
Evaluation of Frost Damage and Pod Set in Faba Bean (Vicia faba L.) under Field Conditions

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Materials
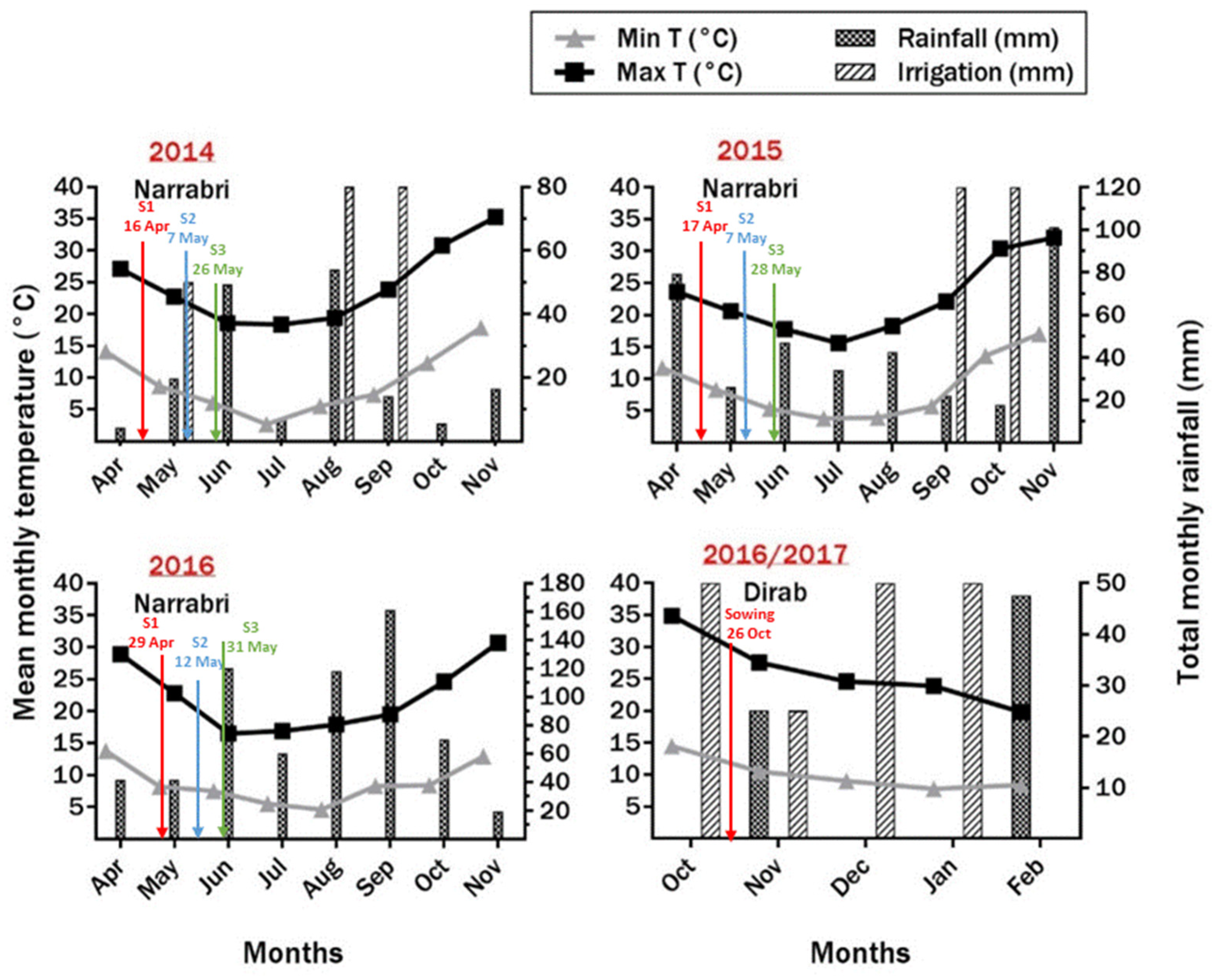
2.2. Environmental Conditions
2.3. Data Collection
2.3.1. The Field Experiments from 2014 to 2016 at Narrabri
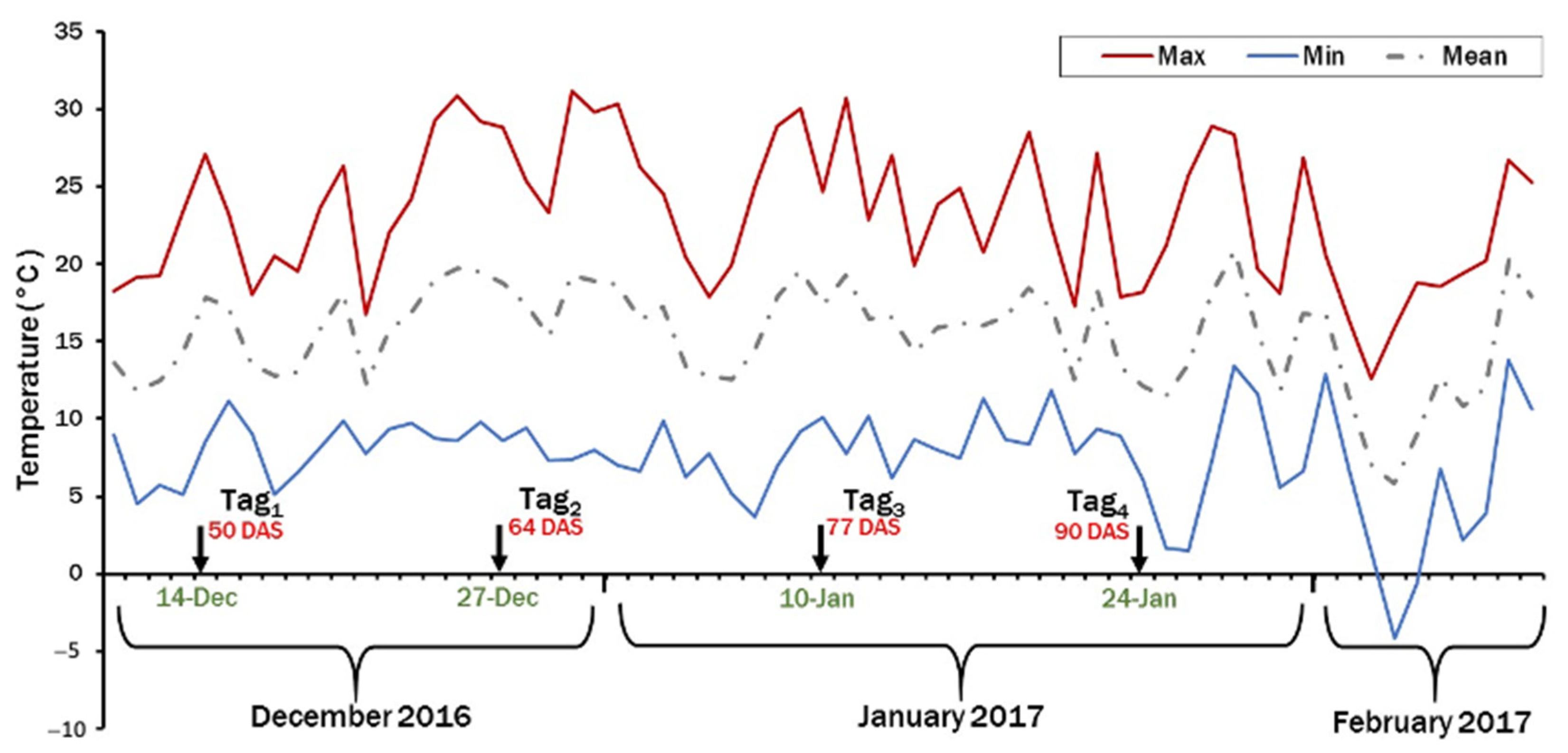
2.3.2. The Field Experiment in 2016/2017 at Dirab (Saudi Arabia)
2.4. Statistical Analysis
3. Results
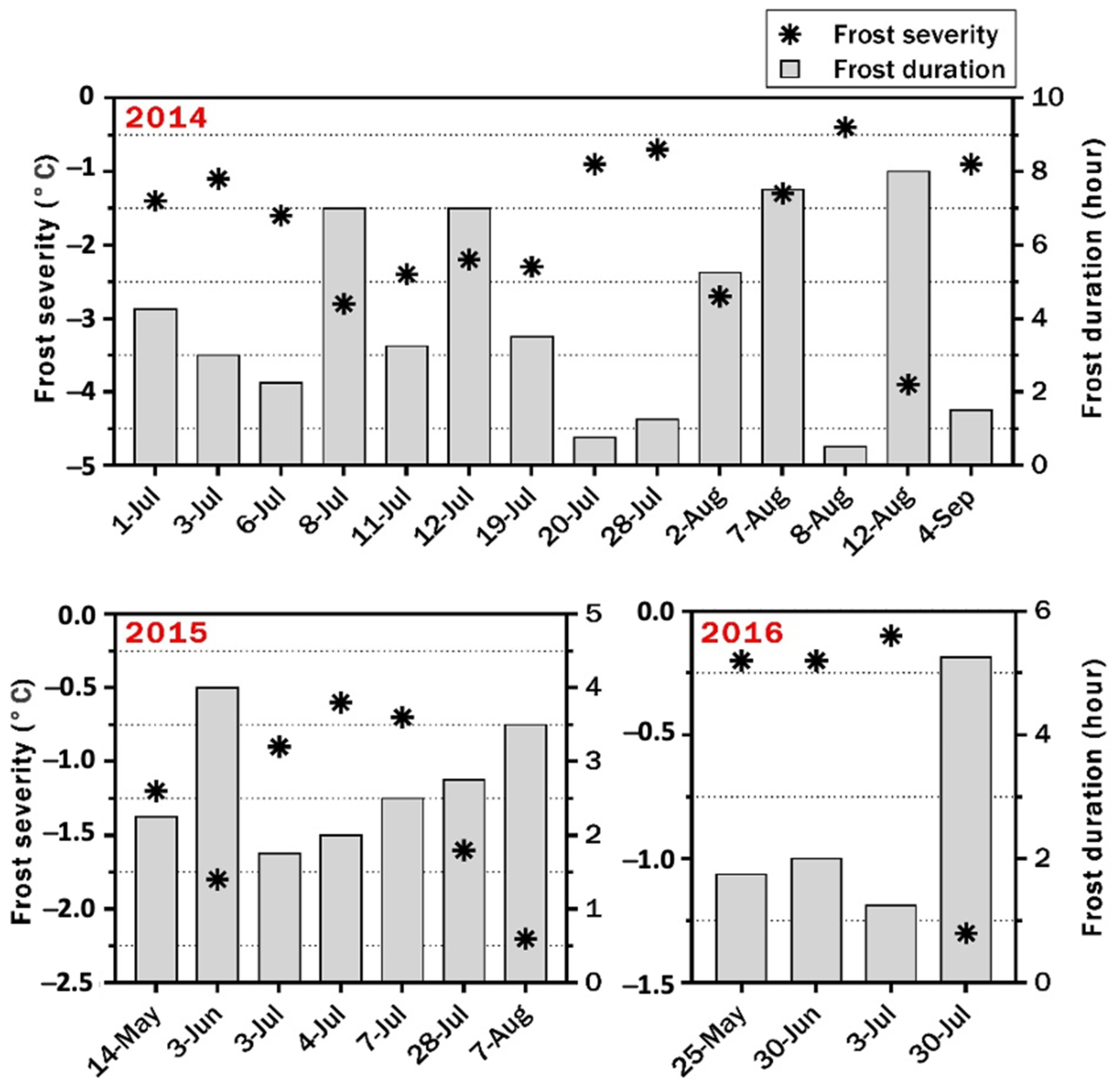
3.1. Response of Genotypes to Frost in 2014 at Narrabri
3.2. Frost Damage in 2015 at Narrabri
3.2.1. The Number of Damaged Plants at the Seedling Stage
3.2.2. A Visual 1–9 Score on the Vegetative and Reproductive Organs
3.2.3. Frost Damage on Flowers
3.3. Frost Damage in 2016 at Narrabri
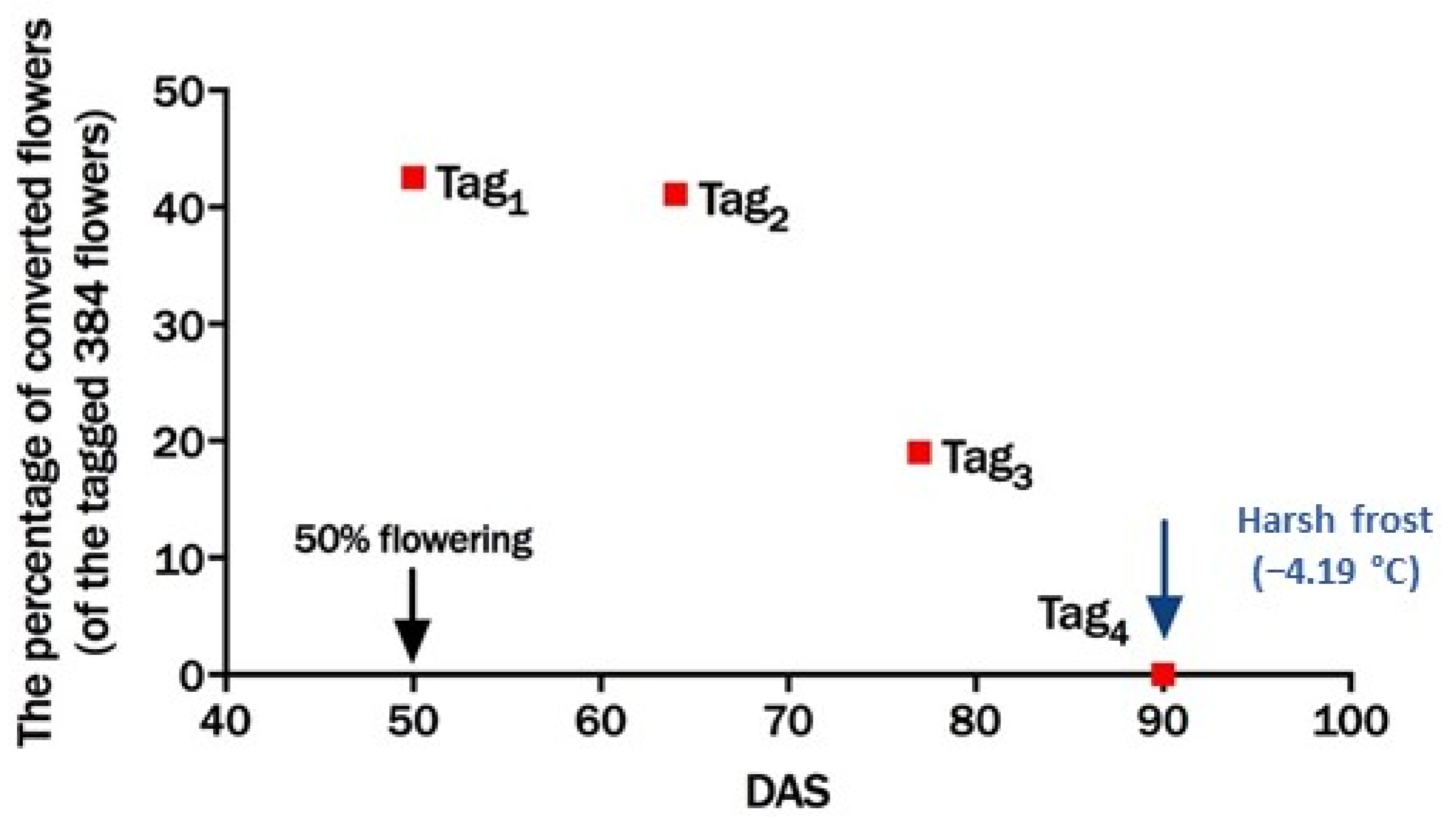
3.4. Frost Damage and Pod Set in 2016/2017 at Dirab (Saudi Arabia)
3.4.1. Response to Frost
3.4.2. Correlation of Frost Tolerance with Phenology and Grain Yield in 2015, 2016, and 2016/2017
4. Discussion
4.1. Effect of Frost Damage in 2014−2016 at Narrabri, and in 2016/2017 at Dirab
4.2. The Effect of Low Temperatures on Pod Set
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henriquez, B.; Olson, M.; Hoy, C.; Jackson, M.; Wouda, T. Frost tolerance of faba bean cultivars (Vicia faba L.) in central Alberta. Can. J. Plant Sci. 2017, 98, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Redden, R.; Paull, J.; Zong, X.; Sass, O.; Yang, T.; Ling, L. Faba Bean. In Broadening the Genetic Base of Grain Legumes; Singh, M., Bisht, I.S., Dutta, M., Eds.; Springer: New Delhi, India, 2014; pp. 75–93. [Google Scholar]
- Sallam, A.; Arbaoui, M.; El-Esawi, M.; Abshire, N.; Martsch, R. Identification and verification of QTL associated with frost tolerance using linkage mapping and GWAS in winter Faba bean. Front. Plant Sci. 2016, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crimp, S.J.; Zheng, B.; Khimashia, N.; Gobbett, D.L.; Chapman, S.; Howden, M.; Nicholls, N. Recent changes in southern Australian frost occurrence: Implications for wheat production risk. Crop Pasture Sci. 2016, 67, 801–811. [Google Scholar] [CrossRef]
- Zheng, B.; Chapman, S.C.; Christopher, J.T.; Frederiks, T.M.; Chenu, K. Frost trends and their estimated impact on yield in the Australian wheatbelt. J. Exp. Bot. 2015, 66, 3611–3623. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.P.; Fuller, A.M.; Kaniouras, S.; Christophers, J.; Fredericks, T. The freezing characteristics of wheat at ear emergence. Eur. J. Agron. 2007, 26, 435–441. [Google Scholar] [CrossRef]
- Paulsen, G.M.; Heyne, E.G. Grain Production of Winter Wheat after Spring Freeze Injury. Agron. J. 1983, 75, 705–707. [Google Scholar] [CrossRef]
- Dalezios, N.R. Environmental Hazards Methodologies for Risk Assessment and Management. Water Intell. Online 2017, 16. [Google Scholar] [CrossRef]
- Snyder, R.; Abreu, J.M. Frost Protection: Fundamentals, Practice and Economics; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2005. [Google Scholar]
- Tshewang, S.; JeSSop, R.; BiRChall, C. Effect of frost on triticale and wheat varieties at flowering in the North Eastern Australian cereal belt. Cereal Res. Commun. 2017, 45, 655–664. [Google Scholar] [CrossRef]
- Hamdi, M.R.; Alaban, M.A.; Jaber, M. Statistical Examination of Frost Characterization: A Case of Global Warming Impact in Jordan. J. Water Resour. Prot. 2011, 3, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Teitel, M.; Peiper, U.M.; Zvieli, Y. Shading screens for frost protection. Agric. For. Meteorol. 1996, 81, 273–286. [Google Scholar] [CrossRef]
- De Melo-Abreu, J.P.; Villalobos, F.J.; Mateos, L. Frost Protection. In Principles of Agronomy for Sustainable Agriculture; Villalobos, F.J., Fereres, E., Eds.; Springer: Cham, Switzerland, 2016; pp. 443–457. [Google Scholar]
- Maqbool, A.; Shafiq, S.; Lake, L. Radiant frost tolerance in pulse crops-a review. Euphytica 2010, 172, 1–12. [Google Scholar] [CrossRef]
- Link, W.; Bond, D. Resistance to freezing in winter faba beans. Grain Legum. 2011, 56, 19–20. [Google Scholar]
- Siddique, K.H.M.; Loss, S.P.; Regan, K.L.; Jettner, R.L. Adaptation and seed yield of cool season grain legumes in Mediterranean environments of south-western Australia. Aust. J. Agric. Res. 1999, 50, 375–387. [Google Scholar] [CrossRef]
- Gnanasambandam, A.; Paull, J.; Torres, A.; Kaur, S.; Leonforte, T.; Li, H.; Zong, X.; Yang, T.; Materne, M. Impact of molecular technologies on faba bean (Vicia faba L.) breeding strategies. Agronomy 2012, 2, 132–166. [Google Scholar] [CrossRef] [Green Version]
- Arbaoui, M.; Link, W. Effect of hardening on frost tolerance and fatty acid composition of leaves and stems of a set of faba bean (Vicia faba L.) genotypes. Euphytica 2008, 162, 211–219. [Google Scholar] [CrossRef]
- Toker, C.; Lluch, C.; Tejera, N.; Serraj, R.; Siddique, K. Abiotic stresses. In Chickpea Breeding and Management; Yadav, S.S., Redden, R.J., Chen, W., Sharma, B., Eds.; CABI: Egham, UK, 2007; pp. 474–496. [Google Scholar]
- Bhandari, K.; Sharma, K.D.; Hanumantha Rao, B.; Siddique, K.H.M.; Gaur, P.; Agrawal, S.K.; Nair, R.M.; Nayyar, H. Temperature sensitivity of food legumes: A physiological insight. Acta Physiol. Plant. 2017, 39, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Cook, S. Using Pollen Cues in Resource Location by a Pollinator and a Pest. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2001. [Google Scholar]
- Macfarlane, R.; Patten, K.; Royce, L.; Wyatt, B. Applied pollination in temperate areas. In Pollination of Cultivated Plants in the Tropics; Roubik, D.W., Ed.; Food & Agriculture Org.: Rome, Italy, 1995; pp. 20–39. [Google Scholar]
- Clarke, H.; Siddique, K. Growth and development. In The Chickpea Book: A Technical Guide to Chickpea Production; Bulletin 1326; Department of Agriculture of Western Australia: Perth, WA, Australia, 1998; pp. 3–10. [Google Scholar]
- Rylott, P.D.; Smith, M.L. Effects of applied plant growth substances on pod set in broad beans (Vicia faba var. major). J. Agric. Sci. 1990, 114, 41–47. [Google Scholar] [CrossRef]
- Sparks, D.L. Advances in Agronomy; Academic Press: New York, NY, USA, 2004. [Google Scholar]
- Ziena, H.M.; El Shehata, A.M.T.; Youssef, M.M. The effect of pod and seed positions on the physical and cooking properties of faba beans (Vicia faba). J. Sci. Food Agric. 1984, 35, 207–211. [Google Scholar] [CrossRef]
- Togun, A.O.; Tayo, T.O. Flowering and pod and seed development in pigeon pea (Cajanus cajan). J. Agric. Sci. 1990, 115, 327–335. [Google Scholar] [CrossRef]
- Eser, D.; Ukur, A.; Adak, M.S. Effect of seed size on yield and yield components in chickpea. Int. Chickpea Newsl. 1991, 25, 13–15. [Google Scholar]
- Matthews, P.; Jenkins, L. Winter Crop Variety Sowing Guide 2020. 2020. Available online: https://grdc.com.au/__data/assets/pdf_file/0016/400048/FINAL-WCVSG-2020-web.pdf?utm_source=website&utm_medium=download_button&utm_campaign=pdf_download&utm_term=North&utm_content=NSW%20Winter%20Crop (accessed on 16 March 2021).
- Sallam, A.; Ghanbari, M.; Martsch, R. Genetic analysis of winter hardiness and effect of sowing date on yield traits in winter faba bean. Sci. Hortic. 2017, 224, 296–301. [Google Scholar] [CrossRef]
- Sallam, A.; Martsch, R.; Moursi, Y.S. Genetic variation in morpho-physiological traits associated with frost tolerance in faba bean (Vicia faba L.). Euphytica 2015, 205, 395–408. [Google Scholar] [CrossRef]
- Murray, G.A.; Eser, D.; Gusta, L.V.; Eteve, G. Winterhardiness in pea, lentil, faba bean and chickpea. In World Crops: Cool Season Food Legumes; Summerfield, R.J., Ed.; Springer: Dordrecht, The Netherlands, 1988; pp. 831–843. [Google Scholar]
- Liu, Z.; Zhao, Y.; Bao, S.; Service, G.W. Low temperature and faba bean (Vicia faba L.) yield. Fabis Newsl. 1987, 18, 14–17. [Google Scholar]
- Rana, M. Vegetable Crops Science; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Mondal, M. A Study of Source-Sink Relation in Mungbean. Ph.D. Thesis, Department of Crop Botany, Bangladesh Agricultural University, Mymensingh, Bangladesh, 2007. [Google Scholar]
- Mondal, M.; Hamid, M. Flowering pattern and reproductive efficiencies in 30 groundnut mutants. Bangladesh J. Nucl. Agric. 1998, 14, 14–17. [Google Scholar]
- French, R.J. The contribution of pod numbers to field pea (Pisum sativum L.) yields in a short growing-season environment. Aust. J. Agric. Res. 1990, 41, 853–862. [Google Scholar] [CrossRef]
- Clifford, P.; Pentland, B.; Baylis, A. Reproductive abscission patterns in faba bean (Vicia faba L. cv. Troy). FABIS Newsl. 1990, 27, 5–9. [Google Scholar]
- Islam, M.; Rahim, M.; Prodhan, A. Flowering pattern, floral abscission and yield attributes in soybean influenced by GABA. J. Bangladesh Agric. Univ. 2010, 8, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Nahar, B.S.; Ikeda, T. Effect of silver-sheet and Figaron on flower production, abscission of reproductive organs, yield and yield components in soybean (Glycine max L.). J. Agron. Crop Sci. 2002, 188, 193–200. [Google Scholar] [CrossRef]
- Fakir, M. A Study on Morphophysiological Selection Criteria Related to Yield in Pigeonpea. Ph.D. Thesis, University of West Indies, St. Augustine, Trinidad, 1997. [Google Scholar]
- Martin, K.; Sauerborn, J. Crops and Their Environment. In Agroecology; Springer: Dordrecht, The Netherlands, 2013; pp. 103–185. [Google Scholar]
- Ohnishi, S.; Miyoshi, T.; Shirai, S. Low temperature stress at different flower developmental stages affects pollen development, pollination, and pod set in soybean. Environ. Exp. Bot. 2010, 69, 56–62. [Google Scholar] [CrossRef]
- Clarke, H.; Siddique, K.; Khan, T. Chickpea improvement in southern Australia: Breeding for tolerance to chilling at flowering. Indian J. Pulses Res. 2005, 18, 1–8. [Google Scholar]
- Hall, A.E.; Grantz, D.A. Drought Resistance of Cowpea Improved by Selecting for Early Appearance of Mature Pods. Crop Sci. 1981, 21, 461–464. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Narrabri | Dirab | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | 2015 | 2016 | 2016/2017 | ||||||||||||
| Frost Damage | Frost-E | Frost-S | Frost-F | Survival-F | Frost-S | Frost-F | Frost-S | Frost-P | |||||||
| Sowing Date | S I | S III | S I | S II | S I | S II | S I | S II | S I | S II | S I | S II | |||
| Genotypes | IX525c-1-10 | 9 | 11 | 6 | 8 | 6 | 7 | 23.1 | 13.7 | 5 | 2 | 3 | 2 | 4 | 5 |
| Fiord | 5 | 10 | 5 | 6 | 6 | 5 | 31 | 15.8 | 4 | 3 | 4 | 2 | 8 | 4 | |
| IX588d/1-4 | 5 | 19 | 3 | 3 | 3 | 5 | 19.8 | 5.8 | 1 | 2 | 3 | 1 | 7 | 6 | |
| Doza | 4 | 9 | 4 | 5 | 6 | 5 | 17.2 | 12.9 | 5 | 2 | 2 | 2 | 6 | 4 | |
| IX585c/1-11 | 4 | 35 | 5 | 5 | 6 | 7 | 26 | 9.4 | 2 | 2 | 1 | 2 | 8 | 4 | |
| Cairo | 3 | 12 | 4 | 6 | 4 | 4 | 25.8 | 5.2 | 4 | 2 | 3 | 2 | 7 | 5 | |
| 11NF010a-2 | 2 | 5 | 3 | 2 | 4 | 3 | 33.3 | 13.3 | 3 | 2 | 1 | 1 | 4 | 4 | |
| IX541a-2-8 | 2 | 10 | 7 | 7 | 6 | 7 | 10.4 | 10 | 6 | 3 | 3 | 4 | 7 | 4 | |
| PBA Warda | 2 | 10 | 4 | 5 | 4 | 4 | 8 | 11.5 | 4 | 2 | 3 | 2 | 3 | 2 | |
| 11NF014d-4 | 1 | 24 | 4 | 2 | 4 | 3 | 24.8 | 17.7 | 2 | 1 | 1 | 2 | 7 | 5 | |
| IX474/4-3 | 1 | 8 | 3 | 2 | 2 | 2 | 24.4 | 10.8 | 1 | 2 | 2 | 1 | 4 | 3 | |
| PBA Nanu | 1 | 12 | 4 | 4 | 6 | 6 | 9.7 | 9.9 | 3 | 3 | 3 | 3 | 6 | 4 | |
| IX559d-2-2 | 1 | 7 | 4 | 6 | 5 | 5 | 16.4 | 10.2 | 4 | 4 | 3 | 2 | 7 | 5 | |
| IX561f-4-2 | 0 | 12 | 4 | 5 | 4 | 5 | 20 | 1.9 | 3 | 2 | 2 | 1 | 7 | 4 | |
| PBA Nasma | 0 | 5 | 5 | 5 | 5 | 5 | 30.8 | 15.6 | 3 | 3 | 1 | 2 | 5 | 5 | |
| Giza Blanka | - | - | - | - | - | - | - | - | - | - | - | - | 7 | 5 | |
| Mean | 2.7 | 12.6 | 4.3 | 4.7 | 4.7 | 4.9 | 21.4 | 10.9 | 3.3 | 2.3 | 2.3 | 1.9 | 6.1 | 4.3 | |
| No. | Genotypes | Tag1 | Tag2 | Tag3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | IX588d/1-4 | 58.3 | A | a | 37.5 | B | bcd | 12.5 | C | cde |
| 2 | IX559d-2-2 | 56.5 | A | ab | 29.2 | B | cde | 25 | B | abc |
| 3 | IX525c-1-10 | 54.6 | A | ab | 45.8 | B | b | 29.2 | C | ab |
| 4 | 11NF010a-2 | 50 | A | abc | 37.5 | B | bcd | 16.7 | C | bcd |
| 5 | Cairo | 50 | A | abc | 41.7 | B | bc | 25 | C | abc |
| 6 | Fiord | 50 | A | abc | 50 | A | b | 16.7 | B | bcd |
| 7 | PBA Warda | 50 | A | abc | 41.7 | B | bc | 8.3 | C | de |
| 8 | IX585c/1-11 | 47.2 | B | abcd | 75 | A | a | 18.8 | C | bcd |
| 9 | 11NF014d-4 | 43.5 | B | bcde | 50 | A | b | 20.8 | C | abcd |
| 10 | IX541a-2-8 | 36.1 | A | cdef | 41.7 | A | bc | 20.8 | B | abcd |
| 11 | Doza | 35.2 | B | def | 45.8 | A | b | 12.5 | C | cde |
| 12 | IX561f-4-2 | 33.2 | A | def | 25 | B | de | 33.3 | A | a |
| 13 | PBA Nasma | 30.4 | B | ef | 41.7 | A | bc | 20.8 | C | abcd |
| 14 | Giza blanka | 29.2 | A | f | 20.8 | B | e | 0 | C | e |
| 15 | PBA Nanu | 28.9 | B | f | 37.5 | A | bcd | 25 | B | abc |
| 16 | IX474/4-3 | 26.7 | B | f | 37.5 | A | bcd | 18.8 | C | bcd |
| Mean | 42.5 | A | 41.1 | A | 19.0 | B | ||||
| No. | Genotypes | Tag1 | Tag2 | Tag3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | PBA Nasma | 16.7 | B | a | 15.0 | C | bcd | 17.3 | A | abc |
| 2 | IX561f-4-2 | 16.3 | A | ab | 15.3 | B | bcd | 15.3 | B | ef |
| 3 | PBA Nanu | 16.0 | A | abc | 15.3 | B | bcd | 15.7 | AB | def |
| 4 | 11NF014d-4 | 15.3 | B | bcd | 15.3 | B | bcd | 16.7 | A | bcd |
| 5 | PBA Warda | 15.3 | B | bcd | 15.0 | B | bcd | 17.0 | A | abc |
| 6 | 11NF010a-2 | 15.0 | B | cde | 15.7 | A | abc | 15.3 | AB | ef |
| 7 | IX541a-2-8 | 15.0 | A | cde | 15.0 | A | bcd | 15.0 | A | f |
| 8 | IX559d-2-2 | 14.7 | C | def | 16.7 | B | a | 18.0 | A | a |
| 9 | IX588d/1-4 | 14.7 | B | def | 14.3 | B | de | 17.0 | A | abc |
| 10 | Cairo | 14.3 | B | defg | 14.7 | B | cd | 16.3 | A | cde |
| 11 | Doza | 14.0 | B | efgh | 14.3 | B | de | 17.0 | A | abc |
| 12 | Giza blanka | 13.7 | C | fgh | 16.0 | B | ab | 16.9 | A | abc |
| 13 | IX474/4-3 | 13.5 | C | gh | 14.5 | B | de | 17.5 | A | ab |
| 14 | IX525c-1-10 | 13.3 | C | gh | 14.7 | B | cd | 15.7 | A | def |
| 15 | Fiord | 13.0 | C | h | 14.7 | B | cd | 17.3 | A | abc |
| 16 | IX585c/1-11 | 13.0 | B | h | 13.5 | B | e | 17.0 | A | abc |
| Mean | 14.6 | B | 15.0 | B | 16.6 | A | ||||
| 2015—Narrabri | Frost-E | Frost-S | Frost-F | Flowering | Maturity | SFD | Height | Yield | |
| Frost-E | - | ||||||||
| Frost-S | n.s. | - | |||||||
| Frost-F | n.s. | 0.5 *** | - | ||||||
| Flowering | 0.4 *** | n.s. | n.s. | - | |||||
| Maturity | −0.5 *** | n.s. | n.s. | −0.7 *** | - | ||||
| SFD | −0.5 *** | n.s. | n.s. | −0.9 *** | 0.9 *** | - | |||
| Height | −0.6 *** | n.s. | n.s. | −0.6 *** | 0.8 *** | 0.8 *** | - | ||
| Yield | −0.6 *** | −0.3** | n.s. | −0.6 *** | 0.7 *** | 0.7 *** | 0.6 *** | - | |
| 2016—Narrabri | Frost-S | Frost-F | Flowering | Maturity | SFD | Yield | |||
| Frost-S | - | ||||||||
| Frost-F | 0.3 ** | - | |||||||
| Flowering | n.s. | n.s. | - | ||||||
| Maturity | n.s. | n.s. | 0.5 *** | - | |||||
| SFD | n.s. | n.s. | −0.9 *** | n.s. | - | ||||
| Yield | −0.3 ** | n.s. | 0.3 ** | n.s. | −0.3 ** | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharbi, N.H.; Alghamdi, S.S.; Migdadi, H.M.; El-Harty, E.H.; Adhikari, K.N. Evaluation of Frost Damage and Pod Set in Faba Bean (Vicia faba L.) under Field Conditions. Plants 2021, 10, 1925. https://doi.org/10.3390/plants10091925
Alharbi NH, Alghamdi SS, Migdadi HM, El-Harty EH, Adhikari KN. Evaluation of Frost Damage and Pod Set in Faba Bean (Vicia faba L.) under Field Conditions. Plants. 2021; 10(9):1925. https://doi.org/10.3390/plants10091925
Chicago/Turabian StyleAlharbi, Najeeb H., Salem S. Alghamdi, Hussein M. Migdadi, Ehab H. El-Harty, and Kedar N. Adhikari. 2021. "Evaluation of Frost Damage and Pod Set in Faba Bean (Vicia faba L.) under Field Conditions" Plants 10, no. 9: 1925. https://doi.org/10.3390/plants10091925