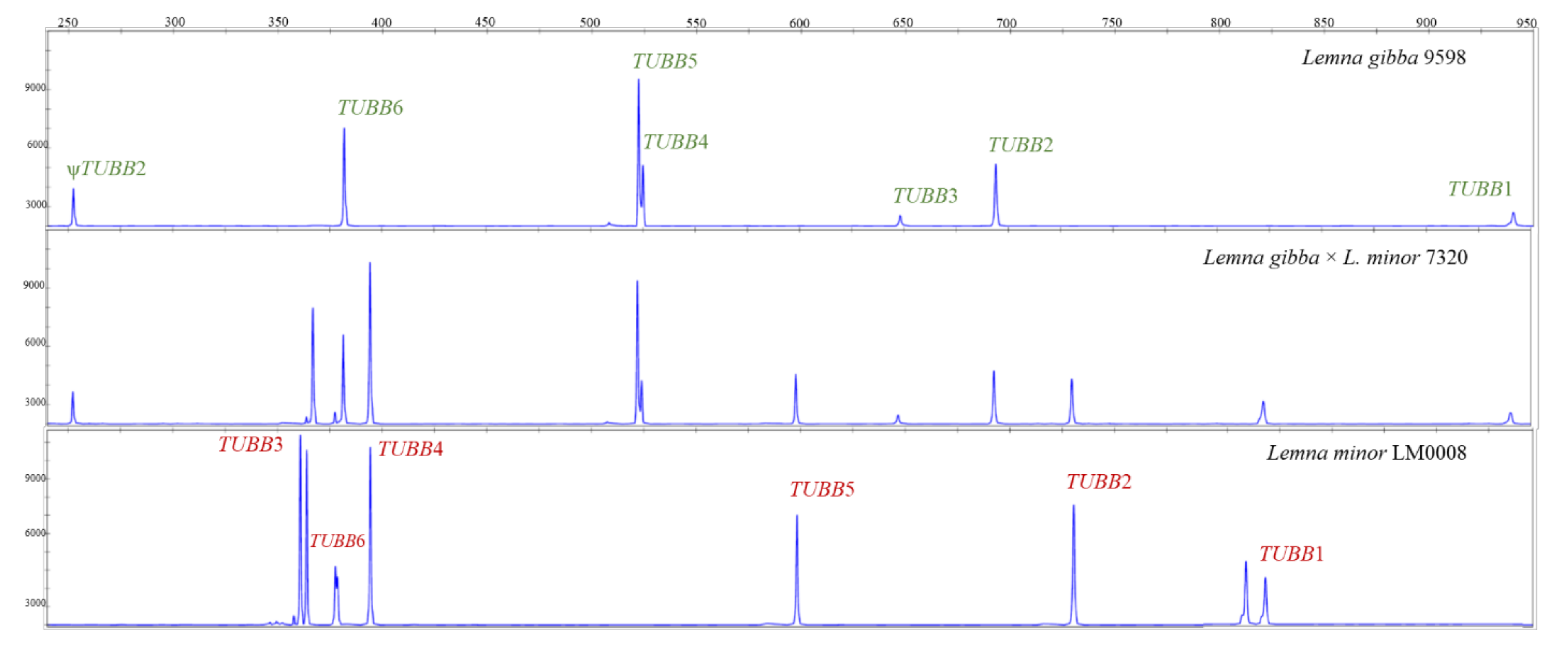
2.3. Lemna gibba × Lemna Minor Hybrids
2.4. Frond Morphology and Flowering
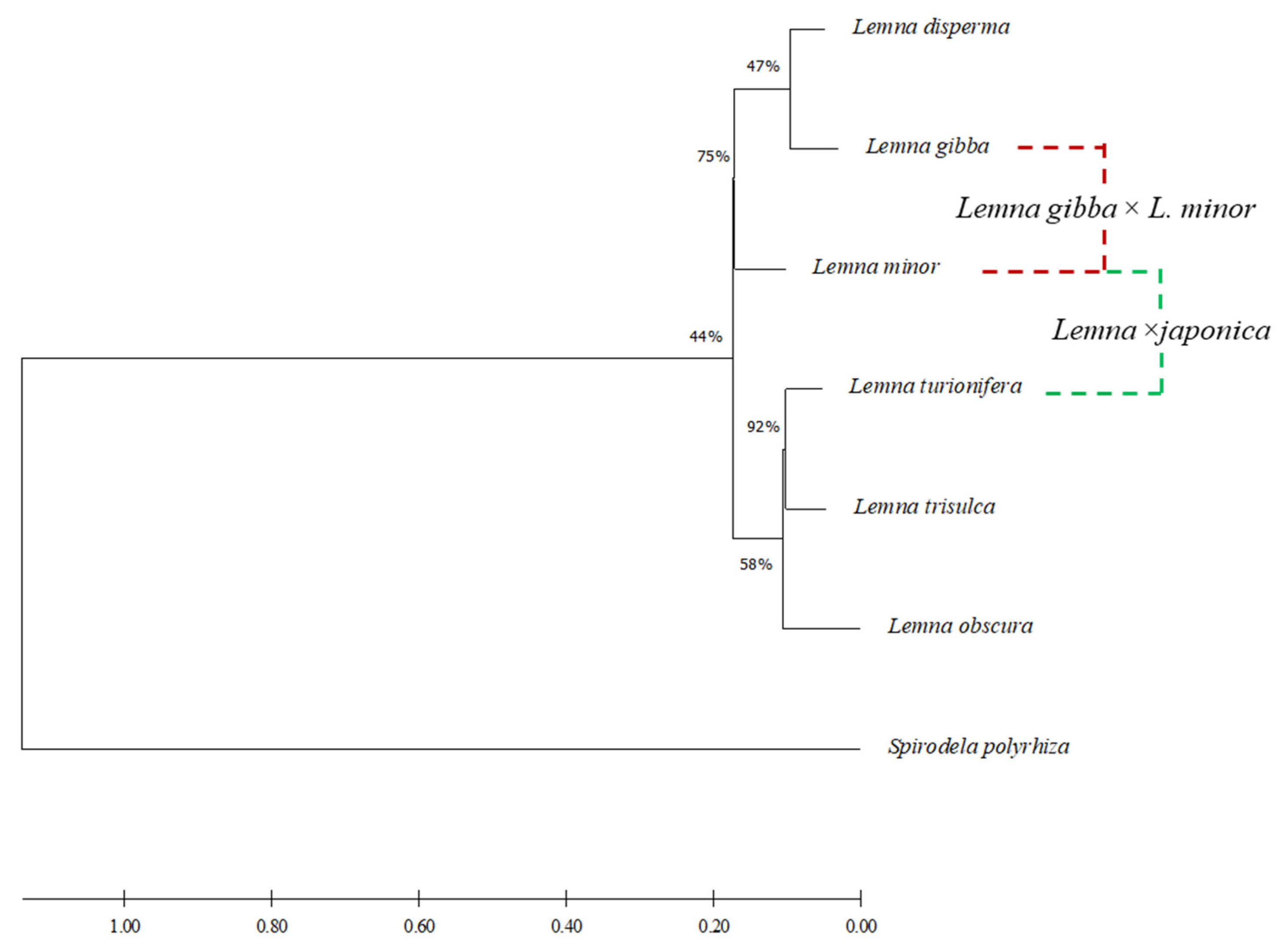
2.5. Infrasectional Structure of Lemna Sect. Lemna
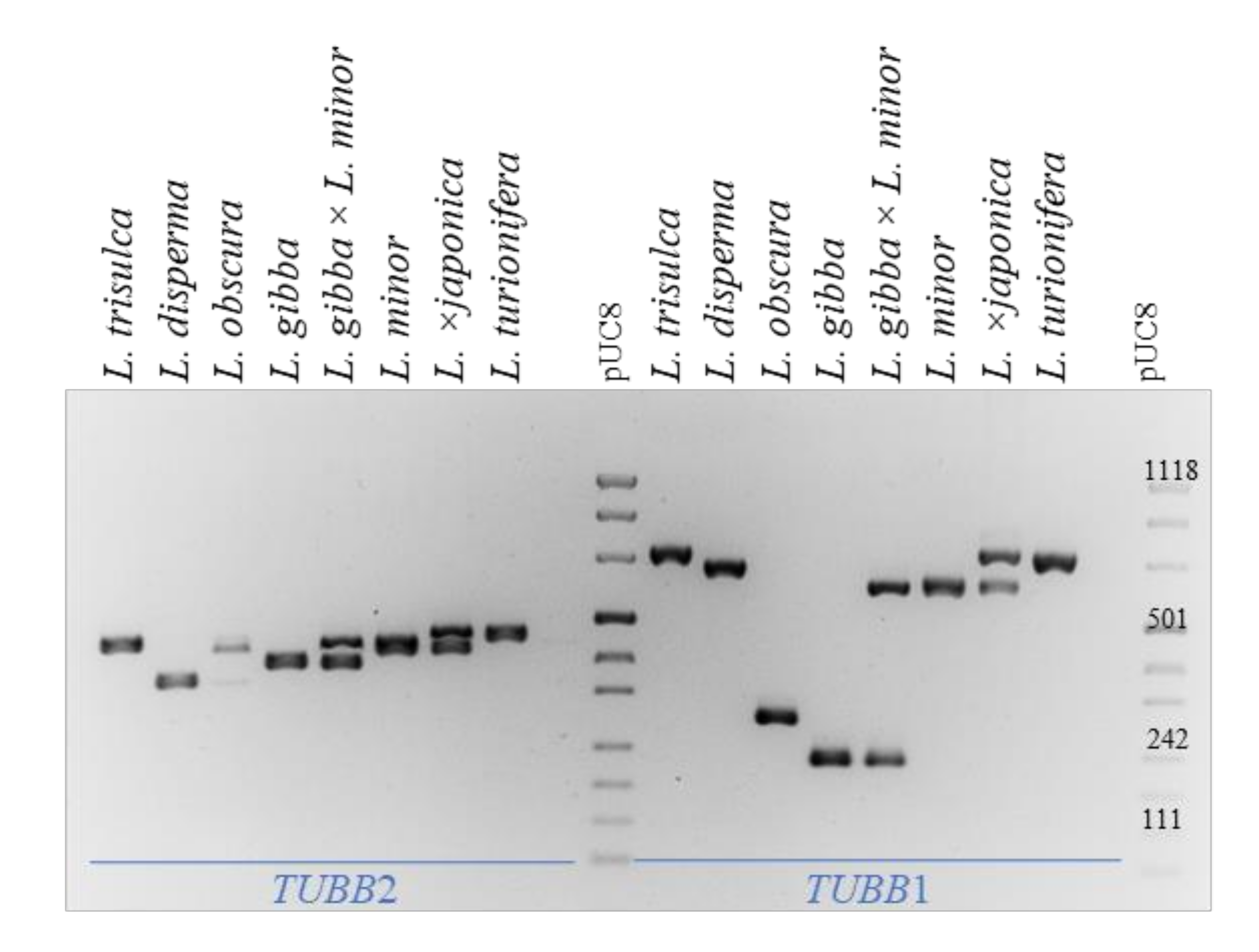
2.6. Intraspecific TUBB2 Polymorphism in Lemna gibba and Lemna minor
3. Discussion
3.1. Lemna ×japonica
3.2. Lemna gibba × Lemna minor
3.3. Lemna sect. Lemna
3.4. Infraspecific Variation
4. Materials and Methods
4.1. Plant Material and Cultivation
4.2. DNA Extraction and TBP Profiling
4.3. DNA Barcoding Analysis
4.4. β-. Tubulin Intron Amplification and Sequencing
4.5. Bioinformatic Sequence Analysis
4.6. Flower Induction
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martinov, I. Techno-Botanical Dictionary (Технo-бoтанический Слoварь); Pechashano v Imperatorskoĭ Tipografii: Saint Petersburg, Russia, 1820. [Google Scholar]
- Tippery, N.P.; Les, D.H. Tiny Plants with Enormous Potential: Phylogeny and Evolution of Duckweeds. In The Duckweed Genomes; Cao, X.H., Fourounjian, P., Wang, W., Eds.; Springer: Berlin, Germany, 2020; pp. 19–38. [Google Scholar] [CrossRef]
- Bog, M.; Appenroth, K.; Sree, K. Key to the determination of taxa of Lemnaceae: An update. Nord. J. Bot. 2020, 38, e02658. [Google Scholar] [CrossRef]
- Les, D.; Crawford, D.; Landolt, E.; Gabel, J.; Kemball, R. Phylogeny and systematics of Lemnaceae, the duckweed family. Syst. Bot. 2002, 27, 221–240. [Google Scholar]
- Bog, M. Genotyping-By-Sequencing for species delimitation in Lemna section Uninerves Hegelm (Lemnaceae). In The Duckweed Genomes; Cao, X.H., Fourounjian, P., Wang, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 115–123. [Google Scholar] [CrossRef]
- De Lange, L. Gibbosity in the complex Lemna gibba/Lemna minor: Literature survey and ecological aspects. Aquat. Bot. 1975, 1, 1327–1332. [Google Scholar] [CrossRef]
- Kandeler, R. Species delimitation in the genus Lemna. Aquat. Bot. 1975, 1, 365–376. [Google Scholar] [CrossRef]
- Landolt, E. Morphological differentiation and geographical distribution of the Lemna gibba-Lemna minor group. Aquat. Bot. 1975, 1, 345–363. [Google Scholar] [CrossRef]
- de Lange, L.; Pieterse, A.H.; van Baarsen-Beckers, I. The occurrence of mixed populations of different genotypes of the Lemna gibba—Lemna minor complex. Acta Bot. Neerl. 1981, 30, 191–197. [Google Scholar] [CrossRef]
- De Lange, L.; Pieterse, A.H. A comparative study of the morphology of Lemna gibba L. and Lemna minor L. Acta Bot. Neerl. 1973, 22, 510–517. [Google Scholar] [CrossRef]
- Giardelli, M.L. Una nueva especia de Lemnacea de la Flora Argentina. Notas Del Mus. De La Plata 1937, 2, 97–100. [Google Scholar]
- Giuga, G. Vita segreta di Lemnacee. In Lemna Symmeter G. Giuga-Species Nova; Blasio: Napoli, Italy, 1973; p. 19. [Google Scholar]
- Bog, M.; Appenroth, K.; Sree, K. Duckweed (Lemnaceae): Its Molecular Taxonomy. Front. Sustain. Food Syst. 2019, 3, 117. [Google Scholar] [CrossRef]
- Wang, W.Q.; Kerstetter, R.; Michael, T. Evolution of genome size in duckweeds (Lemnaceae). J. Bot. 2011, 2011, 570319. [Google Scholar] [CrossRef] [Green Version]
- Hoang, P.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 3234. [Google Scholar] [CrossRef] [PubMed]
- Landolt, E. The family of Lemnaceae—A Monographic Study, Volume 1, Biosystematic investigations in the family of duckweeds (Lemnaceae). Ver. Geobot: Inst. ETH Shift. Rübel 1986, 71, 1–563. [Google Scholar]
- Landolt, E. Contribution on the Lemnaceae of Ecuador. Fragm. Flor. Geobot. 2000, 45, 221–237. [Google Scholar]
- Landolt, E. Description of six new species of Lemnaceae. E-Period. 1980, 70, 13–21. [Google Scholar]
- Hirahaya, M.; Kadono, Y. Biosystematic study of Lemna minor L. sensu lato (Lemnaceae) in Japan with special reference to allozyme variation. Acta Phytotaxon. Geobot. 1995, 46, 117–129. [Google Scholar]
- Braglia, L.; Lauria, M.; Appenroth, K.; Bog, M.; Breviario, D.; Grasso, A.; Gavazzi, F.; Morello, L. Duckweed Species Genotyping and Interspecific Hybrid Discovery by Tubulin-Based Polymorphism Fingerprinting. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Tippery, N.P.; Les, D.H.; Crawford, D.J. Evaluation of phylogenetic relationships in Lemnaceae using nuclear ribosomal data. Plant Biol. 2015, 17, 50–58. [Google Scholar] [CrossRef]
- Ceschin, S.; Abati, S.; Leacche, I.; Zuccarello, V. Ecological comparison between duckweeds in central Italy: The invasive Lemna minuta vs the native L. minor. Plant Biosyst. 2018, 152, 674–683. [Google Scholar] [CrossRef]
- Ceschin, S.; Leacche, I.; Pascucci, S.; Abati, S. Morphological study of Lemna minuta Kunth, an alien species often mistaken for the native L. minor L. (Araceae). Aquat. Bot. 2016, 131, 51–56. [Google Scholar] [CrossRef]
- Bog, M.; Baumbach, H.; Schween, U.; Hellwig, F.; Landolt, E.; Appenroth, K. Genetic structure of the genus Lemna L. (Lemnaceae) as revealed by amplified fragment length polymorphism. Planta 2010, 232, 609–619. [Google Scholar] [CrossRef]
- Bog, M.; Landrock, M.; Drefahl, D.; Sree, K.; Appenroth, K. Fingerprinting by amplified fragment length polymorphism (AFLP) and barcoding by three plastidic markers in the genus Wolffiella Hegelm. Plant Syst. Evol. 2018, 304, 373–386. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Y.; Yan, Y.; Ermakova, M.; Kerstetter, R.; Messing, J. DNA barcoding of the Lemnaceae, a family of aquatic monocots. BMC Plant Biol. 2010, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borisjuk, N.; Chu, P.; Gutierrez, R.; Zhang, H.; Acosta, K.; Friesen, N.; Sree, K.; Garcia, C.; Appenroth, K.; Lam, E. Assessment, validation and deployment strategy of a two-barcode protocol for facile genotyping of duckweed species. Plant Biol. 2015, 17, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Bardini, M.; Lee, D.; Donini, P.; Mariani, A.; Gianì, S.; Toschi, M.; Lowe, C.; Breviario, D. Tubulin-based polymorphism (TBP): A new tool, based on functionally relevant sequences, to assess genetic diversity in plant species. Genome 2004, 47, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Braglia, L.; Gavazzi, F.; Morello, L.; Gianì, S.; Nick, P.; Breviario, D. On the applicability of the Tubulin-Based Polymorphism (TBP) genotyping method: A comprehensive guide illustrated through the application on different genetic resources in the legume family. Plant Methods 2020, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Gavazzi, F.; Braglia, L.; Mastromauro, F.; Giani, S.; Morello, L.; Breviario, D. The Tubulin-Based-Polymorphism Method Provides a Simple and Effective Alternative to the Genomic Profiling of Grape. PLoS ONE 2016, 11, e0163335. [Google Scholar] [CrossRef] [PubMed]
- Van Hoeck, A.; Horemans, N.; Monsieurs, P.; Cao, H.; Vandenhove, H.; Blust, R. The first draft genome of the aquatic model plant Lemna minor opens the route for future stress physiology research and biotechnological applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Sree, K.; Bog, M.; Appenroth, K. Taxonomy of duckweeds (Lemnaceae), potential new crop plants. Emir. J. Food Agric. 2016, 28, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Huang, M.; Han, B.; Sun, X.; Sree, K.; Appenroth, K.; Zhang, J. Flower induction, microscope-aided cross-pollination, and seed production in the duckweed Lemna gibba with discovery of a male-sterile clone. Sci. Rep. 2017, 7, 3047. [Google Scholar] [CrossRef] [Green Version]
- Fourounjian, P.; Slovin, J.; Messing, J. Flowering and Seed Production across the Lemnaceae. Int. J. Mol. Sci. 2021, 22, 2733. [Google Scholar] [CrossRef]
- Creer, S. Choosing and using introns in molecular phylogenetics. Evol. Bioinform. Online 2007, 3, 99–108. [Google Scholar] [CrossRef]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.; Baird, S.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, A.; Abbott, R. Hybrid swarms: Catalysts for multiple evolutionary events in Senecio in the British Isles. Plant Ecol. Divers. 2015, 8, 449–463. [Google Scholar] [CrossRef] [Green Version]
- Novak, S.; Soltis, D.; Soltis, P. Ownbey Tragopogons—40 Years Later. Am. J. Bot. 1991, 78, 1586–1600. [Google Scholar] [CrossRef]
- Renaut, S.; Rowe, H.C.; Ungerer, M.C.; Rieseberg, L.H. Genomics of homoploid hybrid speciation: Diversity and transcriptional activity of long terminal repeat retrotransposons in hybrid sunflowers. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130345. [Google Scholar] [CrossRef] [Green Version]
- Marconi, G.; Landucci, F.; Rosellini, D.; Venanzoni, R.; Albertini, E. DNA barcoding as a tool for early warning and monitoring alien duckweeds (Lemna sp.pl.): The case of Central Italy. Plant Biosyst. 2019, 153, 660–668. [Google Scholar] [CrossRef]
- Mallet, J. Hybrid speciation. Nature 2007, 446, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Benoit, L.K.; Les, D.H.; King, U.M.; Na, H.R.; Chen, L.; Tippery, N.P. Extensive interlineage hybridization in the predominantly clonal Hydrilla verticillata. Am. J. Bot. 2019, 106, 1622–1637. [Google Scholar] [CrossRef] [PubMed]
- Sree, K.; Sudakaran, S.; Appenroth, K. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 204. [Google Scholar] [CrossRef]
- Xu, S.; Stapley, J.; Gablenz, S.; Boyer, J.; Appenroth, K.; Sree, K.; Gershenzon, J.; Widmer, A.; Huber, M. Low genetic variation is associated with low mutation rate in the giant duckweed. Nat. Commun. 2019, 10, 1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenk, R.; Hildebrandt, A. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous Plant-Cell Cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.; Ryan, P. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone ID | Collection | Continent/Region | Country | Classification System | |
|---|---|---|---|---|---|
| Morphological Characters | TBP Analysis | ||||
| 0050 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 0078 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 0150 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 0190 | Landolt Collection | North America | USA | L. japonica | L. gibba * |
| 0198 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 6580 | Landolt Collection | North America | USA | L. minor | L. ×japonica * |
| 6591 | Landolt Collection | North America | USA | L. minor | L. minor |
| 6619 | Jena University | North America | USA | L. turionifera | L. turionifera |
| 6728 | Jena University | North America | USA | L. turionifera | L. ×japonica * |
| 6742 | Landolt Collection | North America | USA | L. japonica | L. ×japonica |
| 6745 | Landolt Collection | North America | USA | L. gibba | L. gibba |
| 6853 | Jena University | North America | Canada | L. turionifera | L. turionifera |
| 6861 | Landolt Collection | Europe | Italy | L. gibba | L. gibba × L. Minor * |
| 7018 | Landolt Collection | Asia | Turkey | L. minor | L. minor |
| 7021 | Landolt Collection | Europe | Spain | L. gibba | L. ×japonica * |
| 7123 | Landolt Collection | North America | Canada | L. minor | L. ×japonica * |
| 7182 | Landolt Collection | East Asia | Japan | L. japonica | L. ×japonica |
| 7295 | Landolt Collection | Africa | Libya | L. minor | L. minor |
| 7320 | Landolt Collection | Africa | Egypt | L. gibba | L. gibba × L. Minor * |
| 7427 | Landolt Collection | East Asia | Japan | L. turionifera | L. turionifera |
| 7432 | Landolt Collection | East Asia | Japan | L. japonica | L. turionifera * |
| 7537 | Landolt Collection | Europe | Spain | L. gibba | L. ×japonica * |
| 7641 | Landolt Collection | Asia | Israel | L. gibba | L. gibba × L. Minor * |
| 7683 | Landolt Collection | Asia | South Korea | L. turionifera | L. turionifera |
| 7705 | Landolt Collection | India | India | L. gibba | L. gibba |
| 7767 | Landolt Collection | Oceania | Australia | L. disperma | L. disperma |
| 7777 | Landolt Collection | Oceania | Australia | L. disperma | L. disperma |
| 7798 | Landolt Collection | South America | Peru | L. gibba | L. gibba |
| 7816 | Landolt Collection | Oceania | Australia | L. disperma | L. disperma |
| 7856 | Landolt Collection | North America | USA | L. obscura | L. obscura |
| 7868 | Jena University | Europe | Ireland | L. japonica | L. ×japonica |
| 7922 | Landolt Collection | South America | Argentina | L. gibba | L. gibba |
| 7951 | Landolt Collection | Asia | China | L. turionifera | L. turionifera |
| 8227 | Landolt Collection | North America | USA | L. obscura | L. obscura |
| 8428 | Landolt Collection | Europe | Switzerland | L. gibba | L. gibba |
| 8434 | Landolt Collection | North America | Canada | L. minor | L. ×japonica * |
| 8653 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 8697 | Landolt Collection | East Asia | Japan | L. japonica | L. ×japonica |
| 8717 | Landolt Collection | Oceania | Australia | L. L. disperma | L. disperma |
| 8760 | Landolt Collection | Europe | Czech Republic | L. turionifera | L. turionifera |
| 8892 | Landolt Collection | North America | USA | L. obscura | L. obscura |
| 9016 | Landolt Collection | East Asia | Japan | L. japonica | L. ×japonica |
| 9109 | Jena University | Europe | Poland | L. turionifera | L. turionifera |
| 9223 | Landolt Collection | Europe | United Kingdom | L. minor | L. minor |
| 9240 | Landolt Collection | Europe, Asia | Russia | L. minor | L. minor |
| 9248 | Landolt Collection | Europe | Italy | L. gibba | L. gibba × L. Minor * |
| 9250 | Landolt Collection | Europe | Finland | L. japonica | L. ×japonica |
| 9253 | Landolt Collection | Europe | Finland | L. minor | L. minor |
| 9254 | Landolt Collection | Europe | Finland | L. turionifera | L. turionifera |
| 9285 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 9330 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 9345 | Landolt Collection | Europe | Switzerland | L. minor | L. minor |
| 9352 | Landolt Collection | Europe | Albania | L. gibba | L. gibba |
| 9421 | Landolt Collection | North America | USA | L. japonica | L. ×japonica |
| 9424 | Landolt Collection | Europe | Germany | L. minor | L. minor |
| 9429 | Jena University | Europe, Asia | Russia | L. turionifera | L. ×japonica * |
| 9435 | Landolt Collection | Europe | Albania | L. gibba | L. gibba |
| 9438 | Landolt Collection | Europe | Czech Republic | L. minor | L. minor |
| 9439 | Landolt Collection | Europe | Germany | L. minor | L. ×japonica * |
| 9470 | Landolt Collection | Europe | United Kingdom | L. turionifera | L. turionifera |
| 9471 | Jena University | Europe | United Kingdom | L. turionifera | L. turionifera |
| 9478 | Jena University | Europe | Poland | L. turionifera | L. turionifera |
| 9480 | Landolt Collection | Europe, Asia | Russia | L. turionifera | L. turionifera |
| 9482 | Landolt Collection | Europe | Italy | L. minor | L. minor |
| 9483 | Landolt Collection | Europe | Albania | L. minor | L. ×japonica * |
| 9485 | Landolt Collection | Europe | Ireland | L. minor | L. minor |
| 9532 | Landolt Collection | Europe | Macedonia | L. minor | L. ×japonica * |
| 9534 | Landolt Collection | Europe | Germany | L. minor | L. minor |
| 9542 | Landolt Collection | Europe | Italy | L. minor | L. ×japonica * |
| 9561 | Landolt Collection | Europe | Sweden | L. minor | L. minor |
| 9562 | Jena University | Europe | Germany | L. gibba | L. gibba × L. Minor * |
| 9574 | Landolt Collection | Oceania | New Zealand | L. minor | L. minor |
| 9577 | Landolt Collection | Europe | Italy | L. gibba | L. gibba |
| 9591 | Landolt Collection | Europe | Hungary | L. gibba | L. ×japonica * |
| 9598 | Landolt Collection | Europe | Germany | L. gibba | L. gibba |
| 9660 | Landolt Collection | Asia | China | L. japonica | L. ×japonica |
| 9942 | Landolt Collection | Europe | Norway | L. minor | L. minor |
| 9951 | Landolt Collection | Europe | France | L. gibba | L. ×japonica * |
| 9952 | Landolt Collection | Europe | France | L. minor | L. minor |
| 9961 | Landolt Collection | Europe | Germany | L. minor | L. minor |
| 9965 | Landolt Collection | Europe | Switzerland | L. gibba | L. ×japonica * |
| 9967 | Landolt Collection | Europe | Switzerland | L. minor | L. minor |
| 9969 | Landolt Collection | Europe | Switzerland | L. minor | L. ×japonica * |
| 9973 | Landolt Collection | Europe | Germany | L. minor | L. minor |
| 9977 | Landolt Collection | Europe | Germany | L. minor | L. minor |
| 9978 | Landolt Collection | Europe | Switzerland | L. minor | L. ×japonica * |
| 9979 | Landolt Collection | Europe | Germany | L. minor | L. minor |
| 9980 | Landolt Collection | Europe | Germany | L. minor | L. ×japonica * |
| 9982 | Landolt Collection | North America | USA | L. japonica | L. ×japonica |
| 9983 | Landolt Collection | Europe | Switzerland | L. japonica | L. ×japonica |
| 9986 | Landolt Collection | North America | USA | L. minor | L. ×japonica * |
| 9991 | Landolt Collection | North America | USA | L. japonica | L. ×japonica |
| 8784b | Landolt Collection | Europe | Sweden | L. japonica | L. ×japonica |
| 9425a | Landolt Collection | Europe | Italy | L. gibba | L. gibba × L. Minor * |
| BOG0024 | Greifswald University | Europe | Germany | L. turionifera | L. turionifera |
| BOG0071 | Greifswald University | Europe | Germany | L. turionifera | L. turionifera |
| BOG0072 | Greifswald University | Europe | Germany | L. turionifera | L. turionifera |
| KJA007 | Jena University | Europe, Asia | Russia | L. turionifera | L. turionifera |
| TBP | ||||||||
|---|---|---|---|---|---|---|---|---|
| Species | L. minor | L. turionifera | L. ×japonica | L. gibba | L. gibba × L. minor | Tot. n | % Incorrect | |
| morphology | L. minor | 21 | 11 | 32 | 34.3 | |||
| L. turionifera | 16 | 2 | 18 | 11.1 | ||||
| L.×japonica | 1 | 19 | 1 | 21 | 9.5 | |||
| L. gibba | 5 | 9 | 6 | 20 | 55.0 | |||
| L. gibba× L. minor | 0 | 0 | 0.0 | |||||
| tot. n | 21 | 17 | 37 | 10 | 6 | 91 | 28.6 | |
| % incorrect | 0.0 | 5.9 | 48.6 | 10.0 | 100.0 | 28.6 | ||
| Haplotype | Polymorphic Sites | Alignment Length (bp) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 35 | 45 | 86 | 175 | 177 | 184 | 197 | 206 | 249 | 286 | 316 | 330 | 337 | 366 | 407 | ||
| L. minor | M1 | T | A | T | G | C | C | T | T | A | C | G | G | G | A | |
| M2 | C | G | G | T | G | T | A | G | T | T | A | A | A | T | ||
| M3 | T | A | T | T | C | C | T | T | A | C | G | G | G | A | ||
| M4 | T | A | T | T | C | C | T | T | A | C | A | G | G | A | ||
| 40 | 68 | 109 | 224 | 250 | 255 | 302 | 329 | 380 | ||||||||
| L. gibba | G1 | T | T | C | G | C | T | A | G | |||||||
| G2 | -- | T | C | G | T | T | A | G | ||||||||
| G3 | -- | A | T | -- | T | C | C | T | ||||||||
| Clone | Origin | Haplotype | SNPs | Clone | Origin | Haplotype | SNPs | ||
|---|---|---|---|---|---|---|---|---|---|
| L. minor | 5500 | Ireland | M1 | 0 | L. gibba | 7742a | Italy | G1 | 0 |
| 9961 | Germany | M1 | 0 | 9598 | Italy | G1 | 0 | ||
| 9424 | Germany | M1 | 0 | 6745 | California—USA | G3 | 6 | ||
| 7194 | Uganda | M2 | 12 | 8124 | Arizona—USA | G3 | 6 | ||
| 7753 | Ethiopia | M2 | 12 | 7705 | India | G2 | 2 | ||
| 7210 | S. Africa | M2 | 12 | 190 | USA | G2 | 2 | ||
| 9495 | Norway | M3 | 1 | 7796 | Italy | G2 | 2 | ||
| LM0011 | Russia | M3 | 1 | 9583 | Poland | n.d | -- | ||
| 7766 | New Zealand | M3 | 1 | 7922 | Argentina | n.d | -- | ||
| 9536 | Germany | M4 | 2 | 9614 | Poland | n.d | -- | ||
| 7022 | Spain | M4 | 2 | 9619 | Albany | n.d | -- | ||
| 9482 | Italy | M4 | 2 | 7245 | S. Africa | n.d | -- | ||
| 9942 | Norway | M4 | 2 | 8428 | Switzerland | n.d | -- | ||
| 9533 | Macedonia | M4 | 2 | L. gibba× L. minor | 7320 | Egypt | G2 M4 | ||
| 8744 | Albania | M4 | 2 | 6861 | Italy | G2 M4 | |||
| LM0010 | Italy | M4 | 2 | 9562 | Italy | G2 M4 | |||
| LM0008 | Russia | M4 | 2 | 9425a | Germany | G2 M4 | |||
| 9252 | Finland | M4 | 2 | 7641 | Israel | G2 M4 | |||
| 8292 | Iran | M4 | 2 | 9248 | Italy | G2 M1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braglia, L.; Breviario, D.; Gianì, S.; Gavazzi, F.; De Gregori, J.; Morello, L. New Insights into Interspecific Hybridization in Lemna L. Sect. Lemna (Lemnaceae Martinov). Plants 2021, 10, 2767. https://doi.org/10.3390/plants10122767
Braglia L, Breviario D, Gianì S, Gavazzi F, De Gregori J, Morello L. New Insights into Interspecific Hybridization in Lemna L. Sect. Lemna (Lemnaceae Martinov). Plants. 2021; 10(12):2767. https://doi.org/10.3390/plants10122767
Chicago/Turabian StyleBraglia, Luca, Diego Breviario, Silvia Gianì, Floriana Gavazzi, Jacopo De Gregori, and Laura Morello. 2021. "New Insights into Interspecific Hybridization in Lemna L. Sect. Lemna (Lemnaceae Martinov)" Plants 10, no. 12: 2767. https://doi.org/10.3390/plants10122767