
Genome-Wide Association Study of Leaf Rust Resistance at Seedling and Adult Plant Stages in a Global Barley Panel
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Screening for Seedling Resistance to P. hordei
2.3. Screening for Adult Plant Resistance (APR) to P. hordei
2.4. Phenotypic Data Analysis
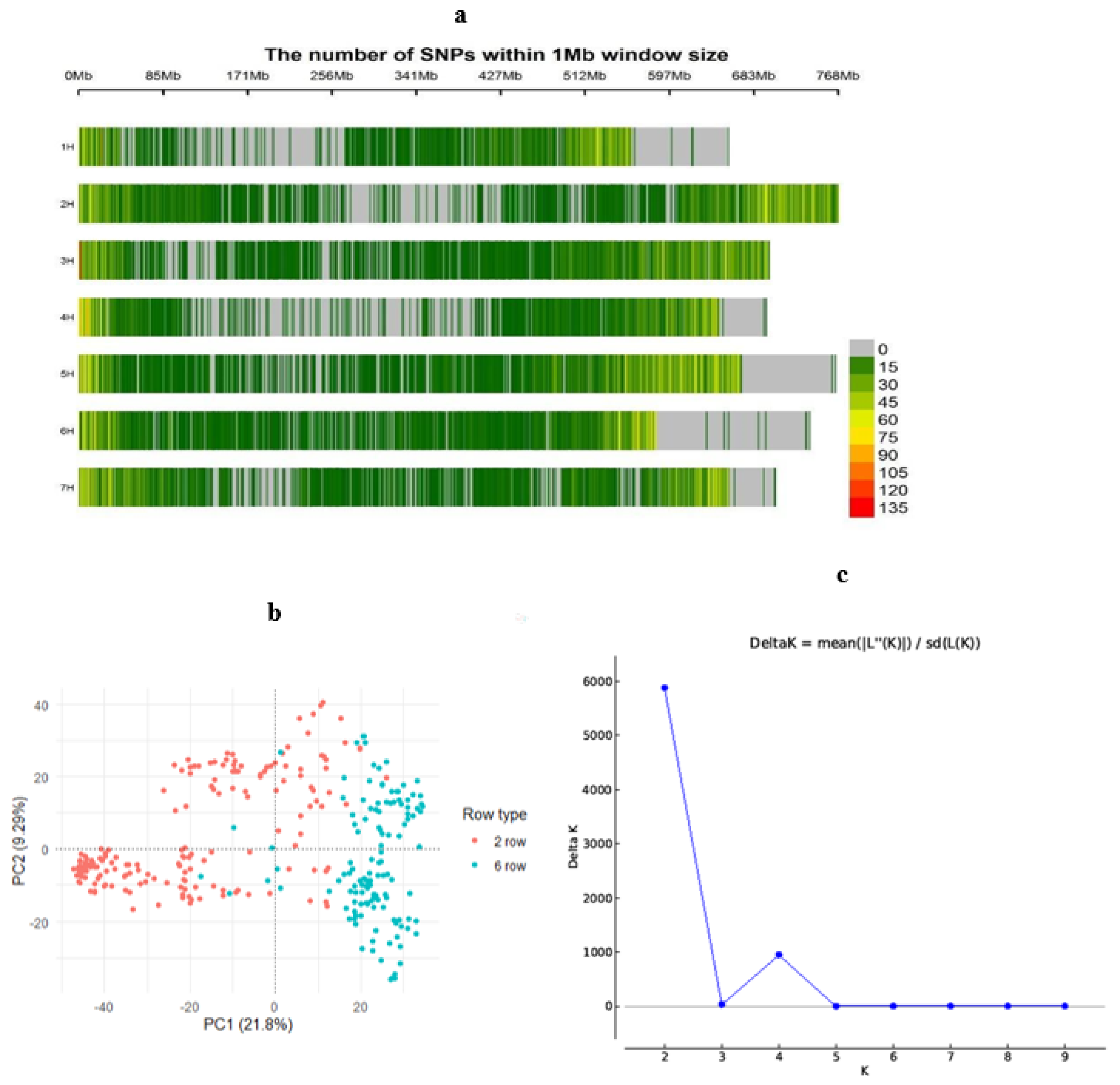
2.5. Genotyping and Population Structure
2.6. Genome-Wide Association Study (GWAS) of Leaf Rust Resistance
2.7. QTL Alignment and Candidate Genes
3. Results
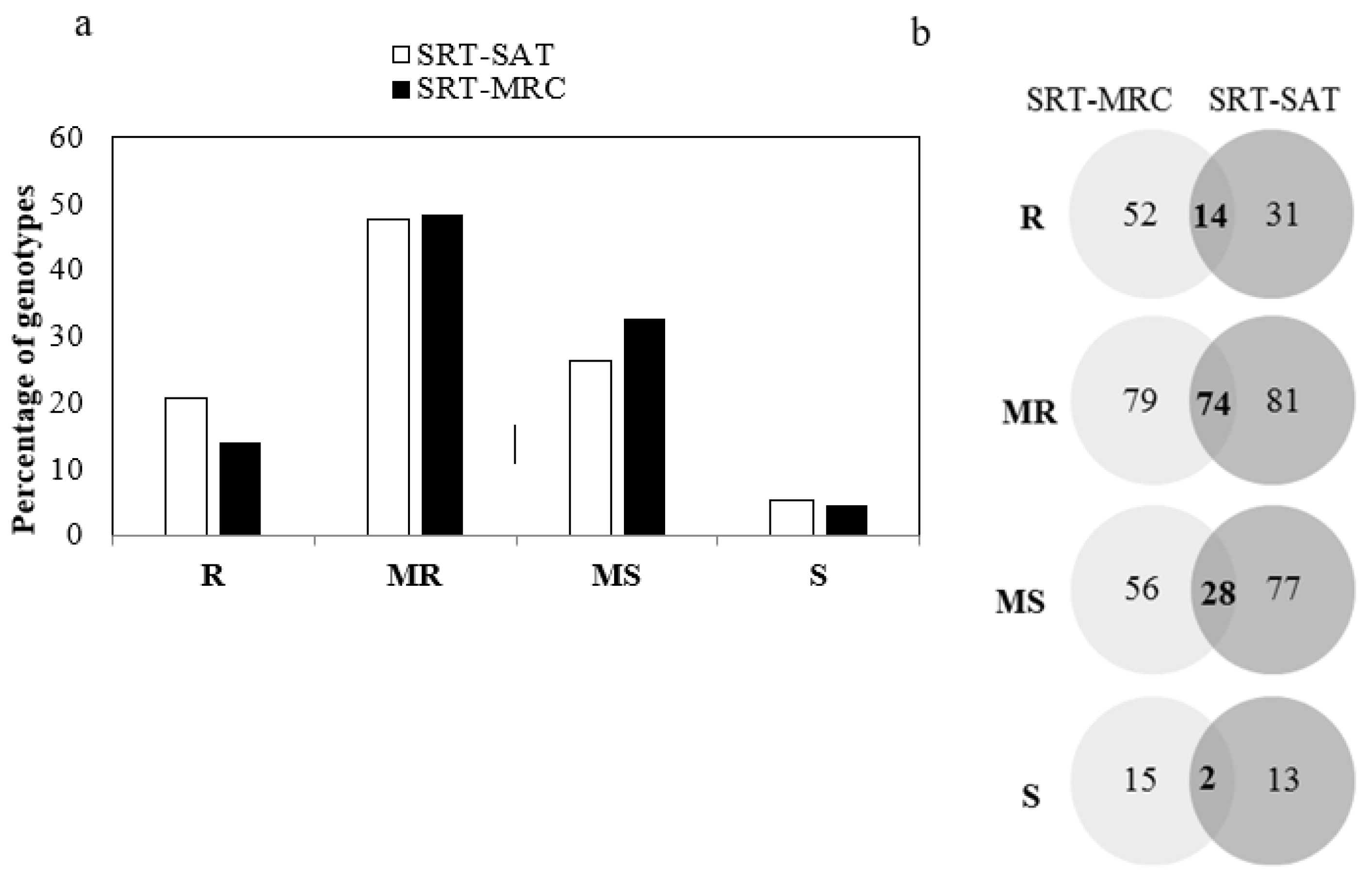
3.1. Seedling Resistance to P. hordei
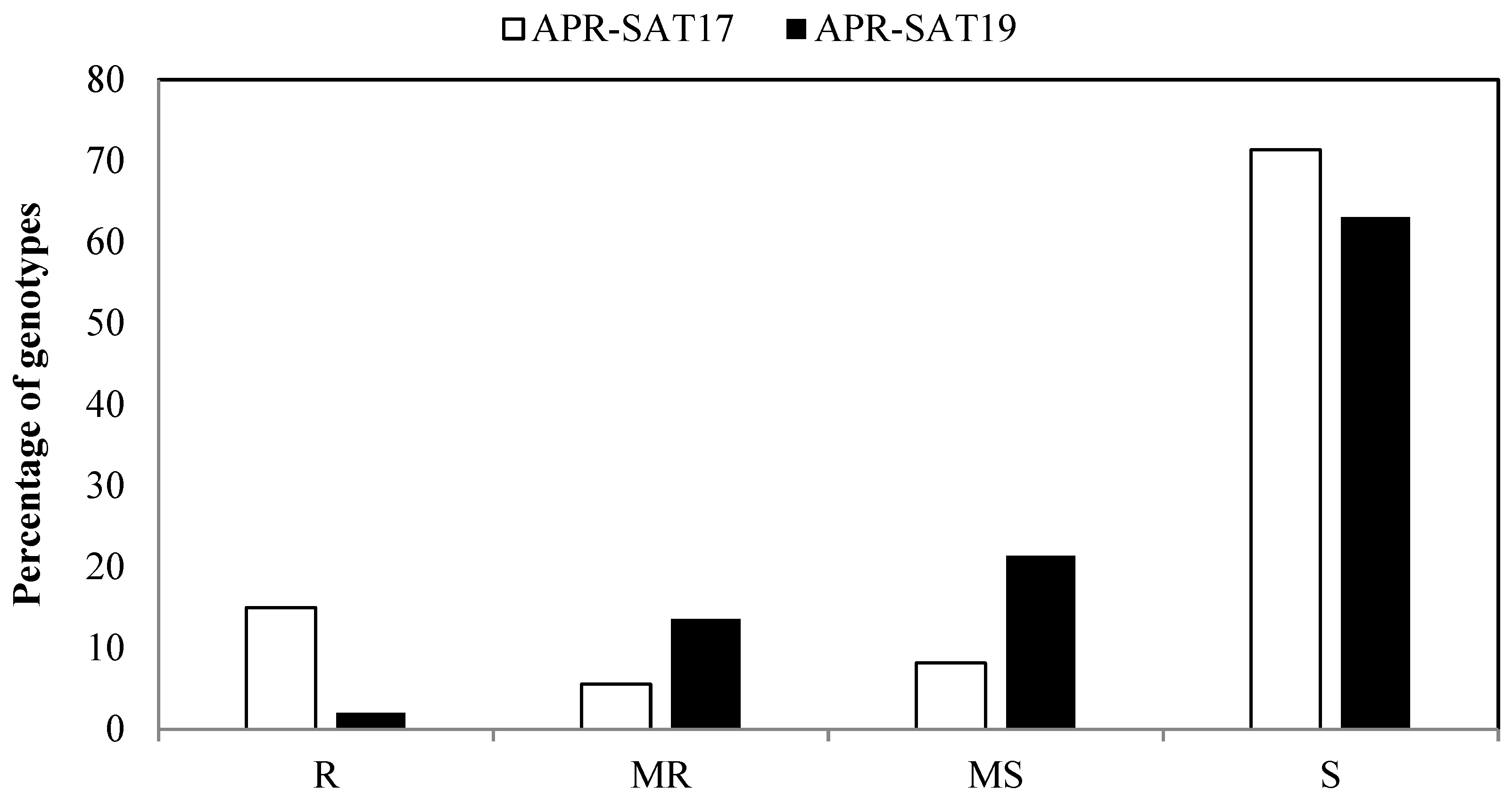
3.2. Phenotyping for Adult Plant Stage Resistance
3.3. Population Structure
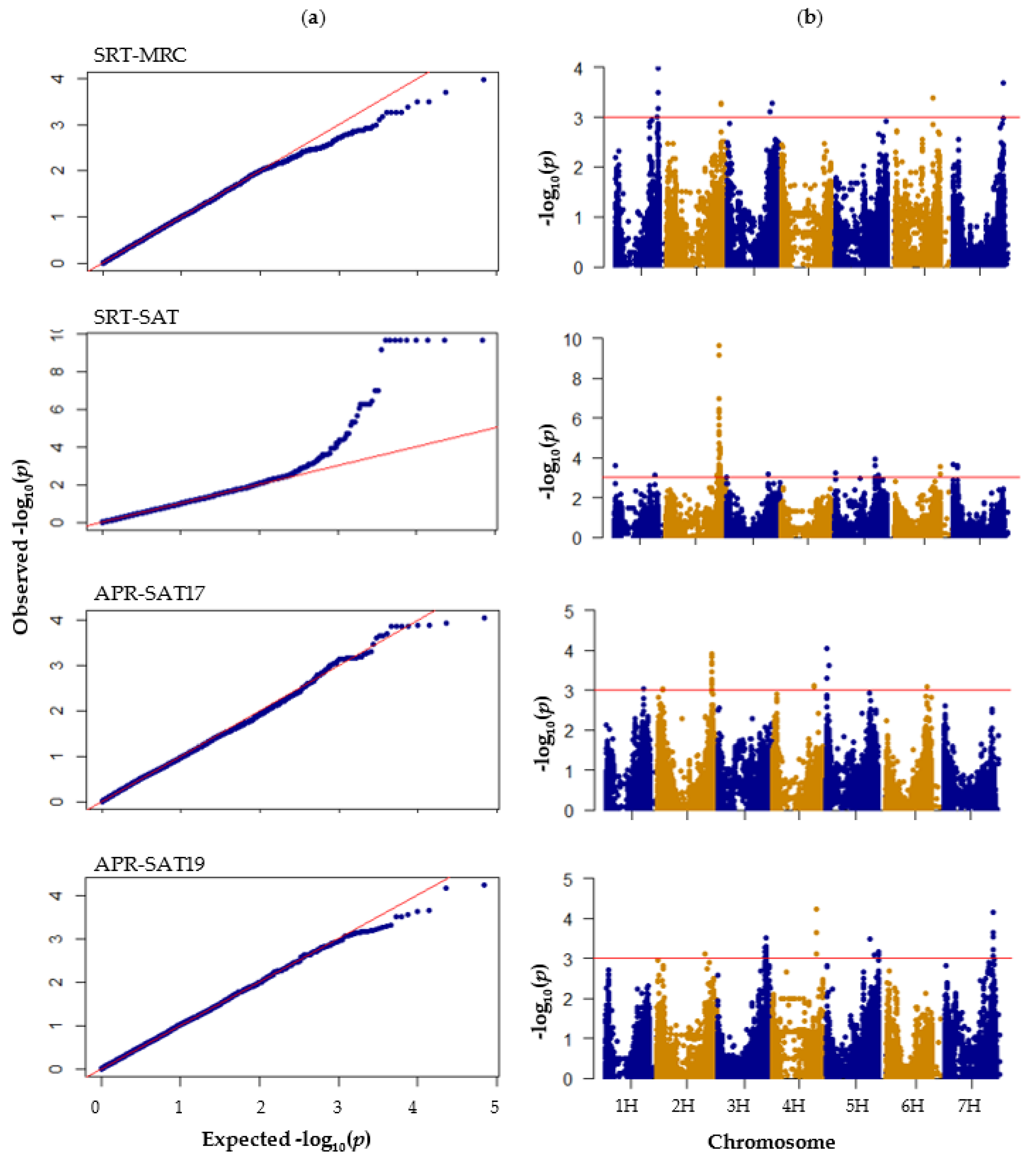
3.4. GWAS of Seedling Stage Resistance to P. hordei
3.5. GWAS of Adult Plant Stage Resistance to P. hordei
3.6. QTL Alignment and Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). 2020. Available online: http://www.fao.org/faostat/en/?#data/ (accessed on 17 November 2021).
- World Integrated Trade Solution (WITS). 2018. Available online: https://wits.worldbank.org (accessed on 4 August 2020).
- Park, R.F.; Golegaonkar, P.G.; Derevnina, L.; Sandhu, K.S.; Karaoglu, H.; Elmansour, H.M.; Dracatos, P.M.; Singh, D. Leaf rust of cultivated barley: Pathology and control. Annu. Rev. Phytopathol. 2015, 53, 565–589. [Google Scholar] [CrossRef]
- Clifford, B. Barley leaf rust. In Diseases, Distribution, Epidemiology, and Control; Elsevier: Amsterdam, The Netherlands, 1985; pp. 173–205. [Google Scholar]
- Chicaiza, O. Genetic Control of Leaf Rust in Barley; North Dakota State University: Fargo, ND, USA, 1996. [Google Scholar]
- Brunner, S.; Keller, B.; Feuillet, C. Molecular mapping of the Rph7. g leaf rust resistance gene in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2000, 101, 783–788. [Google Scholar] [CrossRef]
- Niks, R.; Walther, U.; Jaiser, H.; MartInez, F.; Rubiales, D. Resistance against barley leaf rust (Puccinia hordei) in West-European spring barley germplasm. Agronomie 2000, 20, 769–782. [Google Scholar] [CrossRef] [Green Version]
- Arnst, B.; Martens, J.; Wright, G.; Burnett, P.; Sanderson, F. Incidence, importance and virulence of Puccinia hordei on barley in New Zealand. Ann. Appl. Biol. 1979, 92, 185–190. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Parlevliet, J.; Lindhout, W.; Van Ommeren, A.; Kuiper, H. Level of partial resistance to leaf rust, Puccinia hordei, in West-European barley and how to select for it. Euphytica 1980, 29, 1–8. [Google Scholar] [CrossRef]
- Ballini, E.; Lauter, N.; Wise, R. Prospects for advancing defense to cereal rusts through genetical genomics. Front. Plant. Sci. 2013, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, W.L. Studies in the Inheritance of Resistance to Leaf Rust, Puccinia Anomola Rostr., in Crosses of Barley. I. J. Proc. R. Soc. New South Wales 1927, 218–247. [Google Scholar]
- Hickey, L.; Lawson, W.; Platz, G.; Dieters, M.; Arief, V.; German, S.; Fletcher, S.; Park, R.; Singh, D.; Pereyra, S. Mapping Rph20: A gene conferring adult plant resistance to Puccinia hordei in barley. Theor. Appl. Genet. 2011, 123, 55–68. [Google Scholar] [CrossRef]
- Singh, D.; Dracatos, P.; Derevnina, L.; Zhou, M.; Park, R.F.; Pillen, K. Rph23: A new designated additive adult plant resistance gene to leaf rust in barley on chromosome 7H. Plant Breed. 2015, 134, 62–69. [Google Scholar] [CrossRef]
- Ziems, L.A.; Hickey, L.T.; Platz, G.J.; Franckowiak, J.D.; Dracatos, P.M.; Singh, D.; Park, R.F. Characterization of Rph24: A gene conferring adult plant resistance to Puccinia hordei in barley. Phytopathology 2017, 107, 834–841. [Google Scholar] [CrossRef]
- Kavanagh, P.J.; Singh, D.; Bansal, U.K.; Park, R.F. Inheritance and characterization of the new and rare gene Rph25 conferring seedling resistance in Hordeum vulgare against Puccinia hordei. Plant Breed. 2017, 136, 908–912. [Google Scholar] [CrossRef]
- Yu, X.; Kong, H.Y.; Meiyalaghan, V.; Casonato, S.; Chng, S.; Jones, E.E.; Butler, R.C.; Pickering, R.; Johnston, P.A. Genetic mapping of a barley leaf rust resistance gene Rph26 introgressed from Hordeum bulbosum. Theor. Appl. Genet. 2018, 131, 2567–2580. [Google Scholar] [CrossRef]
- Rothwell, C.T.; Singh, D.; Dracatos, P.M.; Park, R.F. Inheritance and characterization of Rph27: A third race-specific resistance gene in the barley cultivar Quinn. Phytopathology 2020, 110, 1067–1073. [Google Scholar] [CrossRef]
- Mehnaz, M.; Dracatos, P.M.; Park, R.F.; Singh, D. Mining Middle Eastern and Central Asian Barley Germplasm to Understand Diversity for Resistance to Puccinia hordei, Causal Agent of Leaf Rust. Agronomy 2021, 11, 2146. [Google Scholar] [CrossRef]
- Burdon, J.J.; Barrett, L.G.; Rebetzke, G.; Thrall, P.H. Guiding deployment of resistance in cereals using evolutionary principles. Evol. Appl. 2014, 7, 609–624. [Google Scholar] [CrossRef]
- Niks, R.E.; Qi, X.; Marcel, T.C. Quantitative resistance to biotrophic filamentous plant pathogens: Concepts, misconceptions, and mechanisms. Annu. Rev. Phytopathol. 2015, 53, 445–470. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, L.; Germán, S.; Pereyra, S.; Hayes, P.M.; Pérez, C.A.; Capettini, F.; Locatelli, A.; Berberian, N.M.; Falconi, E.E.; Estrada, R. Multi-environment multi-QTL association mapping identifies disease resistance QTL in barley germplasm from Latin America. Theor. Appl. Genet. 2015, 128, 501–516. [Google Scholar] [CrossRef]
- Qi, X.; Niks, R.E.; Stam, P.; Lindhout, P. Identification of QTLs for partial resistance to leaf rust (Puccinia hordei) in barley. Theor. Appl. Genet. 1998, 96, 1205–1215. [Google Scholar] [CrossRef]
- Qi, X.; Fufa, F.; Sijtsma, D.; Niks, R.E.; Lindhout, P.; Stam, P. The evidence for abundance of QTLs for partial resistance to Puccinia hordei on the barley genome. Mol. Breed. 2000, 6, 1–9. [Google Scholar] [CrossRef]
- González, A.M.; Marcel, T.C.; Niks, R.E. Evidence for a minor gene–for–minor gene interaction explaining nonhypersensitive polygenic partial disease resistance. Phytopathology 2012, 102, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Marcel, T.C.; Varshney, R.; Barbieri, M.; Jafary, H.; De Kock, M.; Graner, A.; Niks, R. A high-density consensus map of barley to compare the distribution of QTLs for partial resistance to Puccinia hordei and of defence gene homologues. Theor. Appl. Genet. 2007, 114, 487–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Dracatos, P.; Loughman, R.; Park, R. Genetic mapping of resistance to Puccinia hordei in three barley doubled-haploid populations. Euphytica 2017, 213, 16. [Google Scholar] [CrossRef]
- Zhu, C.; Gore, M.; Buckler, E.S.; Yu, J. Status and prospects of association mapping in plants. Plant Genome 2008, 1, 5–20. [Google Scholar] [CrossRef]
- Yu, J.; Buckler, E.S. Genetic association mapping and genome organization of maize. Curr. Opin. Biotechnol. 2006, 17, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Bjørnstad, Å.; Xu, Y. The genetic dissection of quantitative traits in crops. Electron. J. Biotechnol. 2010, 13, 16–17. [Google Scholar] [CrossRef] [Green Version]
- Amezrou, R.; Verma, R.P.S.; Chao, S.; Brueggeman, R.S.; Belqadi, L.; Arbaoui, M.; Rehman, S.; Gyawali, S. Genome-wide association studies of net form of net blotch resistance at seedling and adult plant stages in spring barley collection. Mol. Breed. 2018, 38, 58. [Google Scholar] [CrossRef]
- Burlakoti, R.; Gyawali, S.; Chao, S.; Smith, K.; Horsley, R.; Cooper, B.; Muehlbauer, G.; Neate, S. Genome-wide association study of spot form of net blotch resistance in the Upper Midwest barley breeding programs. Phytopathology 2017, 107, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Tamang, P.; Neupane, A.; Mamidi, S.; Friesen, T.; Brueggeman, R. Association mapping of seedling resistance to spot form net blotch in a worldwide collection of barley. Phytopathology 2015, 105, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Visioni, A.; Rehman, S.; Viash, S.S.; Singh, S.P.; Vishwakarma, R.; Gyawali, S.; Al-Abdallat, A.M.; Verma, R.P.S. Genome wide association mapping of spot blotch resistance at seedling and adult plant stages in barley. Front. Plant Sci. 2020, 11, 642. [Google Scholar] [CrossRef]
- Ziems, L.; Hickey, L.; Hunt, C.; Mace, E.; Platz, G.; Franckowiak, J.; Jordan, D. Association mapping of resistance to Puccinia hordei in Australian barley breeding germplasm. Theor. Appl. Genet. 2014, 127, 1199–1212. [Google Scholar] [CrossRef] [PubMed]
- Turuspekov, Y.; Ormanbekova, D.; Rsaliev, A.; Abugalieva, S. Genome-wide association study on stem rust resistance in Kazakh spring barley lines. BMC Plant Biol. 2016, 16, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visioni, A.; Gyawali, S.; Selvakumar, R.; Gangwar, O.P.; Shekhawat, P.S.; Bhardwaj, S.C.; Al-Abdallat, A.M.; Kehel, Z.; Verma, R.P. Genome wide association mapping of seedling and adult plant resistance to barley stripe rust (Puccinia striiformis f. sp. hordei) in India. Front. Plant Sci. 2018, 9, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prins, R.; Dreisigacker, S.; Pretorius, Z.; Van Schalkwyk, H.; Wessels, E.; Smit, C.; Bender, C.; Singh, D.; Boyd, L.A. Stem rust resistance in a geographically diverse collection of spring wheat lines collected from across Africa. Front. Plant Sci. 2016, 7, 973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stakman, E.C.; Stewart, D.; Loegering, W.Q. Identification of Physiologic Races of Puccinia Graminis Var. Tritici; USDA: Washington, DC, USA, 1962.
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Peterson, R.F.; Campbell, A.; Hannah, A. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Stubbs, R.; Prescott, J.; Saari, E.; Dubin, H. Cereal Disease Methodology Manual; CIMMYT: Veracruz, Mexico, 1986; p. 46. [Google Scholar]
- Jeger, M.; Viljanen-Rollinson, S. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]
- Slotta, T.A.B.; Brady, L.; Chao, S. High throughput tissue preparation for large-scale genotyping experiments. Mol. Ecol. Resour. 2008, 8, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Bayer, M.M.; Rapazote-Flores, P.; Ganal, M.; Hedley, P.E.; Macaulay, M.; Plieske, J.; Ramsay, L.; Russell, J.; Shaw, P.D.; Thomas, W. Development and evaluation of a barley 50k iSelect SNP array. Front. Plant Sci. 2017, 8, 1792. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Yashveer, S.; Rehman, S.; Gyawali, S.; Kumar, Y.; Chao, S.; Sarker, A.; Verma, R.P.S. Genetic and Agro-morphological diversity in global barley (Hordeum vulgare L.) collection at ICARDA. Genet. Resour. Crop Evol. 2021, 68, 1315–1330. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Serrote, C.M.L.; Reiniger, L.R.S.; Silva, K.B.; dos Santos Rabaiolli, S.M.; Stefanel, C.M. Determining the polymorphism information content of a molecular marker. Gene 2020, 726, 144175. [Google Scholar] [CrossRef] [PubMed]
- Tsehay, S.; Ortiz, R.; Johansson, E.; Bekele, E.; Tesfaye, K.; Hammenhag, C.; Geleta, M. New Transcriptome-Based SNP Markers for Noug (Guizotia abyssinica) and Their Conversion to KASP Markers for Population Genetics Analyses. Genes 2020, 11, 1373. [Google Scholar] [CrossRef]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar]
- Storey, J.D. A direct approach to false discovery rates. J. R. Stat. Soc. Ser. B 2002, 64, 479–498. [Google Scholar] [CrossRef] [Green Version]
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Mao, Y.; Xie, C.; Smith, H.; Luo, L.; Xu, S. Mapping quantitative trait loci using naturally occurring genetic variance among commercial inbred lines of maize (Zea mays L.). Genetics 2005, 169, 2267–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Tian, F.; Pan, Y.; Buckler, E.S.; Zhang, Z. A SUPER powerful method for genome wide association study. PLoS ONE 2014, 9, e107684. [Google Scholar] [CrossRef] [PubMed]
- Segura, V.; Vilhjálmsson, B.J.; Platt, A.; Korte, A.; Seren, Ü.; Long, Q.; Nordborg, M. An efficient multi-locus mixed-model approach for genome-wide association studies in structured populations. Nat. Genet. 2012, 44, 825–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Ersoz, E.; Lai, C.-Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A.; Bradbury, P.J.; Yu, J.; Arnett, D.K.; Ordovas, J.M. Mixed linear model approach adapted for genome-wide association studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Liu, X.; Zhou, Y.; Summers, R.M.; Zhang, Z. BLINK: A package for the next level of genome-wide association studies with both individuals and markers in the millions. Gigascience 2019, 8, giy154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorrips, R.E. MapChart: Software for the Graphical Presentation of Linkage Maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantalapiedra, C.P.; Boudiar, R.; Casas, A.M.; Igartua, E.; Contreras-Moreira, B. BARLEYMAP: Physical and genetic mapping of nucleotide sequences and annotation of surrounding loci in barley. Mol. Breed. 2015, 35, 13. [Google Scholar] [CrossRef]
- International Barley Genome Sequencing Consortium (IBGSC). Available online: https://webblast.ipk-gatersleben.de/barley_ibsc/ (accessed on 2 October 2020).
- Ziems, L.; Franckowiak, J.; Platz, G.; Mace, E.; Park, R.; Singh, D.; Jordan, D.; Hickey, L. Investigating successive Australian barley breeding populations for stable resistance to leaf rust. Theor. Appl. Genet. 2017, 130, 2463–2477. [Google Scholar] [CrossRef]
- Druka, A.; Franckowiak, J.; Lundqvist, U.; Bonar, N.; Alexander, J.; Houston, K.; Radovic, S.; Shahinnia, F.; Vendramin, V.; Morgante, M. Genetic dissection of barley morphology and development. Plant Physiol. 2011, 155, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Hackett, C.A.; Niks, R.E.; Hedley, P.E.; Booth, C.; Druka, A.; Marcel, T.C.; Vels, A.; Bayer, M.; Milne, I. An eQTL analysis of partial resistance to Puccinia hordei in barley. PLoS ONE 2010, 5, e8598. [Google Scholar] [CrossRef] [Green Version]
- Berger, G.L.; Liu, S.; Hall, M.D.; Brooks, W.S.; Chao, S.; Muehlbauer, G.J.; Baik, B.-K.; Steffenson, B.; Griffey, C.A. Marker-trait associations in Virginia Tech winter barley identified using genome-wide mapping. Theor. Appl. Genet. 2013, 126, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Roane, C.; Starling, T. Inheritance of reaction to Puccinia hordei in barley. 2. Gene symbols for loci in differential cultivars. Phytopathology 1967, 57, 66–68. [Google Scholar]
- Von Korff, M.; Wang, H.; Léon, J.; Pillen, K. AB-QTL analysis in spring barley. I. Detection of resistance genes against powdery mildew, leaf rust and scald introgressed from wild barley. Theor. Appl. Genet. 2005, 111, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Borovkova, I.; Jin, Y.; Steffenson, B. Chromosomal location and genetic relationship of leaf rust resistance genes Rph9 and Rph12 in barley. Phytopathology 1998, 88, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Vatter, T.; Maurer, A.; Perovic, D.; Kopahnke, D.; Pillen, K.; Ordon, F. Identification of QTL conferring resistance to stripe rust (Puccinia striiformis f. sp. hordei) and leaf rust (Puccinia hordei) in barley using nested association mapping (NAM). PLoS ONE 2018, 13, e0191666. [Google Scholar] [CrossRef] [Green Version]
- Backes, G.; Madsen, L.; Jaiser, H.; Stougaard, J.; Herz, M.; Mohler, V.; Jahoor, A. Localisation of genes for resistance against Blumeria graminis f. sp. hordei and Puccinia graminis in a cross between a barley cultivar and a wild barley (Hordeum vulgare ssp. spontaneum) line. Theor. Appl. Genet. 2003, 106, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Ivandic, V.; Walther, U.; Graner, A. Molecular mapping of a new gene in wild barley conferring complete resistance to leaf rust (Puccinia hordei Otth). Theor. Appl. Genet. 1998, 97, 1235–1239. [Google Scholar] [CrossRef]
- Schnaithmann, F.; Kopahnke, D.; Pillen, K. A first step toward the development of a barley NAM population and its utilization to detect QTLs conferring leaf rust seedling resistance. Theor. Appl. Genet. 2014, 127, 1513–1525. [Google Scholar] [CrossRef]
- Schweizer, P.; Stein, N. Large-scale data integration reveals colocalization of gene functional groups with meta-QTL for multiple disease resistance in barley. Mol. Plant-Microbe Interact. 2011, 24, 1492–1501. [Google Scholar] [CrossRef] [Green Version]
- Borovkova, I.; Steffenson, B.; Jin, Y.; Kilian, A.; Kleinhofs, A.; Blake, T. Identification and mapping of a leaf rust resistance gene in barley line Q21861. Genome 1997, 40, 236–241. [Google Scholar] [CrossRef]
- Castro, A.J.; Gamba, F.; German, S.; Gonzalez, S.; Hayes, P.M.; Pereyra, S.; Perez, C. Quantitative trait locus analysis of spot blotch and leaf rust resistance in the BCD47× Baronesse barley mapping population. Plant Breed. 2012, 131, 258–266. [Google Scholar] [CrossRef]
- Park, R.; Burdon, J.; McIntosh, R. Studies on the origin, spread, and evolution of an important group of Puccinia recondita f. sp. tritici pathotypes in Australasia. Eur. J. Plant Pathol. 1995, 101, 613–622. [Google Scholar] [CrossRef]
- Comadran, J.; Thomas, W.; Van Eeuwijk, F.; Ceccarelli, S.; Grando, S.; Stanca, A.; Pecchioni, N.; Akar, T.; Al-Yassin, A.; Benbelkacem, A. Patterns of genetic diversity and linkage disequilibrium in a highly structured Hordeum vulgare association-mapping population for the Mediterranean basin. Theor. Appl. Genet. 2009, 119, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Cockram, J.; White, J.; Leigh, F.J.; Lea, V.J.; Chiapparino, E.; Laurie, D.A.; Mackay, I.J.; Powell, W.; O'Sullivan, D.M. Association mapping of partitioning loci in barley. BMC Genet. 2008, 9, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massman, J.; Cooper, B.; Horsley, R.; Neate, S.; Dill-Macky, R.; Chao, S.; Dong, Y.; Schwarz, P.; Muehlbauer, G.; Smith, K. Genome-wide association mapping of Fusarium head blight resistance in contemporary barley breeding germplasm. Mol. Breed. 2011, 27, 439–454. [Google Scholar] [CrossRef]
- Pasam, R.K.; Sharma, R.; Malosetti, M.; van Eeuwijk, F.A.; Haseneyer, G.; Kilian, B.; Graner, A. Genome-wide association studies for agronomical traits in a world wide spring barley collection. BMC Plant Biol. 2012, 12, 16. [Google Scholar] [CrossRef] [Green Version]
- Dinh, H.X.; Singh, D.; Periyannan, S.; Park, R.F.; Pourkheirandish, M. Molecular genetics of leaf rust resistance in wheat and barley. Theor. Appl. Genet. 2020, 133, 2035–2050. [Google Scholar] [CrossRef]
- Remington, D.L.; Thornsberry, J.M.; Matsuoka, Y.; Wilson, L.M.; Whitt, S.R.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler, E.S. Structure of linkage disequilibrium and phenotypic associations in the maize genome. Proc. Natl. Acad. Sci. USA 2001, 98, 11479–11484. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Tör, M.; Brown, D.; Cooper, A.; Woods-Tör, A.; Sjölander, K.; Jones, J.D.; Holub, E.B. Arabidopsis downy mildew resistance gene RPP27 encodes a receptor-like protein similar to CLAVATA2 and tomato Cf-9. Plant Physiol. 2004, 135, 1100–1112. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.-Y.; Wang, G.-L.; Chen, L.-L.; Kim, H.-S.; Pi, L.-Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.-X.; Zhu, L.-H. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bent, A.F.; Kunkel, B.N.; Dahlbeck, D.; Brown, K.L.; Schmidt, R.; Giraudat, J.; Leung, J.; Staskawicz, B.J. RPS2 of Arabidopsis thaliana: A leucine-rich repeat class of plant disease resistance genes. Science 1994, 265, 1856–1860. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.R.; Godiard, L.; Straube, E.; Ashfield, T.; Lewald, J.; Sattler, A.; Innes, R.W.; Dangl, J.L. Structure of the Arabidopsis RPM1 gene enabling dual specificity disease resistance. Science 1995, 269, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Mindrinos, M.; Katagiri, F.; Yu, G.-L.; Ausubel, F.M. The A. thaliana disease resistance gene RPS2 encodes a protein containing a nucleotide-binding site and leucine-rich repeats. Cell 1994, 78, 1089–1099. [Google Scholar] [CrossRef]
- Warren, R.F.; Henk, A.; Mowery, P.; Holub, E.; Innes, R.W. A mutation within the leucine-rich repeat domain of the Arabidopsis disease resistance gene RPS5 partially suppresses multiple bacterial and downy mildew resistance genes. Plant Cell 1998, 10, 1439–1452. [Google Scholar] [CrossRef] [Green Version]
- Mohr, T.J.; Mammarella, N.D.; Hoff, T.; Woffenden, B.J.; Jelesko, J.G.; McDowell, J.M. The Arabidopsis downy mildew resistance gene RPP8 is induced by pathogens and salicylic acid and is regulated by W box cis elements. Mol. Plant-Microbe Interact. 2010, 23, 1303–1315. [Google Scholar] [CrossRef] [Green Version]
- Cloutier, S.; McCallum, B.D.; Loutre, C.; Banks, T.W.; Wicker, T.; Feuillet, C.; Keller, B.; Jordan, M.C. Leaf rust resistance gene Lr1, isolated from bread wheat (Triticum aestivum L.) is a member of the large psr567 gene family. Plant Mol. Biol. 2007, 65, 93–106. [Google Scholar] [CrossRef]
- Feuillet, C.; Travella, S.; Stein, N.; Albar, L.; Nublat, A.; Keller, B. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. Proc. Natl. Acad. Sci. USA 2003, 100, 15253–15258. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Brooks, S.A.; Li, W.; Fellers, J.P.; Trick, H.N.; Gill, B.S. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics 2003, 164, 655–664. [Google Scholar] [CrossRef]
- Thind, A.K.; Wicker, T.; Šimková, H.; Fossati, D.; Moullet, O.; Brabant, C.; Vrána, J.; Doležel, J.; Krattinger, S.G. Rapid cloning of genes in hexaploid wheat using cultivar-specific long-range chromosome assembly. Nat. Biotechnol. 2017, 35, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dracatos, P.M.; Barto¡, J.; Elmansour, H.; Singh, D.; Karafiátová, M.; Zhang, P.; Steuernagel, B.; Svačina, R.; Cobbin, J.C.A.; Clark, B.; et al. The Coiled-Coil NLR Rph1, Confers Leaf Rust Resistance in Barley Cultivar Sudan. Plant Physiol. 2018, 179, 1362–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bariana, H.S.; Babu, P.; Forrest, K.L.; Park, R.F.; Bansal, U.K. Discovery of the New Leaf Rust Resistance Gene Lr82 in Wheat: Molecular Mapping and Marker Development. Genes 2022, 13, 964. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Baldauf, S.; Lim, E.-K.; Bowles, D.J. Phylogenetic Analysis of the UDP-glycosyltransferase Multigene Family of Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 4338–4343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.T.; Koo, Y.J.; Seo, H.S.; Kim, M.C.; Do Choi, Y.; Kim, J.H. Overexpression of AtSGT1, an Arabidopsis salicylic acid glucosyltransferase, leads to increased susceptibility to Pseudomonas syringae. Phytochemistry 2008, 69, 1128–1134. [Google Scholar] [CrossRef]
- George Thompson, A.M.; Iancu, C.V.; Neet, K.E.; Dean, J.V.; Choe, J.-y. Differences in salicylic acid glucose conjugations by UGT74F1 and UGT74F2 from Arabidopsis thaliana. Sci. Rep. 2017, 7, 46629. [Google Scholar] [CrossRef] [Green Version]
- Parlevliet, J.E.; Van Ommeren, A. Partial resistance of barley to leaf rust, Puccinia hordei. II. Relationship between field trials, micro plot tests and latent period. Euphytica 1975, 24, 293–303. [Google Scholar] [CrossRef]
- Parlevliet, J. Further evidence of polygenic inheritance of partial resistance in barley to leaf rust, Puccinia hordei. Euphytica 1978, 27, 369–379. [Google Scholar] [CrossRef]
- He, Y.; Xu, J.; Wang, X.; He, X.; Wang, Y.; Zhou, J.; Zhang, S.; Meng, X. The Arabidopsis pleiotropic drug resistance transporters PEN3 and PDR12 mediate camalexin secretion for resistance to Botrytis cinerea. Plant Cell 2019, 31, 2206–2222. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Lagudah, E.S.; Spielmeyer, W.; Singh, R.P.; Huerta-Espino, J.; McFadden, H.; Bossolini, E.; Selter, L.L.; Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Rinaldo, A.; Gilbert, B.; Boni, R.; Krattinger, S.G.; Singh, D.; Park, R.F.; Lagudah, E.; Ayliffe, M. The Lr34 adult plant rust resistance gene provides seedling resistance in durum wheat without senescence. Plant Biotechnol. J. 2017, 15, 894–905. [Google Scholar] [CrossRef] [Green Version]
- Boni, R.; Chauhan, H.; Hensel, G.; Roulin, A.; Sucher, J.; Kumlehn, J.; Brunner, S.; Krattinger, S.G.; Keller, B. Pathogen-inducible Ta-Lr34res expression in heterologous barley confers disease resistance without negative pleiotropic effects. Plant Biotechnol. J. 2018, 16, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Krattinger, S.G.; Sucher, J.; Selter, L.L.; Chauhan, H.; Zhou, B.; Tang, M.; Upadhyaya, N.M.; Mieulet, D.; Guiderdoni, E.; Weidenbach, D. The wheat durable, multipathogen resistance gene Lr34 confers partial blast resistance in rice. Plant Biotechnol. J. 2016, 14, 1261–1268. [Google Scholar] [CrossRef]
- Denancé, N.; Szurek, B.; Noël, L.D. Emerging functions of nodulin-like proteins in non-nodulating plant species. Plant Cell Physiol. 2014, 55, 469–474. [Google Scholar] [CrossRef]
- Gyawali, S.; Chao, S.; Vaish, S.S.; Singh, S.P.; Rehman, S.; Vishwakarma, S.R.; Verma, R.P.S. Genome wide association studies (GWAS) of spot blotch resistance at the seedling and the adult plant stages in a collection of spring barley. Mol. Breed. 2018, 38, 62. [Google Scholar] [CrossRef]
- Milne, R.J.; Dibley, K.E.; Schnippenkoetter, W.; Mascher, M.; Lui, A.C.; Wang, L.; Lo, C.; Ashton, A.R.; Ryan, P.R.; Lagudah, E.S. The wheat Lr67 gene from the sugar transport protein 13 family confers multipathogen resistance in barley. Plant Physiol. 2019, 179, 1285–1297. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef]
- Dinh, H.X.; Singh, D.; de la Cruz, D.G.; Hensel, G.; Kumlehn, J.; Mascher, M.; Stein, N.; Perovic, D.; Ayliffe, M.; Moscou, M.J. The barley leaf rust resistance gene Rph3 encodes a predicted membrane protein and is induced upon infection by avirulent pathotypes of Puccinia hordei. Nat. Commun. 2022, 13, 2386. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | a Isolate | b Chr | c cM | p-Value | FDR | R2(%) | Allele Frequency | d Allele Effect | GAPIT and TASSEL Models |
|---|---|---|---|---|---|---|---|---|---|
| SCRI_RS_118785 | SRT-MRC | 1 | 56.44 | 1.30 × 10−03 | 1.36 × 10−03 | 3.34 | 53 | A (−0.400) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-51304 | SRT-MRC | 1 | 116.78 | 3.30 × 10−04 | 8.58 × 10−04 | 4.19 | 234 | C (−0.382) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-51989 | SRT-MRC | 1 | 118.13 | 1.31 × 10−03 | 1.36 × 10−03 | 3.33 | 211 | C (−0.309) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| ** JHI-Hv50k-2016-117483 | SRT-MRC | 2 | 106.53 | 5.50 × 10−04 | 1.14 × 10−03 | 3.86 | 286 | A (−0.484) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-191945 | SRT-MRC | 3 | 66.69 | 7.90 × 10−04 | 1.14 × 10−03 | 3.67 | 127 | A (−0.309) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-195152 | SRT-MRC | 3 | 75.21 | 5.30 × 10−04 | 1.14 × 10−03 | 3.91 | 135 | C (0.355) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-353355 | SRT-MRC | 5 | 151.88 | 1..21 × 10−03 | 1..33 × 10−03 | 3.52 | 81 | C (−0.374) | MLM (K + Q), MLM (K + PCA), BLINK, FarmCPU |
| JHI-Hv50k-2016-409979 | SRT-MRC | 6 | 66.08 | 4.20 × 10−04 | 1.05 × 10−03 | 4.03 | 263 | A (−0.477) | MLM (K + Q), MLM (K + PCA) |
| ** JHI-Hv50k-2016-511500 | SRT-MRC | 7 | 126.7 | 1.32 × 10−03 | 1.36 × 10−03 | 3.35 | 181 | C (0.312) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-515270 | SRT-MRC | 7 | 132.22 | 2.00 × 10−04 | 7.74 × 10−04 | 4.66 | 229 | C (0.371) | MLM (K + Q, MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-5369 | SRT-SAT | 1 | 4.96 | 2.50 × 10−04 | 7.74 × 10−04 | 4.39 | 231 | A (0.409) | MLM (K + Q), MLM (K + PCA), FarmCPU |
| JHI-Hv50k-2016-41795 | SRT-SAT | 1 | 89.31 | 7.10 × 10−04 | 1.14 × 10−03 | 3.7 | 297 | A (0.691) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-107670 | SRT-SAT | 2 | 80.03 | 7.30 × 10−04 | 1.14 × 10−03 | 3.71 | 79 | C (0.407) | MLM (K + Q), MLM (K + PCA), BLINK |
| JHI-Hv50k-2016-109913 | SRT-SAT | 2 | 88.39 | 4.50 × 10−04 | 1.08 × 10−03 | 3.99 | 223 | A (−0.386) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK |
| JHI-Hv50k-2016-111647 | SRT-SAT | 2 | 89.77 | 2.23 × 10-10 | 3.74 × 10−09 | 13.65 | 30 | A (−1.14) | MLM (K + Q), MLM (K + PCA), BLINK, FarmCPU |
| JHI-Hv50k-2016-110777 | SRT-SAT | 2 | 90.16 | 2.23 × 10-10 | 3.74 × 10−09 | 13.65 | 286 | C (1.14) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-111042 | SRT-SAT | 2 | 90.72 | 9.51 × 10−07 | 8.83 × 10−06 | 7.94 | 55 | A (−0.693) | MLM (K +Q), MLM (K + PCA), BLINK, FarmCPU |
| JHI-Hv50k-2016-111819 | SRT-SAT | 2 | 91.01 | 2.23 × 10-10 | 3.74 × 10−09 | 13.65 | 30 | C (−1.14) | MLM (K +Q), MLM (K + PCA) |
| JHI-Hv50k-2016-112041 | SRT-SAT | 2 | 91.15 | 6.97 × 10-10 | 9.06 × 10−09 | 12.85 | 29 | C (−1.13) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-113615 | SRT-SAT | 2 | 92.71 | 7.70 × 10−04 | 1.14 × 10−03 | 3,66 | 276 | C (0.53029) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-111864 | SRT-SAT | 2 | 92.78 | 2.30 × 10-10 | 3.74 × 10−09 | 13.67 | 283 | C (1.14) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-113653 | SRT-SAT | 2 | 95.16 | 3.00 × 10−05 | 2.44 × 10−04 | 5.69 | 261 | C (0.584) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-113742 | SRT-SAT | 2 | 97.8 | 6.00 × 10−05 | 3.90 × 10−04 | 5.24 | 292 | C (0.802) | MLM (K + Q), MLM (K + PCA), BLINK |
| ** JHI-Hv50k-2016-119586 | SRT-SAT | 2 | 107.37 | 1.21 × 10−03 | 1.33 × 10−03 | 3.38 | 154 | A (−0.336) | MLM (K + Q), MLM (K + PCA) |
| SCRI_RS_14819 | SRT-SAT | 3 | 2.69 | 9.10 × 10−04 | 1.15 × 10−03 | 3.6 | 21 | A (−0.634) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK |
| JHI-Hv50k-2016-189928 | SRT-SAT | 3 | 63.39 | 6.20 × 10−04 | 1.14 × 10−03 | 3.79 | 74 | C (−0.469) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-338772 | SRT-SAT | 5 | 130.69 | 1.25 × 10−03 | 1.35 × 10−03 | 3.44 | 282 | C (−0.534) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-310432 | SRT-SAT | 5 | 66.69 | 1.10 × 10−04 | 5.50 × 10−04 | 4.85 | 156 | A (−0.373) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-318662 | SRT-SAT | 5 | 90.03 | 7.90 × 10−04 | 1.14 × 10−03 | 3.64 | 95 | A (−0.422) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-430648 | SRT-SAT | 6 | 118.98 | 2.70 × 10−04 | 7.98 × 10−04 | 4.3 | 292 | A (−0.677) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-431728 | SRT-SAT | 6 | 121.68 | 6.50 × 10−04 | 1.14 × 10−03 | 3.77 | 32 | A (0.541) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK |
| JHI-Hv50k-2016-435962 | SRT-SAT | 7 | 0.21 | 1.39 × 10−03 | 1.39 × 10−03 | 3.3 | 235 | A (−0.354) | MLM (K + Q), MLM (K + PCA), MLMM, FarmCPU |
| JHI-Hv50k-2016-465087 | SRT-SAT | 7 | 48.73 | 2.50 × 10−04 | 7.74 × 10−04 | 4.36 | 299 | G (−0.767) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK |
| JHI-Hv50k-2016-466598 | SRT-SAT | 7 | 50.71 | 3.20 × 10−04 | 8.58 × 10−04 | 4.19 | 235 | C (0.391) | MLM (K + Q), MLM (K + PCA) |
| Marker | a Env | b Chr | c cM | p-Value | FDR | R2(%) | Allele Frequency | d Allele Effect | Best-Fit TASSEL and GAPIT Models |
|---|---|---|---|---|---|---|---|---|---|
| BOPA2_12_30298 | APR-SAT17 | 1 | 66.29 | 9.23 × 10−04 | 1.15 × 10−03 | 3.7 | 78 | A (−63.43) | GLM (Q), MLM (K + Q), MLM (K + /PCA) |
| JHI-Hv50k-2016-80368 | APR-SAT17 | 2 | 43.84 | 9.57 × 10−04 | 1.17 × 10−03 | 3.65 | 19 | A (−113.86) | GLM (Q), MLM (K + Q). MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-80601 | APR-SAT17 | 2 | 50.92 | 1.01 × 10−03 | 1.22 × 10−03 | 3.62 | 62 | C (−68.16) | GLM (Q), MLM (K + Q), MLM (K + PCA), FarmCPU |
| SCRI_RS_126890 | APR-SAT17 | 2 | 105.56 | 1.22 × 10−04 | 5.66 × 10−04 | 5.03 | 204 | A (−69.22) | GLM (Q), MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| ** SCRI_RS_153420 | APR-SAT17 | 2 | 107.26 | 6.90 × 10−04 | 1.14 × 10−03 | 3.89 | 194 | C (−59.36) | GLM (Q), MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| SCRI_RS_135365 | APR-SAT17 | 4 | 59.99 | 8.39 × 10−04 | 1.14 × 10−03 | 3.85 | 263 | C (−84.34) | GLM (Q), MLM (K + Q), MLM (K + PCA), BLINK, FarmCPU |
| JHI-Hv50k-2016-277557 | APR-SAT17 | 5 | 1.11 | 9.20 × 10−05 | 4.98 × 10−04 | 5.25 | 162 | A (−66.16) | GLM (Q), MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-277561 | APR-SAT17 | 5 | 1.45 | 4.96 × 10−04 | 1.14 × 10−03 | 4.22 | 143 | C (59.30) | GLM (Q), MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-286440 | APR-SAT17 | 5 | 38.12 | 2.46 × 10−04 | 7.74 × 10−04 | 4.71 | 78 | A (−70.43) | GLM (Q), MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-415383 | APR-SAT17 | 6 | 75.78 | 8.41 × 10−04 | 1.14 × 10−03 | 3.76 | 43 | A (−79.99) | GLM (Q), MLM (K + Q), MLM (K + PCA), BLINK |
| JHI-Hv50k-2016-65232 | APR-SAT19 | 2 | 7.44 | 1.14 × 10−03 | 1.32 × 10−03 | 3.72 | 273 | C (12.87) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-201579 | APR-SAT19 | 2 | 57.01 | 7.72 × 10−04 | 1.14 × 10−03 | 3.98 | 104 | A (7.31) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-201394 | APR-SAT19 | 3 | 91.22 | 8.84 × 10−04 | 1.15 × 10−03 | 3.89 | 169 | C (−7.33) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-201622 | APR-SAT19 | 3 | 93.81 | 7.17 × 10−04 | 1.14 × 10−03 | 4.03 | 194 | A (−7.64) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-202076 | APR-SAT19 | 3 | 97.8 | 1.19 × 10−03 | 1.33 × 10−03 | 3.69 | 107 | A (−7.21) | MLM (K + Q), MLM (K + PCA) |
| BOPA2_12_31536 | APR-SAT19 | 4 | 63.39 | 5.88 × 10−05 | 3.90 × 10−04 | 5.73 | 142 | C (−8.31) | MLM (K + Q), MLM (K + PCA), MLMM,, BLINK, FarmCPU |
| JHI-Hv50k-2016-320334 | APR-SAT19 | 5 | 94.72 | 3.20 × 10−04 | 8.58 × 10−04 | 4.6 | 188 | A (−7.27) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| JHI-Hv50k-2016-342925 | APR-SAT19 | 5 | 136.6 | 8.23 × 10−04 | 1.14 × 10−03 | 3.94 | 21 | C (−12.89) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-364245 | APR-SAT19 | 5 | 168.12 | 8.04 × 10−04 | 1.14 × 10−03 | 4.12 | 15 | C (13.85) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-366258 | APR-SAT19 | 5 | 169.38 | 6.92 × 10−04 | 1.14 × 10−03 | 4.05 | 17 | C (−14.27) | MLM (K + Q), MLM (K + PCA) |
| JHI-Hv50k-2016-201456 | APR-SAT19 | 7 | 76.84 | 8.84 × 10−04 | 1.15 × 10−03 | 3.89 | 122 | C (7.33) | MLM (K + Q), MLM (K + PCA), MLMM |
| JHI-Hv50k-2016-496331 | APR-SAT19 | 7 | 97.31 | 1.36 × 10−03 | 1.38 × 10−03 | 3.63 | 262 | C (−10.70) | MLM, (K + Q), MLM (K + PCA) MLMM, BLINK, FarmCPU |
| SCRI_RS_150401 | APR-SAT19 | 7 | 110.27 | 7.06 × 10−05 | 4.17 × 10−04 | 5.64 | 38 | C (11.35) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
| ** JHI-Hv50k-2016-510780 | APR-SAT19 | 7 | 126.7 | 1.06 × 10−03 | 1.25 × 10−03 | 3.77 | 261 | C (−11.12) | MLM (K + Q), MLM (K + PCA), MLMM, BLINK, FarmCPU |
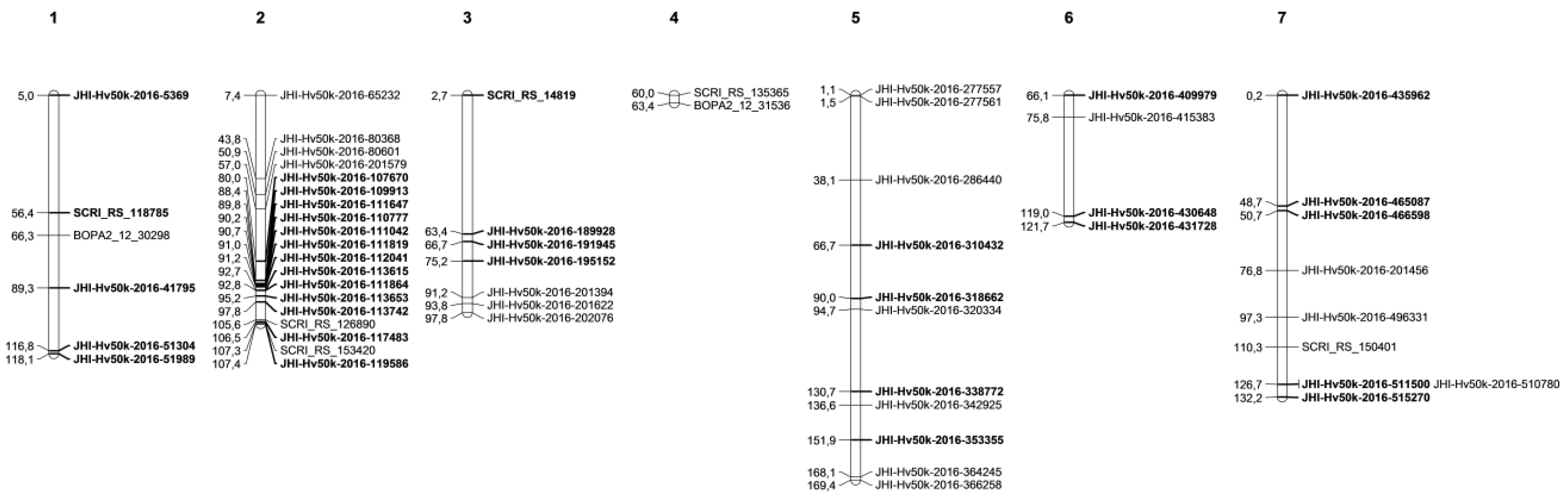
| Associated Marker | a Chr | b cM | Gene Identifier | Homology | c Previously Mapped LR QTL/Genes |
|---|---|---|---|---|---|
| Seedling resistance (SRT) | |||||
| SRT-MRC | |||||
| SCRI_RS_118785 | 1 | 56.44 | HORVU1Hr1G059300 | Receptor-like protein kinase | _ |
| JHI-Hv50k-2016-51304 | 1 | 116.78 | HORVU1Hr1G086590 | ROP guanine nucleotide exchange factor 5 | RphQ3 [35] |
| JHI-Hv50k-2016-51989 | 1 | 118.13 | HORVU1Hr1G087340 | Guanylate-binding family protein | RphQ18 (RphQ3) [35,66] |
| ** JHI-Hv50k-2016-117483 | 2 | 106.53 | HORVU2Hr1G103180 | Terpene synthase 04 | _ |
| JHI-Hv50k-2016-191945 | 3 | 66.69 | HORVU3Hr1G074330 | Nucleic acid binding | Rph10.o [67] |
| JHI-Hv50k-2016-195152 | 3 | 75.21 | HORVU3Hr1G079230 | Leucine-rich repeat protein kinase family protein | _ |
| JHI-Hv50k-2016-353355 | 5 | 151.88 | HORVU5Hr1G114220 | Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family | Rph9.z [67] |
| JHI-Hv50k-2016-409979 | 6 | 66.08 | HORVU6Hr1G070350 | Undescribed protein | _ |
| ** JHI-Hv50k-2016-511500 | 7 | 126.7 | HORVU7Hr1G115100 | RphQ14 (Rph3/19); Rph3.c; RphQ28 [35,66,67] | |
| JHI-Hv50k-2016-515270 | 7 | 132.22 | HORVU7Hr1G118430 | Disease resistance RPP13-like protein 4 | _ |
| SRT-SAT | |||||
| JHI-Hv50k-2016-5369 | 1 | 4.96 | HORVU1Hr1G002310 | 50S ribosomal protein L2 | Rphq14 [68] |
| JHI-Hv50k-2016-41795 | 1 | 89.31 | HORVU1Hr1G075680 | Receptor-like protein kinase 5 | _ |
| JHI-Hv50k-2016-107670 | 2 | 80.03 | HORVU2Hr1G092270 | Lactoylglutathione lyase/glyoxalase I family protein | _ |
| JHI-Hv50k-2016-109913 | 2 | 88.39 | HORVU2Hr1G096190 | Disease resistance protein RPP13 | _ |
| JHI-Hv50k-2016-111647 | 2 | 89.77 | HORVU2Hr1G097860 | Integrase-type DNA-binding superfamily protein | Rphq11 [68] |
| JHI-Hv50k-2016-110777 | 2 | 90.16 | HORVU2Hr1G096910 | UDP-Glycosyltransferase superfamily protein | Rphq11 [68] |
| JHI-Hv50k-2016-111042 | 2 | 90.72 | HORVU2Hr1G097140 | Zinc finger CCHC domain-containing protein 9 | _ |
| JHI-Hv50k-2016-111819 | 2 | 91.01 | HORVU2Hr1G097940 | Homeobox-leucine zipper protein 4 | _ |
| JHI-Hv50k-2016-112041 | 2 | 91.15 | HORVU2Hr1G098100 | Disease resistance protein | _ |
| JHI-Hv50k-2016-113615 | 2 | 92.71 | HORVU2Hr1G099350 | Alcohol dehydrogenase 1 | _ |
| JHI-Hv50k-2016-111864 | 2 | 92.78 | HORVU2Hr1G097980 | Serine/arginine repetitive matrix protein 1 isoform X1 | _ |
| JHI-Hv50k-2016-113653 | 2 | 95.16 | HORVU2Hr1G099440 | Terpene synthase 04 | _ |
| JHI-Hv50k-2016-113742 | 2 | 97.8 | HORVU2Hr1G099470 | Bifunctional inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein | _ |
| ** JHI-Hv50k-2016-119586 | 2 | 107.37 | HORVU2Hr1G105090 | Protein of unknown function (DUF581) | _ |
| SCRI_RS_14819 | 3 | 2.69 | HORVU3Hr1G002010 | Pentatricopeptide repeat-containing protein | QRph-3H.6; Rph5; Rph7.g [67,69,70] |
| JHI-Hv50k-2016-189928 | 3 | 63.39 | _ | _ | Rph10.o [67] |
| JHI-Hv50k-2016-310432 | 5 | 66.69 | ADQ48070.1 | bZIP transcription factor 27 | _ |
| JHI-Hv50k-2016-318662 | 5 | 90.03 | HORVU5Hr1G078310 | Protein FLOWERING LOCUST | 11_11473 [22] |
| JHI-Hv50k-2016-338772 | 5 | 130.69 | EKC/KEOPS complex subunit bud32 | RphQ25; QLr.S42-5H.a (Rph9; Rph12) [66,71,72] | |
| JHI-Hv50k-2016-430648 | 6 | 118.98 | HORVU6Hr1G092660 | Disease resistance RPP8-like protein 3 | QPh.6H-3 (QTL_Backes) [73,74] |
| JHI-Hv50k-2016-431728 | 6 | 121.68 | HORVU6Hr1G093310 | dTDP-4-dehydrorhamnose reductase | _ |
| JHI-Hv50k-2016-435962 | 7 | 0.21 | HORVU7Hr1G000040 | Disease resistance protein (CC-NBS-LRR class) family | RphQ12 [35] |
| JHI-Hv50k-2016-465087 | 7 | 48.73 | HORVU7Hr1G030310 | Formin Homology 14 | _ |
| JHI-Hv50k-2016-466598 | 7 | 50.71 | HORVU7Hr1G031610 | Protein tesmin/TSO1-like CXC 5 | _ |
| Adult Plant Resistance (APR) | |||||
| APR-SAT17 | |||||
| BOPA2_12_30298 | 1 | 66.29 | HORVU1Hr1G066050 | Cyclic nucleotide-gated channel 14 | _ |
| JHI-Hv50k-2016-80368 | 2 | 43.84 | HORVU2Hr1G022560 | ABC transporter C family member 10 | Rph16. RphQ20 [66,75] |
| JHI-Hv50k-2016-80601 | 2 | 50.92 | HORVU2Hr1G022900 | Ankyrin repeat family protein | RphQ7 [35] |
| SCRI_RS_126890 | 2 | 105.56 | HORVU2Hr1G102880 | Protein of unknown function (DUF760) | _ |
| ** SCRI_RS_153420 | 2 | 107.26 | HORVU2Hr1G102930 | High mobility group B protein 6 | _ |
| SCRI_RS_135365 | 4 | 59.99 | HORVU4Hr1G064860 | LisH and RanBPM domains containing protein | MQTL9 (Rphq19) [26,76,77] |
| JHI-Hv50k-2016-277557 | 5 | 1.11 | AAV49984.1 | Nodulin-like/Major Facilitator Superfamily protein | _ |
| JHI-Hv50k-2016-277561 | 5 | 1.45 | AAV49985.1 | Hordoindoline-B1 | _ |
| JHI-Hv50k-2016-286440 | 5 | 38.12 | HORVU5Hr1G010930 | Unknown function/putative serine protease do-like htrA | Rph2.t; Rph2 (Rph2.b) [67,78] |
| JHI-Hv50k-2016-415383 | 6 | 75.78 | HORVU6Hr1G077750 | Phosphate-responsive 1 family protein | _ |
| APR-SAT19 | |||||
| JHI-Hv50k-2016-65232 | 2 | 7.44 | HORVU2Hr1G005650 | Cullin-associated NEDD8-dissociated protein 1 | Rph1.a; RphQ19 (RphQ5/6) [66,67] |
| JHI-Hv50k-2016-201579 | 2 | 57.01 | _ | _ | _ |
| JHI-Hv50k-2016-201394 | 3 | 91.22 | HORVU3Hr1G086030 | Eukaryotic aspartyl protease family protein | _ |
| JHI-Hv50k-2016-201622 | 3 | 93.81 | HORVU3Hr1G086290 | Pentatricopeptide repeat-containing protein | _ |
| JHI-Hv50k-2016-202076 | 3 | 97.8 | ACB56486.1 | Respiratory burst oxidase protein F | MQTL7C (Rphq20) [26,77] |
| BOPA2_12_31536 | 4 | 63.39 | HORVU4Hr1G068990 | Nodulin-related protein 1, putative | MQTL9; Rphq19 [26,76,77] |
| JHI-Hv50k-2016-320334 | 5 | 94.72 | HORVU5Hr1G080280 | ATPase ASNA1 homolog | _ |
| JHI-Hv50k-2016-342925 | 5 | 136.6 | HORVU5Hr1G103380 | Mitochondrial transcription termination factor family protein | Rph9.i [67] |
| JHI-Hv50k-2016-364245 | 5 | 168.12 | HORVU5Hr1G123050 | Chromatin accessibility complex protein 1 | QLr.HEB-5-5H.b5H MQTL13 (Rphq7), MQTL14 (Rphq16) [26,76,77] |
| JHI-Hv50k-2016-366258 | 5 | 169.38 | HORVU5Hr1G124630 | Glutathione S-transferase T1 | _ |
| JHI-Hv50k-2016-201456 | 7 | 76.84 | HORVU3Hr1G086080 | S-acyltransferase | _ |
| JHI-Hv50k-2016-496331 | 7 | 97.31 | HORVU7Hr1G100300 | Disease resistance protein | RphQ13, Rphq9 [23,35] |
| SCRI_RS_150401 | 7 | 110.27 | HORVU7Hr1G106860 | Glycine-rich domain-containing protein 2 | QPh.7H-3 (Rphq9), QTL_Castro, “Ris44-Bmac156” [26,73,79] |
| ** JHI-Hv50k-2016-510780 | 7 | 126.7 | HORVU7Hr1G114330 | Homogentisate phytyltransferase 1 | RphQ14 (Rph3/19) Rph3.c; RphQ28 [35,66,67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amouzoune, M.; Rehman, S.; Benkirane, R.; Verma, S.; Gyawali, S.; Al-Jaboobi, M.; Verma, R.P.S.; Kehel, Z.; Amri, A. Genome-Wide Association Study of Leaf Rust Resistance at Seedling and Adult Plant Stages in a Global Barley Panel. Agriculture 2022, 12, 1829. https://doi.org/10.3390/agriculture12111829
Amouzoune M, Rehman S, Benkirane R, Verma S, Gyawali S, Al-Jaboobi M, Verma RPS, Kehel Z, Amri A. Genome-Wide Association Study of Leaf Rust Resistance at Seedling and Adult Plant Stages in a Global Barley Panel. Agriculture. 2022; 12(11):1829. https://doi.org/10.3390/agriculture12111829
Chicago/Turabian StyleAmouzoune, Mariam, Sajid Rehman, Rachid Benkirane, Swati Verma, Sanjaya Gyawali, Muamar Al-Jaboobi, Ramesh Pal Singh Verma, Zakaria Kehel, and Ahmed Amri. 2022. "Genome-Wide Association Study of Leaf Rust Resistance at Seedling and Adult Plant Stages in a Global Barley Panel" Agriculture 12, no. 11: 1829. https://doi.org/10.3390/agriculture12111829