
Protein, Amino Acid, Oil, Fatty Acid, Sugar, Anthocyanin, Isoflavone, Lutein, and Antioxidant Variations in Colored Seed-Coated Soybeans

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Soybean Genotypes
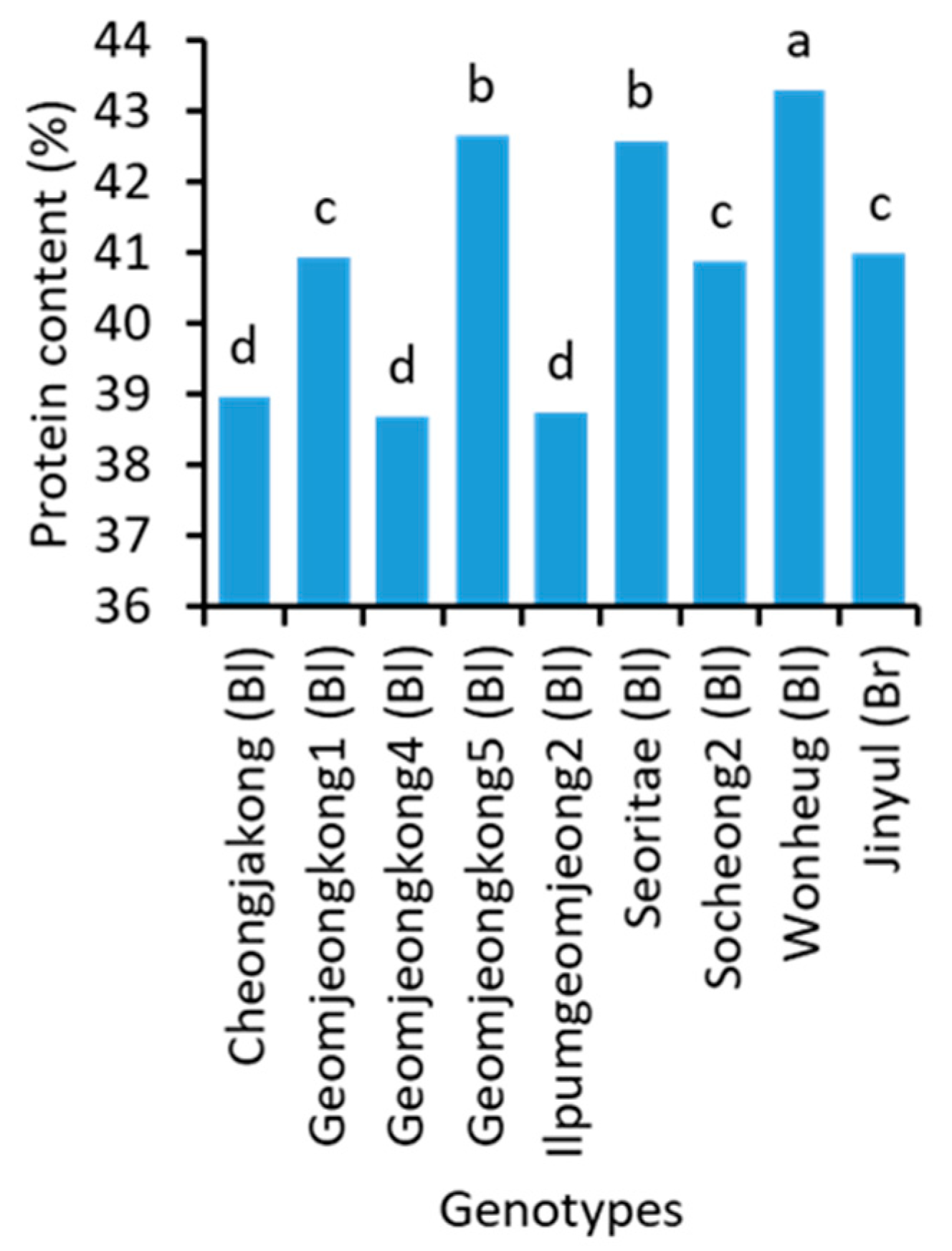
2.2. Protein and Amino Acids
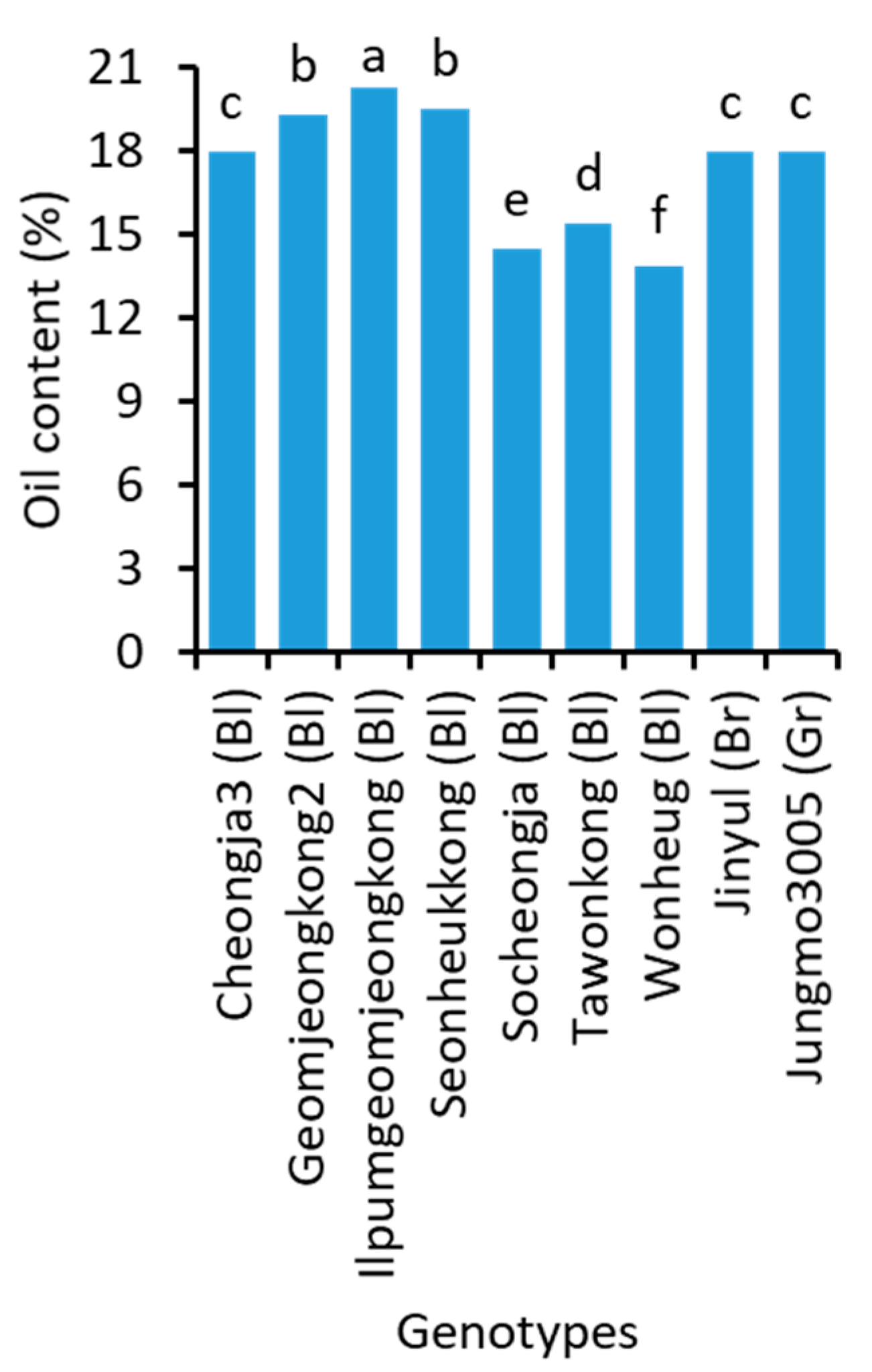
2.3. Oil Content and Fatty Acid Composition
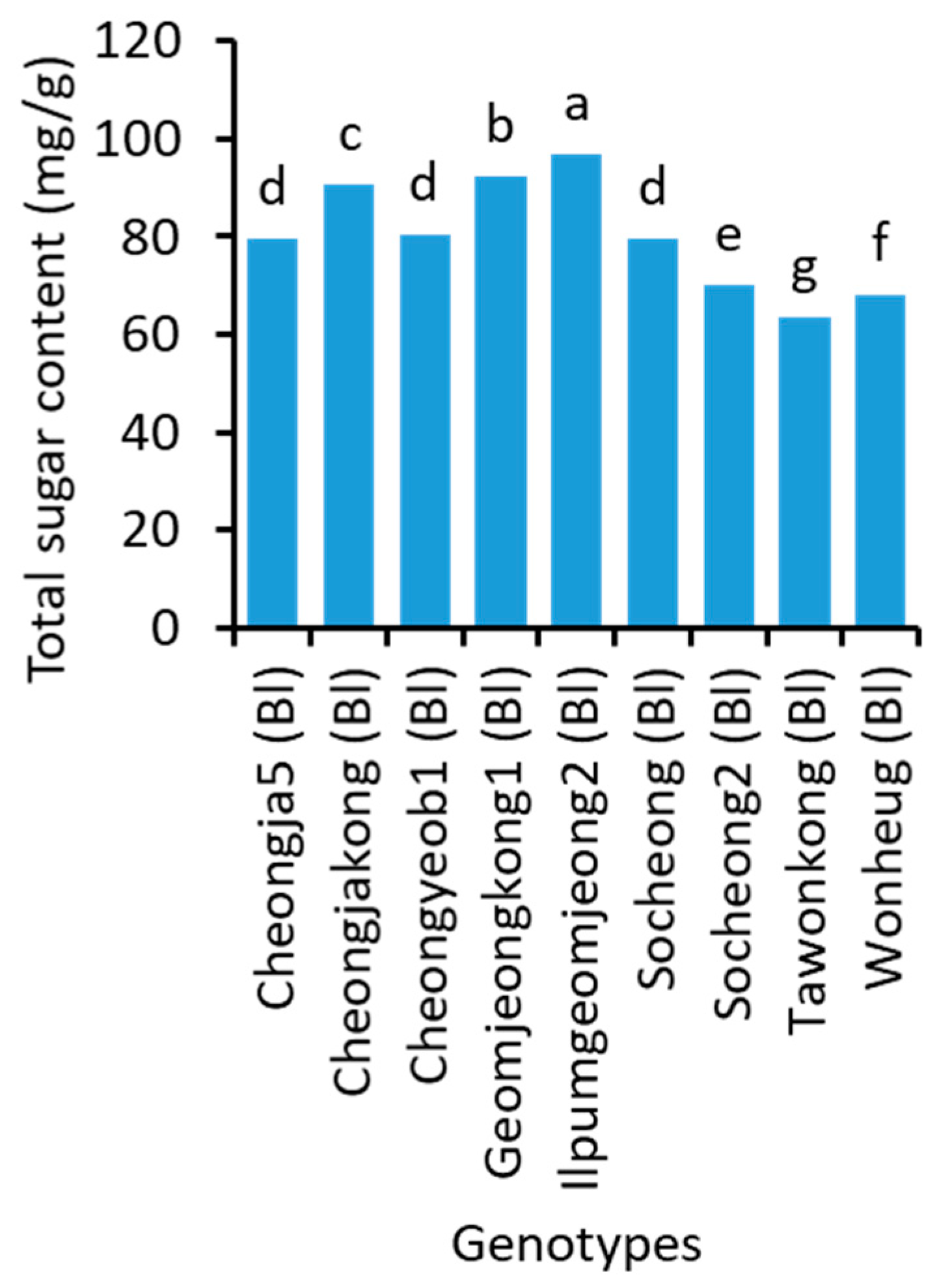
2.4. Sugar Content
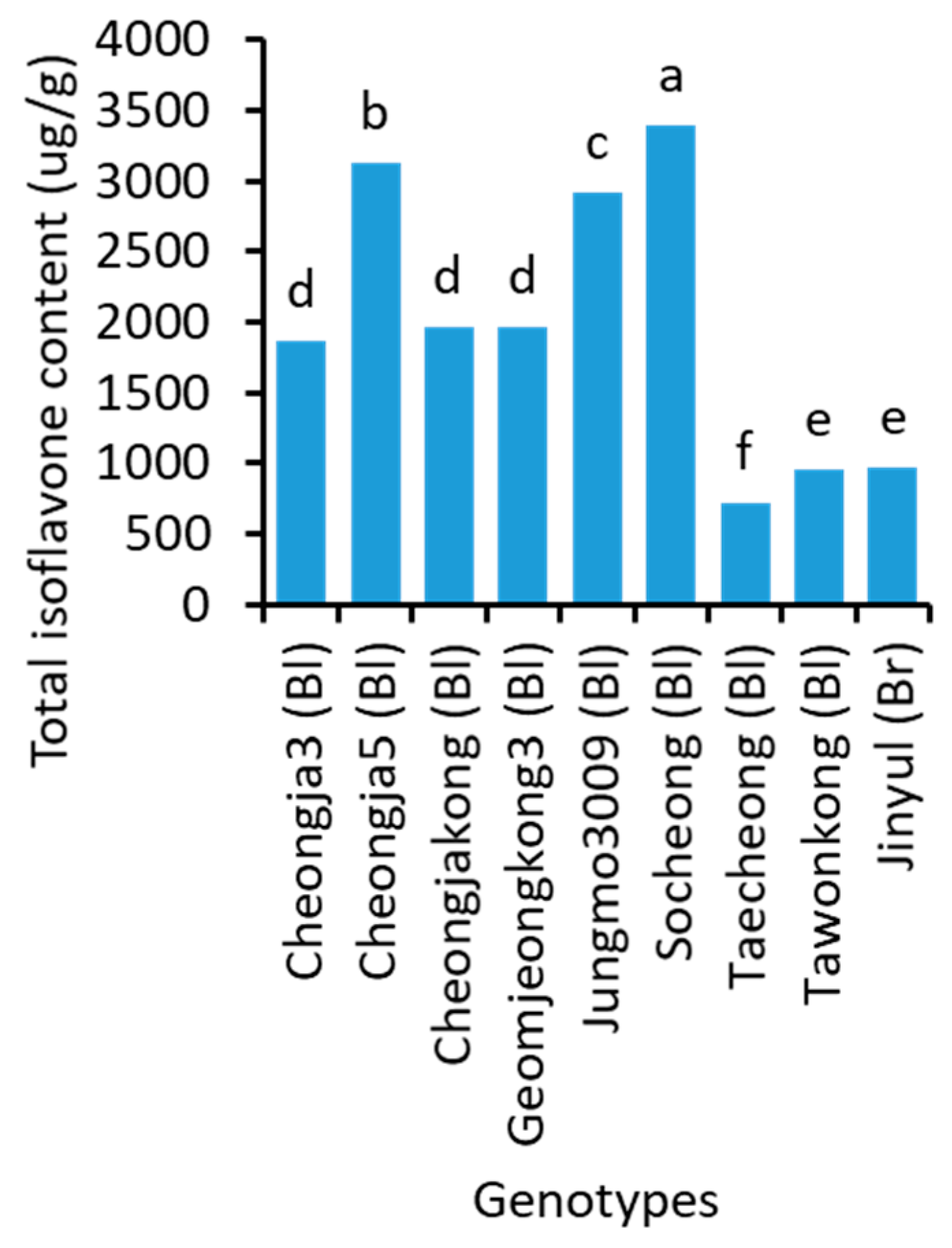
2.5. Isoflavone Content
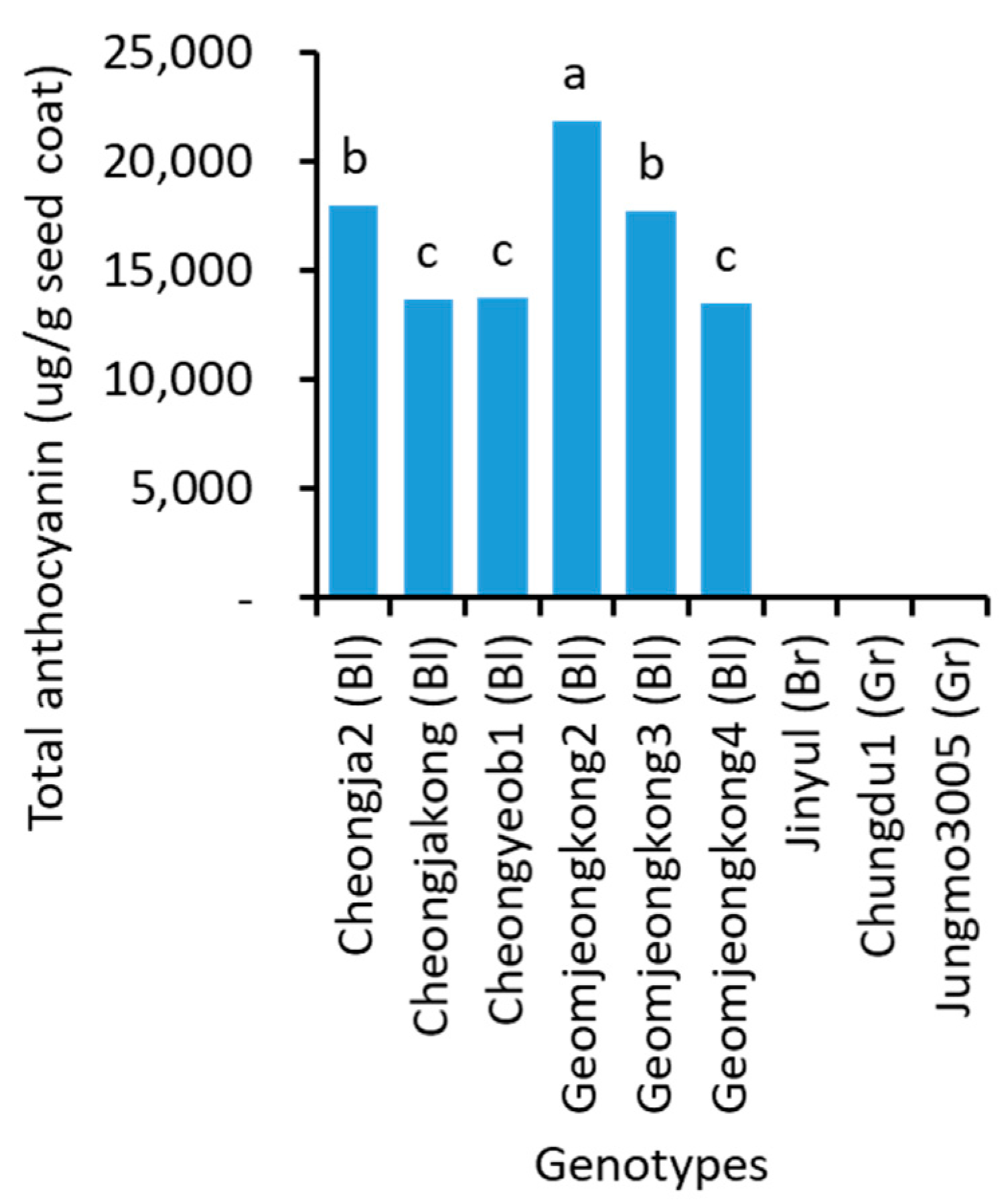
2.6. Anthocyanin Content
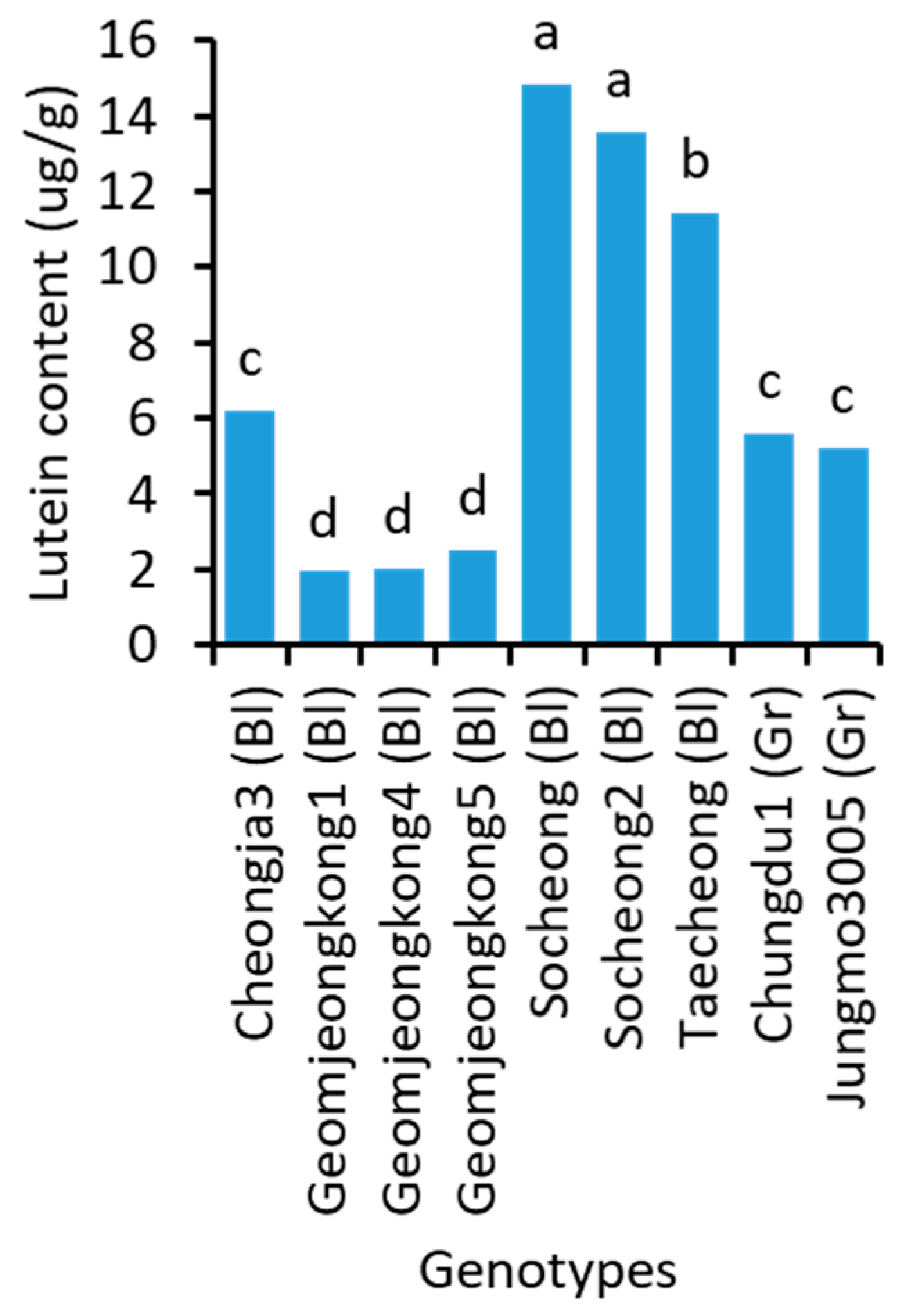
2.7. Lutein Content
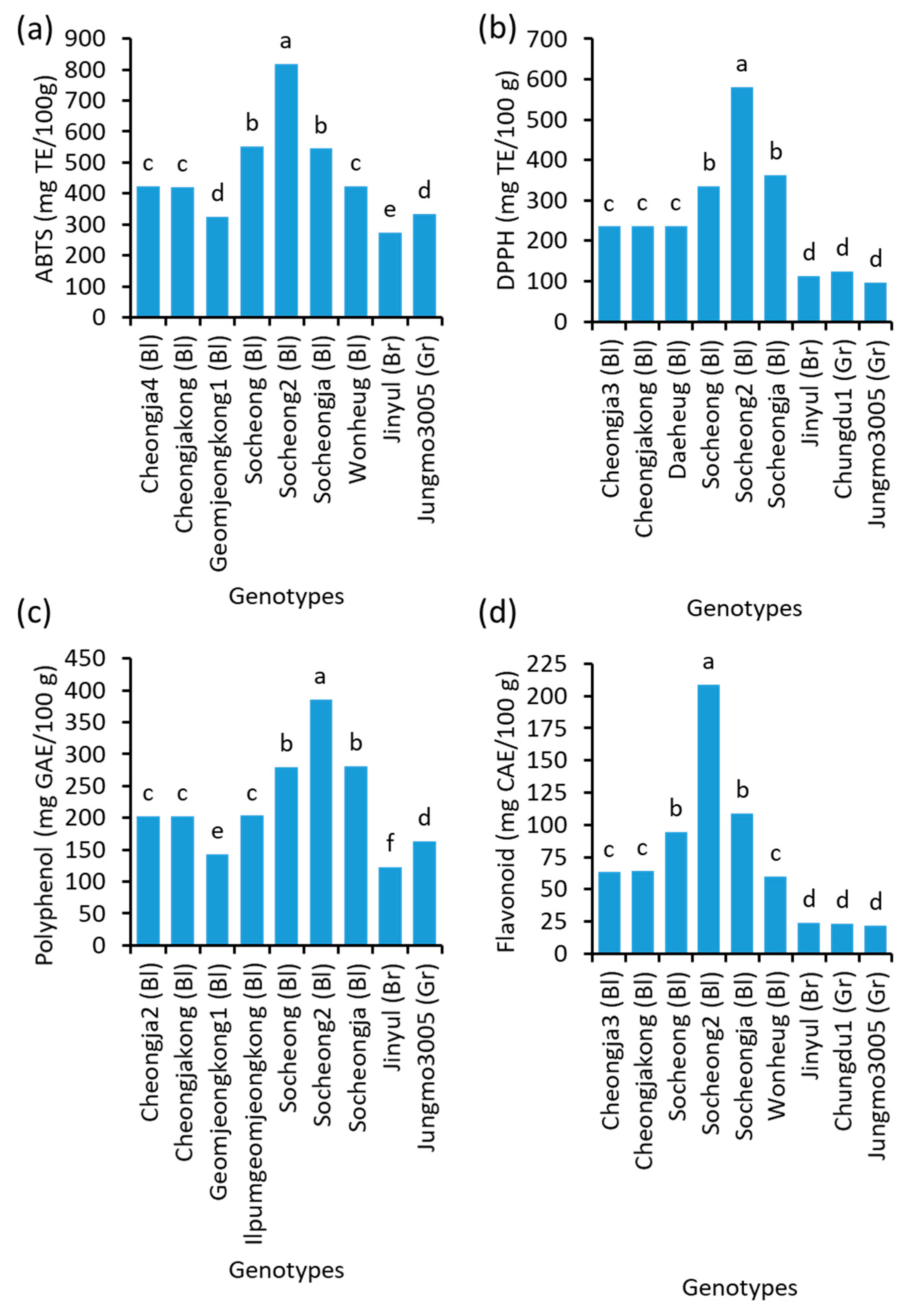
2.8. Antioxidants’ Potential
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Chemicals and Reagents
4.3. Protein and Amino Acids’ Analysis
4.4. Measurement of Oil and Fatty Acids
4.5. Determination of Free Sugar Content
4.6. Isoflavone Analysis
4.7. Anthocyanin Analysis
4.8. Determination of Lutein Content
4.9. Sample Extraction for ABTS, DPPH, Total Polyphenol, and Total Flavonoid Analyses
4.10. ABTS Radical Scavenging Assay
4.11. DPPH Radical Scavenging Assay
4.12. Measurement of Total Polyphenol Content
4.13. Measurement of Total Flavonoid Content
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT Database Collections. Available online: https://www.fao.org (accessed on 30 June 2021).
- Zhao, S.; Xu, X.; Wei, D.; Lin, X.; Qiu, S.; Ciampitti, I.; He, P. Soybean yield, nutrient uptake and stoichiometry under different climate regions of northeast China. Sci. Rep. 2020, 10, 8431. [Google Scholar] [CrossRef]
- Brummer, E.C.; Graef, G.L.; Orf, J.; Wilcox, J.R.; Shoemaker, R.C. Mapping QTL for Seed Protein and Oil Content in Eight Soybean Populations. Crop. Sci. 1997, 37, 370–378. [Google Scholar] [CrossRef]
- Hubert, J.; Berger, M.; Nepveu, F.; Paul, F.; Daydé, J. Effects of fermentation on the phytochemical composition and antioxidant properties of soy germ. Food Chem. 2008, 109, 709–721. [Google Scholar] [CrossRef]
- Lee, J.H.; Cho, K.M. Changes occurring in compositional components of black soybeans maintained at room temperature for different storage periods. Food Chem. 2012, 131, 161–169. [Google Scholar] [CrossRef]
- Lee, J.H.; Kang, N.S.; Shin, S.-O.; Shin, S.-H.; Lim, S.-G.; Suh, D.-Y.; Baek, I.-Y.; Park, K.-Y.; Ha, T.J. Characterisation of anthocyanins in the black soybean (Glycine max L.) by HPLC-DAD-ESI/MS analysis. Food Chem. 2009, 112, 226–231. [Google Scholar] [CrossRef]
- Song, J.; Liu, Z.; Hong, H.; Ma, Y.; Tian, L.; Li, X.; Li, Y.-H.; Guan, R.; Guo, Y.; Qiu, L.-J. Identification and Validation of Loci Governing Seed Coat Color by Combining Association Mapping and Bulk Segregation Analysis in Soybean. PLoS ONE 2016, 11, e0159064. [Google Scholar] [CrossRef]
- Choung, M.-G.; Baek, I.-Y.; Kang, S.-T.; Han, W.-Y.; Shin, D.-C.; Moon, H.-P.; Kang, K.-H. Isolation and Determination of Anthocyanins in Seed Coats of Black Soybean (Glycine max (L.) Merr.). J. Agric. Food Chem. 2001, 49, 5848–5851. [Google Scholar] [CrossRef]
- Clough, S.J.; Tuteja, J.H.; Li, M.; Marek, L.F.; Shoemaker, R.C.; O Vodkin, L. Features of a 103-kb gene-rich region in soybean include an inverted perfect repeat cluster ofCHSgenes comprising theIlocus. Genome 2004, 47, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Ji, R.; Li, Z.; Yu, Y.; Nakano, M.; Long, Y.; Feng, L.; Qin, C.; Lu, D.; Zhan, J.; et al. Soybean DICER-LIKE2 Regulates Seed Coat Color via Production of Primary 22-Nucleotide Small Interfering RNAs from Long Inverted Repeats. Plant Cell 2020, 32, 3662–3673. [Google Scholar] [CrossRef]
- Todd, J.J.; Vodkin, L.O. Duplications That Suppress and Deletions That Restore Expression from a Chalcone Synthase Multigene Family. Plant Cell 1996, 8, 687–699. [Google Scholar] [CrossRef]
- Kim, S.Y.; Wi, H.-R.; Choi, S.; Ha, T.J.; Lee, B.W.; Lee, M. Inhibitory effect of anthocyanin-rich black soybean testa (Glycine max (L.) Merr.) on the inflammation-induced adipogenesis in a DIO mouse model. J. Funct. Foods 2015, 14, 623–633. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K.C. Antioxidant Capacity of Seed Coat, Dehulled Bean, and Whole Black Soybeans in Relation to Their Distributions of Total Phenolics, Phenolic Acids, Anthocyanins, and Isoflavones. J. Agric. Food Chem. 2008, 56, 8365–8373. [Google Scholar] [CrossRef]
- Cho, K.M.; Ha, T.J.; Lee, Y.B.; Seo, W.D.; Kim, J.Y.; Ryu, H.W.; Jeong, S.H.; Kang, Y.M.; Lee, J.H. Soluble phenolics and antioxidant properties of soybean (Glycine max L.) cultivars with varying seed coat colours. J. Funct. Foods 2013, 5, 1065–1076. [Google Scholar] [CrossRef]
- Koh, K.; Youn, J.E.; Kim, H.-S. Identification of anthocyanins in black soybean (Glycine max (L.) Merr.) varieties. J. Food Sci. Technol. 2011, 51, 377–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Wang, C.; Gao, X.; Chen, Y.; Santhanam, R.K.; Wang, C.; Xu, L.; Chen, H. Interaction characterization of preheated soy protein isolate with cyanidin-3-O-glucoside and their effects on the stability of black soybean seed coat anthocyanins extracts. Food Chem. 2019, 271, 266–273. [Google Scholar] [CrossRef]
- Guo, Y.; Ma, M.; Jiang, F.; Jiang, W.; Wang, H.; Du, S.K. Protein quality and antioxidant properties of soymilk derived from black soybean after in vitro simulated gastrointestinal digestion. Int. J. Food Sci. Technol. 2020, 55, 720–728. [Google Scholar] [CrossRef]
- Lim, Y.; Kwon, S.-J.; Qu, S.; Kim, D.-G.; Eom, S. Antioxidant Contributors in Seed, Seed Coat, and Cotyledon of γ-ray-Induced Soybean Mutant Lines with Different Seed Coat Colors. Antioxidants 2021, 10, 353. [Google Scholar] [CrossRef]
- Fathima, A.; Rao, J.R. Selective toxicity of Catechin—a natural flavonoid towards bacteria. Appl. Microbiol. Biotechnol. 2016, 100, 6395–6402. [Google Scholar] [CrossRef]
- Sánchez-Fidalgo, S.; da Silva, M.; Cárdeno, A.; Aparicio-Soto, M.; Salvador, M.; Sawaya, A.; Souza-Brito, A.; Alarcon-De-La-Lastra, C. Abarema cochliacarpos reduces LPS-induced inflammatory response in murine peritoneal macrophages regulating ROS-MAPK signal pathway. J. Ethnopharmacol. 2013, 149, 140–147. [Google Scholar] [CrossRef]
- Brückner, M.; Westphal, S.; Domschke, W.; Kucharzik, T.; Lügering, A. Green tea polyphenol epigallocatechin-3-gallate shows therapeutic antioxidative effects in a murine model of colitis. J. Crohns Colitis 2012, 6, 226–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalia, S.; Marchetti, N.; Bianchi, A. Comparative Evaluation of Different Co-Antioxidants on the Photochemical- and Functional-Stability of Epigallocatechin-3-gallate in Topical Creams Exposed to Simulated Sunlight. Molecules 2013, 18, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Hao, X.; Simi, B.; Ju, J.; Jiang, H.; Reddy, B.S.; Yang, C.S. Green tea polyphenols inhibit colorectal aberrant crypt foci (ACF) formation and prevent oncogenic changes in dysplastic ACF in azoxymethane-treated F344 rats. Carcinogenesis 2007, 29, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.-Y.; Kim, E.-H.; Chung, I.-M.; Ahn, J.-K. Variation of β-carotene Concentration in Soybean Seed and Sprout. Korean J. Crop. Sci. 2012, 57, 324–330. [Google Scholar] [CrossRef]
- Garewal, H. Antioxidants in oral cancer prevention. Am. J. Clin. Nutr. 1995, 62, 1410S–1416S. [Google Scholar] [CrossRef]
- Le Marchand, L.; Hankin, J.H.; Kolonel, L.N.; Beecher, G.R.; Wilkens, L.R.; Zhao, L.P. Intake of specific carotenoids and lung cancer risk. Cancer Epidemiol. Biomark. Prev. 1993, 2, 183–187. [Google Scholar]
- Malenčić, D.; Cvejić, J.; Miladinović, J. Polyphenol Content and Antioxidant Properties of Colored Soybean Seeds from Central Europe. J. Med. Food 2012, 15, 89–95. [Google Scholar] [CrossRef]
- Sharma, K.; Kumar, V.; Kaur, J.; Tanwar, B.; Goyal, A.; Sharma, R.; Gat, Y.; Kumar, A. Health effects, sources, utilization and safety of tannins: A critical review. Toxin Rev. 2019. [Google Scholar] [CrossRef]
- Andres, A.; Donovan, S.; Kuhlenschmidt, M. Soy isoflavones and virus infections. J. Nutr. Biochem. 2009, 20, 563–569. [Google Scholar] [CrossRef]
- Isanga, J.; Zhang, G.-N. Soybean Bioactive Components and their Implications to Health—A Review. Food Rev. Int. 2008, 24, 252–276. [Google Scholar] [CrossRef]
- Park, S.-B.; Lee, J.-H.; Kim, H.-D.; Soe, K.-H.; Jeong, H.-S.; Kim, D.-H.; Lee, S.-E. Screening of Plant Extracts with Cholinesterase Inhibition Activity. Korean J. Plant. Resour. 2018, 31, 433–452. [Google Scholar] [CrossRef]
- Lee, K.J.; Baek, D.-Y.; Lee, G.-A.; Cho, G.-T.; So, Y.-S.; Lee, J.-R.; Ma, K.-H.; Chung, J.-W.; Hyun, D.Y. Phytochemicals and Antioxidant Activity of Korean Black Soybean (Glycine max L.) Landraces. Antioxidants 2020, 9, 213. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-M.; Yoon, H.; Lee, S.; Ko, H.-C.; Shin, M.-J.; Lee, M.-C.; Oh, S.; Desta, K.T. Comparison of Isoflavone Composition and Content in Seeds of Soybean (Glycine max (L.) Merrill) Germplasms with Different Seed Coat Colors and Days to Maturity. Korean J. Plant. Resour. 2020, 33, 558–577. [Google Scholar] [CrossRef]
- Bursać, M.; Krstonošić, M.A.; Miladinović, J.; Malenčić, Đ.; Gvozdenović, L.; Cvejić, J.H. Isoflavone Composition, Total Phenolic Content and Antioxidant Capacity of Soybeans with Colored Seed Coat. Nat. Prod. Commun. 2017, 12, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-M.; Yoon, H.; Lee, S.; Ko, H.-C.; Shin, M.-J.; Lee, M.C.; Hur, O.S.; Ro, N.Y.; Desta, K.T. Isoflavones, anthocyanins, phenolic content, and antioxidant activities of black soybeans (Glycine max (L.) Merrill) as affected by seed weight. Sci. Rep. 2020, 10, 19960. [Google Scholar] [CrossRef] [PubMed]
- Dhungana, S.K.; Kulkarni, K.P.; Kim, M.; Ha, B.-K.; Kang, S.; Song, J.T.; Shin, D.-H.; Lee, A.J.-D. Environmental Stability and Correlation of Soybean Seed Starch with Protein and Oil Contents. Plant. Breed. Biotechnol. 2017, 5, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Rani, A.; Solanki, S.; Hussain, S. Influence of growing environment on the biochemical composition and physical characteristics of soybean seed. J. Food Compos. Anal. 2006, 19, 188–195. [Google Scholar] [CrossRef]
- Qin, P.; Song, W.; Yang, X.; Sun, S.; Zhou, X.; Yang, R.; Li, N.; Hou, W.; Wu, C.; Han, T.; et al. Regional Distribution of Protein and Oil Compositions of Soybean Cultivars in China. Crop. Sci. 2014, 54, 1139–1146. [Google Scholar] [CrossRef]
- Assefa, Y.; Bajjalieh, N.; Archontoulis, S.; Casteel, S.; Davidson, D.; Kovács, P.; Naeve, S.; Ciampitti, I.A. Spatial Characterization of Soybean Yield and Quality (Amino Acids, Oil, and Protein) for United States. Sci. Rep. 2018, 8, 14653. [Google Scholar] [CrossRef] [Green Version]
- Zarkadas, C.G.; Voldeng, H.D.; Yu, Z.R.; Choi, V.K. Assessment of the protein quality of nine northern adapted yellow and brown seed coated soybean cultivars by amino acid analysis. J. Agric. Food Chem. 1999, 47, 5009–5018. [Google Scholar] [CrossRef]
- Carrera, C.S.; Reynoso, C.M.; Funes, G.J.; Martínez, M.J.; Dardanelli, J.; Resnik, S.L. Amino acid composition of soybean seeds as affected by climatic variables. Pesqui. Agropecuária 2011, 46, 1579–1587. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, Y.-S.; Kim, S.T.; Yoon, W.-B.; Han, W.Y.; Kang, I.-K.; Choung, M.-G. Seed coat color and seed weight contribute differential responses of targeted metabolites in soybean seeds. Food Chem. 2017, 214, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Sobko, O.; Stahl, A.; Hahn, V.; Zikeli, S.; Claupein, W.; Gruber, S. Environmental Effects on Soybean (Glycine max (L.) Merr) Production in Central and South Germany. Agronomy 2020, 10, 1847. [Google Scholar] [CrossRef]
- Choi, Y.-M.; Yoon, H.; Shin, M.-J.; Lee, Y.; Hur, O.S.; Lee, B.C.; Ha, B.-K.; Wang, X.; Desta, K.T. Change in Protein, Oil and Fatty Acid Contents in Soybeans (Glycine max (L.) Merr.) of Different Seed Coat Colors and Seed Weight. bioRxiv 2021. [Google Scholar] [CrossRef]
- Shin, E.-C.; Hwang, C.-E.; Lee, B.-W.; Kim, H.-T.; Ko, J.-M.; Baek, I.-Y.; Lee, Y.-B.; Choi, J.-S.; Cho, E.-J.; Seo, W.-T.; et al. Chemometric Approach to Fatty Acid Profiles in Soybean Cultivars by Principal Component Analysis (PCA). Prev. Nutr. Food Sci. 2012, 17, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Abdelghany, A.M.; Zhang, S.; Azam, M.; Shaibu, A.S.; Feng, Y.; Li, Y.; Tian, Y.; Hong, H.; Li, B.; Sun, J. Profiling of seed fatty acid composition in 1025 Chinese soybean accessions from diverse ecoregions. Crop. J. 2020, 8, 635–644. [Google Scholar] [CrossRef]
- Dhakal, K.H.; Jeong, Y.-S.; Lee, J.-D.; Baek, I.-Y.; Ha, T.-J.; Hwang, Y.-H. Fatty acid composition in each structural part of soybean seed and sprout. J. Crop. Sci. Biotechnol. 2009, 12, 97–101. [Google Scholar] [CrossRef]
- Katan, M.B.; Zock, P.; Mensink, R.P. Trans Fatty Acids and their Effects on Lipoproteins in Humans. Annu. Rev. Nutr. 1995, 15, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, E.E.; Kuo, T.M.; Kenty, M.M. Seed Protein and its Relationship to Soluble Sugars in Soybean. Crop. Sci. 1997, 37, 770–773. [Google Scholar] [CrossRef]
- Hymowitz, T.; Collins, F.I.; Panczner, J.; Walker, W.M. Relationship Between the Content of Oil, Protein, and Sugar in Soybean Seed 1. Agron. J. 1972, 64, 613–616. [Google Scholar] [CrossRef]
- Jiang, G.-L.; Chen, P.; Zhang, J.; Florez-Palacios, L.; Zeng, A.; Wang, X.; Bowen, R.A.; Miller, A.; Berry, H. Genetic Analysis of Sugar Composition and Its Relationship with Protein, Oil, and Fiber in Soybean. Crop. Sci. 2018, 58, 2413–2421. [Google Scholar] [CrossRef]
- Hou, A.; Chen, P.; Alloatti, J.; Li, D.; Mozzoni, L.; Zhang, B.; Shi, A. Genetic Variability of Seed Sugar Content in Worldwide Soybean Germplasm Collections. Crop. Sci. 2009, 49, 903–912. [Google Scholar] [CrossRef]
- Kim, E.-H.; Ro, H.-M.; Kim, S.-L.; Kim, H.-S.; Chung, I.-M. Analysis of Isoflavone, Phenolic, Soyasapogenol, and Tocopherol Compounds in Soybean [Glycine max (L.) Merrill] Germplasms of Different Seed Weights and Origins. J. Agric. Food Chem. 2012, 60, 6045–6055. [Google Scholar] [CrossRef]
- Tepavcevic, V.; Atanacković, M.; Miladinović, J.; Malenčić, D.; Popović, J.; Cvejić, J. Isoflavone Composition, Total Polyphenolic Content, and Antioxidant Activity in Soybeans of Different Origin. J. Med. Food 2010, 13, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chang, S.K.C. Characterization of Phenolic Substances and Antioxidant Properties of Food Soybeans Grown in the North Dakota−Minnesota Region. J. Agric. Food Chem. 2008, 56, 9102–9113. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Rani, A.; Dixit, A.K.; Pratap, D.; Bhatnagar, D. A comparative assessment of total phenolic content, ferric reducing-anti-oxidative power, free radical-scavenging activity, vitamin C and isoflavones content in soybean with varying seed coat colour. Food Res. Int. 2010, 43, 323–328. [Google Scholar] [CrossRef]
- Lee, S.-J.; Seguin, P.; Kim, J.-J.; Moon, H.-I.; Ro, H.-M.; Kim, E.-H.; Seo, S.-H.; Kang, E.-Y.; Ahn, J.-K.; Chung, I.-M. Isoflavones in Korean soybeans differing in seed coat and cotyledon color. J. Food Compos. Anal. 2010, 23, 160–165. [Google Scholar] [CrossRef]
- Wu, H.-J.; Deng, J.-C.; Yang, C.-Q.; Zhang, J.; Wang, X.-C.; Yang, F.; Yang, W.-Y.; Liu, J. Metabolite profiling of isoflavones and anthocyanins in black soybean [Glycine max (L.) Merr.] seeds by HPLC-MS and geographical differentiation analysis in Southwest China. Anal. Methods 2016, 9, 792–802. [Google Scholar] [CrossRef]
- Hoeck, J.A.; Fehr, W.R.; Murphy, P.A.; Welke, G.A. Influence of Genotype and Environment on Isoflavone Contents of Soybean. Crop. Sci. 2000, 40, 48–51. [Google Scholar] [CrossRef]
- Zhang, R.F.; Zhang, F.X.; Zhang, M.W.; Wei, Z.C.; Yang, C.Y.; Zhang, Y.; Tang, X.J.; Deng, Y.Y.; Chi, J.W. Phenolic Composition and Antioxidant Activity in Seed Coats of 60 Chinese Black Soybean (Glycine max L. Merr.) Varieties. J. Agric. Food Chem. 2011, 59, 5935–5944. [Google Scholar] [CrossRef]
- Monma, M.; Ito, M.; Saito, M.; Chikuni, K. Carotenoid Components in Soybean Seeds Varying with Seed Color and Maturation Stage. Biosci. Biotechnol. Biochem. 1994, 58, 926–930. [Google Scholar] [CrossRef]
- Kanamaru, K.; Wang, S.; Abe, J.; Yamada, T.; Kitamura, K. Identification and Characterization of Wild Soybean (Glycine soja Sieb. et Zecc.) Strains with High Lutein Content. Breed. Sci. 2006, 56, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-M.; Lim, H.; Kim, H.-S.; Lee, J.-S. Lutein Contents of Soybeans (Glycine max L.) Cultivated in Korea. Korean J. Food Sci. Technol. 2007, 39, 580–583. [Google Scholar]
- Seguin, P.; Tremblay, G.; Pageau, D.; Liu, W.; Turcotte, P. Soybean Lutein Concentration: Impact of Crop Management and Genotypes. Crop. Sci. 2011, 51, 1151–1160. [Google Scholar] [CrossRef]
- Mourtzinis, S.; Gaspar, A.P.; Naeve, S.L.; Conley, S. Planting Date, Maturity, and Temperature Effects on Soybean Seed Yield and Composition. Agron. J. 2017, 109, 2040–2049. [Google Scholar] [CrossRef] [Green Version]
- Primomo, V.S.; Falk, D.E.; Ablett, G.R.; Tanner, J.W.; Rajcan, I. Genotype × Environment Interactions, Stability, and Agronomic Performance of Soybean with Altered Fatty Acid Profiles. Crop. Sci. 2002, 42, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Je, J.-Y.; Park, P.-J.; Jung, W.-K.; Kim, S.-K. Amino acid changes in fermented oyster (Crassostrea gigas) sauce with different fermentation periods. Food Chem. 2005, 91, 15–18. [Google Scholar] [CrossRef]
- Kim, G.-H.; Hwang, Y.-S.; Ahn, K.-G.; Kim, G.-P.; Kim, M.-J.; Hong, S.-B.; Moon, J.-K.; Choung, M.-G. Determination of Soluble Carbohydrates in Soybean Seeds Using High Performance Liquid Chromatography with Evaporative Light Scattering Detection. J. Korean Soc. Food Sci. Nutr. 2014, 43, 1062–1067. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, K.H.; Lee, M.-H.; Kim, H.-T.; Seo, W.D.; Kim, J.Y.; Baek, I.-Y.; Jang, D.S.; Ha, T.J. Identification, characterisation, and quantification of phenolic compounds in the antioxidant activity-containing fraction from the seeds of Korean perilla (Perilla frutescens) cultivars. Food Chem. 2013, 136, 843–852. [Google Scholar] [CrossRef]
- Celli, G.B.; Pereira-Netto, A.B.; Beta, T. Comparative analysis of total phenolic content, antioxidant activity, and flavonoids profile of fruits from two varieties of Brazilian cherry (Eugenia uniflora L.) throughout the fruit developmental stages. Food Res. Int. 2011, 44, 2442–2451. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SN | Genotype | Seed Coat Color | Cotyledon Color | HSW (g) | Status | Days to Maturity (Category) |
|---|---|---|---|---|---|---|
| 1 | Cheongja2 | Black | Green | 26.0 | Cultivar | 108 (Early) |
| 2 | Cheongja3 | Black | Green | 32.1 | Cultivar | 117 (Mid) |
| 3 | Cheongja4 | Black | Green | 30.1 | Cultivar | 126 (Late) |
| 4 | Cheongja5 | Black | Green | 37.0 | Cultivar | 131 (Extremely late) |
| 5 | Cheongjakong | Black | Green | 30.4 | Cultivar | 111 (Mid) |
| 6 | Cheongyeob1 | Black | Yellow | 35.8 | Cultivar | 117 Mid |
| 7 | Daeheug | Black | Yellow | 34.3 | Cultivar | 108 (Early) |
| 8 | Geomjeongkong1 | Black | Yellow | 29.8 | Cultivar | 107 (Early) |
| 9 | Geomjeongkong2 | Black | Yellow | 28.3 | Cultivar | 112 (Mid) |
| 10 | Geomjeongkong3 | Black | Yellow | 31.0 | Cultivar | 111 (Mid) |
| 11 | Geomjeongkong4 | Black | Yellow | 28.0 | Cultivar | 107 (Early) |
| 12 | Geomjeongkong5 | Black | Yellow | 23.2 | Cultivar | 111 (Mid) |
| 13 | Heugmi | Black | Yellow | 24.8 | Cultivar | 105 (Early) |
| 14 | Heugsung | Black | Yellow | 29.2 | Cultivar | 110 (Early) |
| 15 | Ilpumgeomjeong2 | Black | Yellow | 25.0 | Cultivar | 109 (Early) |
| 16 | Ilpumgeomjeongkong | Black | Yellow | 28.0 | Cultivar | 102 (Early) |
| 17 | Jungmo3009 | Black | Green | 29.3 | Cultivar | 126 (Late) |
| 18 | Jungmo3011 | Black | Green | 66.4 | Cultivar | 155 (Extremely Late) |
| 19 | Seonheukkong | Black | Yellow | 34.2 | Cultivar | 112 (Mid) |
| 20 | Seoritae | Black | Green | 40.0 | Landrace | 148 (Extremely Late) |
| 21 | Socheong | Black | Green | 15.7 | Cultivar | 113 (Mid) |
| 22 | Socheong2 | Black | Green | 12.2 | Cultivar | 109 (Early) |
| 23 | Socheongja | Black | Green | 12.0 | Cultivar | 120 (Mid) |
| 24 | Taecheong | Black | Green | 44.5 | Cultivar | 127 (Late) |
| 25 | Tawonkong | Black | Yellow | 9.4 | Cultivar | 105 (Early) |
| 26 | Wonheug | Black | Yellow | 8.8 | Cultivar | 111 (Mid) |
| 27 | Jinyul | Brown | Yellow | 28.3 | Cultivar | 107 (Early) |
| 28 | Chungdu1 | Green | Green | 23.6 | Cultivar | 113 (Mid) |
| 29 | Jungmo3005 | Green | Green | 24.3 | Cultivar | 114 (Mid) |
| Palmitic | Stearic | Oleic | Linoleic | Linolenic | |
|---|---|---|---|---|---|
| Palmitic | 1 | ||||
| Stearic | 0.361 ** | 1 | |||
| Oleic | −0.299 * | −0.411 ** | 1 | ||
| Linoleic | 0.152 | 0.347 ** | −0.973 *** | 1 | |
| Linolenic | 0.100 | 0.077 | −0.713 *** | 0.595 *** | 1 |
| Protein | Oil | Stachyose | Raffinose | Sucrose | Glucose | Galactose | Fructose | Total Sugar | |
|---|---|---|---|---|---|---|---|---|---|
| Protein | 1 | ||||||||
| Oil | −0.525 **** | 1 | |||||||
| Stachyose | −0.235 ** | −0.151 | 1 | ||||||
| Raffinose | 0.213 ** | −0.279 * | −0.103 | 1 | |||||
| Sucrose | −0.034 | 0.016 | 0.125 | 0.4367 **** | 1 | ||||
| Glucose | 0.132 | 0.048 | −0.487 **** | 0.274 *** | 0.297 **** | 1 | |||
| Galactose | −0.126 | −0.068 | −0.082 | 0.282 *** | 0.088 | −0.131 | 1 | ||
| Fructose | −0.106 | 0.072 | −0.399 *** | −0.232 | −0.282 ** | 0.531 **** | 0.050 | 1 | |
| Total sugar | −0.074 | −0.036 | 0.318 **** | 0.461 **** | 0.967 **** | 0.272 *** | 0.120 | −0.239 * | 1 |
| ABTS | DPPH | Flavonoid | Polyphenol | |
|---|---|---|---|---|
| ABTS | 1 | |||
| DPPH | 0.962 **** | 1 | ||
| Flavonoid | 0.945 **** | 0.932 **** | 1 | |
| Polyphenol | 0.953 **** | 0.896 **** | 0.924 **** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhungana, S.K.; Seo, J.-H.; Kang, B.-K.; Park, J.-H.; Kim, J.-H.; Sung, J.-S.; Baek, I.-Y.; Shin, S.-O.; Jung, C.-S. Protein, Amino Acid, Oil, Fatty Acid, Sugar, Anthocyanin, Isoflavone, Lutein, and Antioxidant Variations in Colored Seed-Coated Soybeans. Plants 2021, 10, 1765. https://doi.org/10.3390/plants10091765
Dhungana SK, Seo J-H, Kang B-K, Park J-H, Kim J-H, Sung J-S, Baek I-Y, Shin S-O, Jung C-S. Protein, Amino Acid, Oil, Fatty Acid, Sugar, Anthocyanin, Isoflavone, Lutein, and Antioxidant Variations in Colored Seed-Coated Soybeans. Plants. 2021; 10(9):1765. https://doi.org/10.3390/plants10091765
Chicago/Turabian StyleDhungana, Sanjeev Kumar, Jeong-Hyun Seo, Beom-Kyu Kang, Ji-Hee Park, Jun-Hoi Kim, Jung-Sook Sung, In-Youl Baek, Sang-Ouk Shin, and Chan-Sik Jung. 2021. "Protein, Amino Acid, Oil, Fatty Acid, Sugar, Anthocyanin, Isoflavone, Lutein, and Antioxidant Variations in Colored Seed-Coated Soybeans" Plants 10, no. 9: 1765. https://doi.org/10.3390/plants10091765