
Genetic Diversity of Black Amaranth (Amaranthus quitensis Kunth) Landraces of Ecuadorian Highlands: Association Genotypes—Color Morphotypes

Abstract
:1. Introduction
2. Materials and Methods
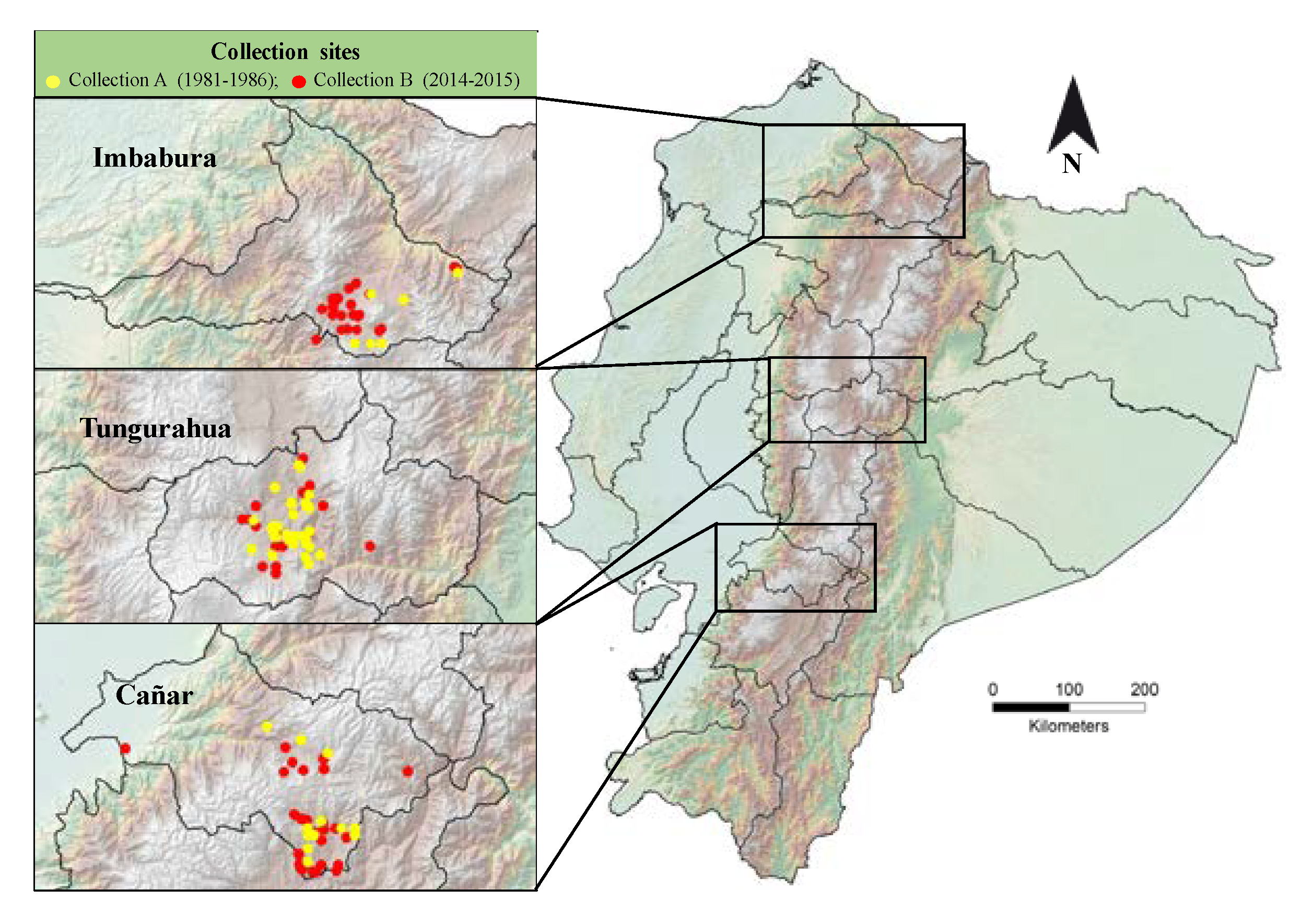
2.1. Plant Material
2.2. DNA Extraction and Microsatellite Genotyping
2.3. Data Analysis
3. Results
3.1. Genetic Diversity
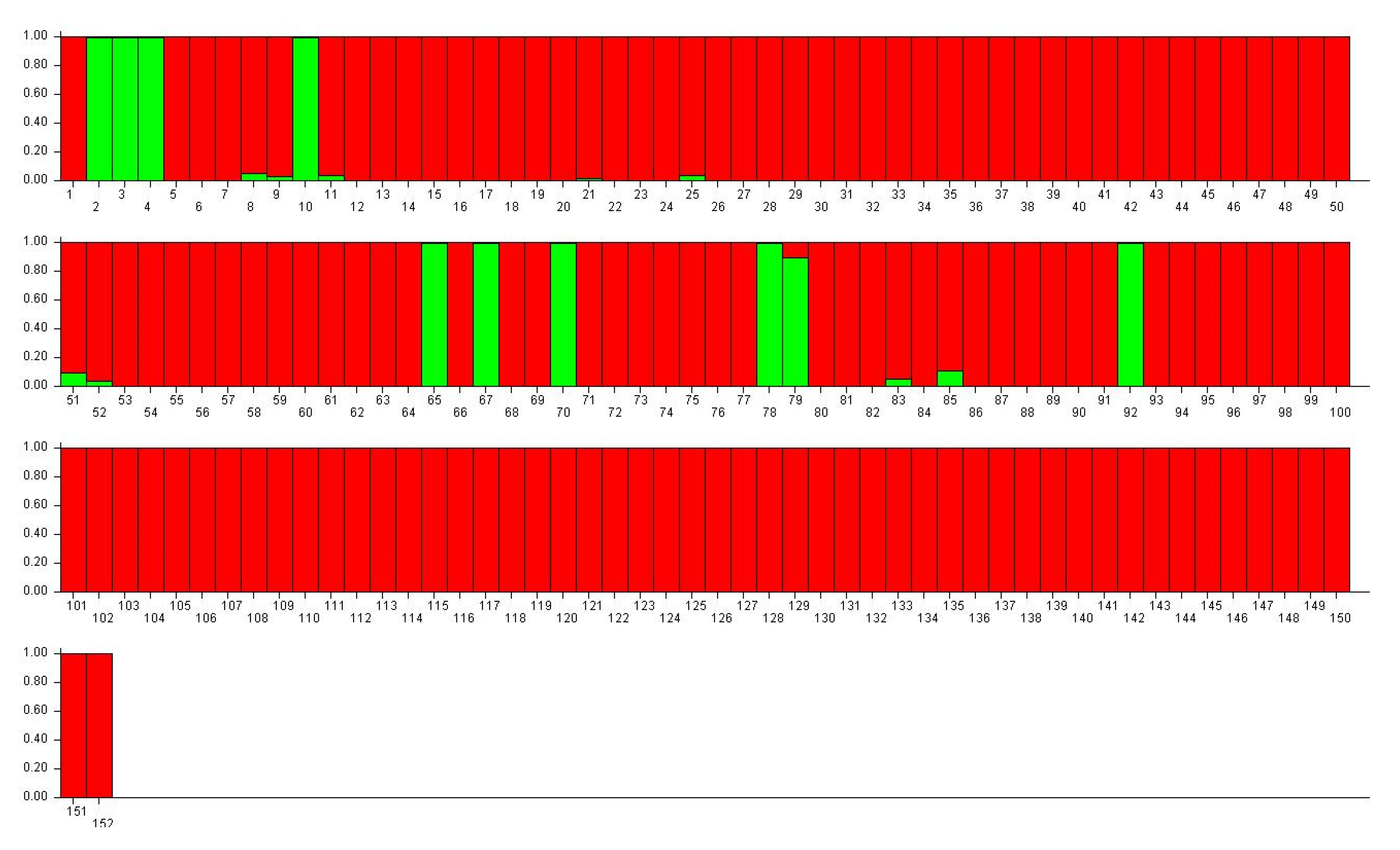
3.2. Genetic Structure
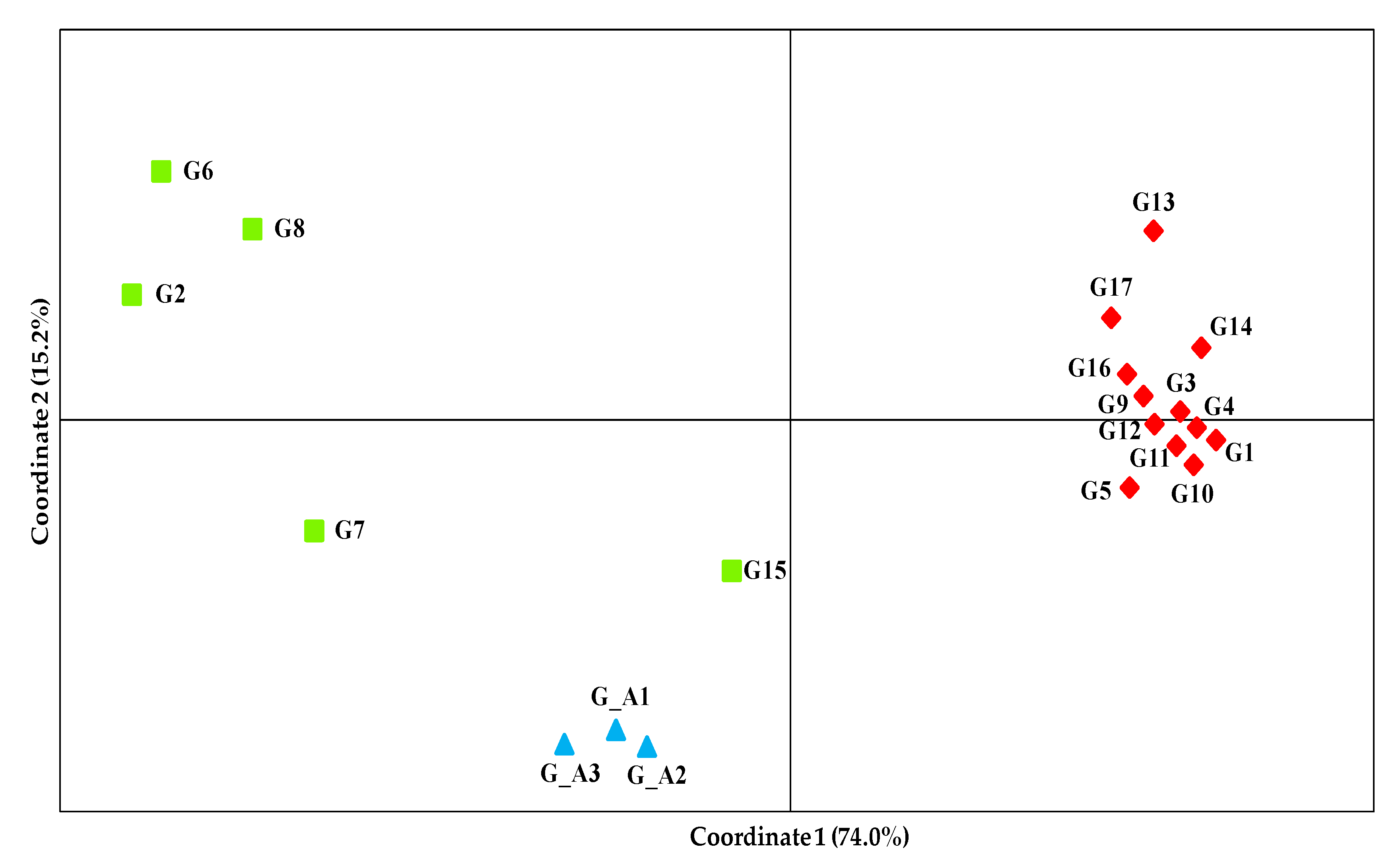
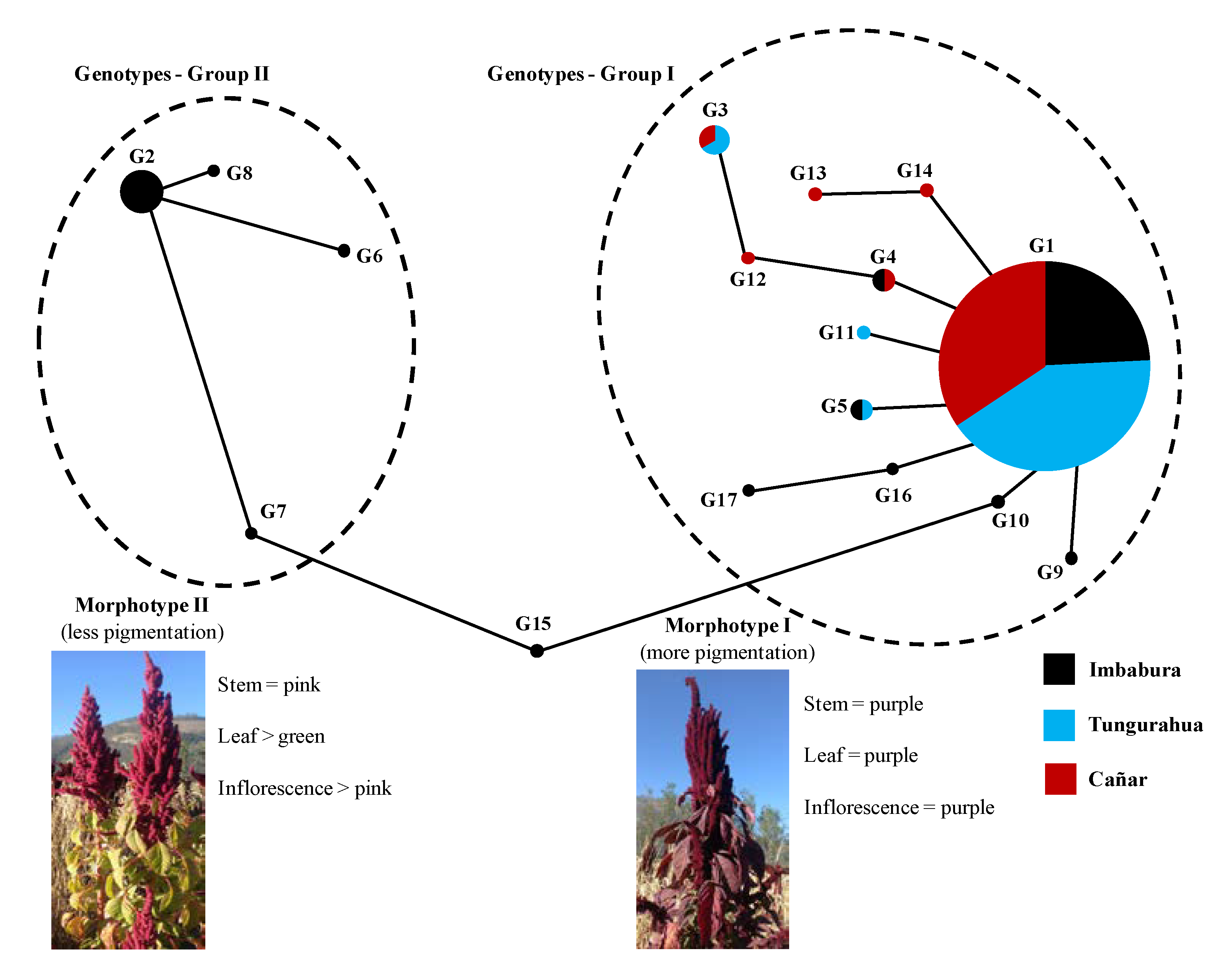
3.3. Genetic Relationships among Genotypes
4. Discussion
4.1. Genetic Diversity
4.2. Genetic Structure and Association between Genotypes and Color Morphotypes
4.3. Implications for Conservation of Black Amaranth Landraces
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sauer, J.D. The grain amaranths and their relatives: A revised taxonomic and geographic survey. Ann. Mo. Bot. Gard. 1967, 54, 103–137. [Google Scholar] [CrossRef]
- Stetter, M.G.; Schmid, K.J. Analysis of phylogenetic relationships and genome size evolution of the Amaranthus genus using GBS indicates the ancestors of an ancient crop. Mol. Phylogenet. Evol. 2017, 109, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Waselkov, K.E.; Boleda, A.S.; Olsen, K.M. A phylogeny of the genus Amaranthus (Amaranthaceae) based on several low-copy nuclear loci and chloroplast regions. Syst. Bot. 2018, 43, 439–458. [Google Scholar] [CrossRef]
- Thapa, R.; Blair, M.W. Morphological Assessment of Cultivated and Wild Amaranth Species Diversity. Agronomy 2018, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Park, Y.J. Evaluation of genetic structure of amaranth accessions from the United States. Weed Turfgrass Sci. 2013, 2, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Oo, W.H.; Park, J.Y. Analysis of the Genetic Diversity and Population Structure of Amaranth Accessions from South America Using 14 SSR Markers. Korean J. Crop. Sci. 2013, 58, 336–346. [Google Scholar] [CrossRef]
- Sauer, J.D. Grain amaranths. Evolution of Crop Plants; Simmonds, N.W., Ed.; Longman Group: London, UK, 1976; pp. 4–7. [Google Scholar]
- Stetter, M.G.; Vidal-Villarejo, M.; Schmid, K.J. Parallel seed color adaptation during multiple domestication attempts of an ancient New World grain. Mol. Biol. Evol. 2020, 37, 1407–1419. [Google Scholar] [CrossRef]
- Clouse, J.W.; Adhikary, D.; Page, J.T.; Ramaraj, T.; Deyholos, M.K.; Udall, J.A.; Fairbanks, D.J.; Jellen, E.N.; Maughan, P.J. The amaranth genome: Genome, transcriptome, and physical map assembly. Plant Genome 2016, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Kietlinski, K.; Jimenez, F.; Jellen, E.N.; Maughan, P.J.; Smith, S.M.; Pratt, D.B. Relationships between the Weedy (Amaranthaceae) and the Grain Amaranths. Crop. Sci. 2014, 54, 220–228. [Google Scholar] [CrossRef]
- Thapa, R.; Edwards, M.; Blair, M.W. Relationship of Cultivated Grain Amaranth Species and Wild Relative Accessions. Genes 2021, 12, 1849. [Google Scholar] [CrossRef]
- Peralta, E.; Villacrés, E.; Mazón, N.; Rivera, M.; Subía, C. El Ataco, Sangorache o Amaranto Negro (Amaranthus hybridus L.) en Ecuador; Publicación Miscelánea No.143. Programa Nacional de Leguminosas y Granos Andinos; Estación Experimental Santa Catalina INIAP: Quito, Ecuador, 2008; p. 63. [Google Scholar]
- Aderibigbe, O.R.; Ezekiel, O.O.; Owolade, S.O.; Korese, J.K.; Sturm, B.; Hensel, O. Exploring the potentials of underutilized grain amaranth (Amaranthus spp.) along the value chain for food and nutrition security: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, A.; Shukla, S. Amaranth: A new millennium crop of nutraceutical values. Critical Reviews. Food Sci. Nutr. 2013, 53, 109–125. [Google Scholar] [CrossRef]
- Martínez-López, A.; Milla-Linares, M.C.; Rodríguez-Martín, N.M.; Millán, F.; Montserrat-de la Paz, M. Nutraceutical value of kiwicha (Amaranthus caudatus L.). J. Funct. Foods 2020, 65, 103735. [Google Scholar] [CrossRef]
- Ballabio, C.; Uberti, F.; Di Lorenzo, C.; Brandolini, A.; Penas, E.; Restani, P. Biochemical and immunochemical characterization of different varieties of amaranth (Amaranthus L. ssp.) as a safe ingredient for gluten-free products. J. Agric. Food Chem. 2011, 59, 12969–12974. [Google Scholar] [CrossRef]
- Massange-Sánchez, J.A.; Palmeros-Suárez, P.A.; Martínez-Gallardo, N.A.; Castrillón-Arbelaez, P.A.; Avilés-Arnaut, H.; Alatorre-Cobos, F.; Tiessen, A.; Délano-Frier, J.P. The novel and taxonomically restricted Ah24 gene from grain amaranth (Amaranthus hypochondriacus) has a dual role in development and defense. Front. Plants Sci. 2015, 6, 602. [Google Scholar] [CrossRef] [Green Version]
- Saucedo, A.L.; Hernández-Domínguez, E.E.; de Luna-Valdez, L.A.; Guevara-García, A.A.; Escobedo-Moratilla, A.; Bojorquéz-Velázquez, E.; Del Río-Portilla, F.; Fernández-Velasco, D.A.; Barba de la Rosa, A.P. Insights on structure and function of a late embryogenesis abundant protein from Amaranthus cruentus: An intrinsically disordered protein involved in protection against desiccation, oxidant conditions, and osmotic stress. Front. Plant Sci. 2017, 8, 497. [Google Scholar] [CrossRef]
- Peralta, E. El amaranto en Ecuador: “Estado del Arte”; Programa Nacional de Leguminosas y Granos Andinos Estación; Experimental Santa Catalina INIAP: Quito, Ecuador, 2012; p. 42. [Google Scholar]
- Cai, Y.; Sun, M.; Wu, H.; Huang, R.; Corke, H. Characterization and Quantification of Betacynanin Pigments from Diverse Amaranthus Species. J. Agric. Food Chem. 1998, 46, 2064–2070. [Google Scholar] [CrossRef]
- Peralta, E. Producción y distribución de semilla de buena calidad con pequeños agricultores de GRANOS ANDINOS: Chocho, quinua, Amaranto-sistema no convencional-Publicación Miscelánea No. 169. Programa Nacional de Leguminosas y Granos Andinos; Estación Experimental Santa Catalina. INIAP: Quito, Ecuador, 2010; p. 68. [Google Scholar]
- DENAREF (Departamento Nacional de Recursos Fitogenéticos). Banco Nacional de Germoplasma. Estación Experimental Santa Catalina; INIAP: Quito, Ecuador, 2016; p. 34. [Google Scholar]
- Patrimonio Alimentario. Ministerio de Cultura y Patrimonio. Fascículo 3. Oca Tubérculos de la Región Andina. Amaranto/Sangorache Grano de Proteína. Available online: https://www.culturaypatrimonio.gob.ec/wp-content/uploads/downloads/2013/11/3-P.Alimentario-lunes.pdf (accessed on 11 September 2022).
- Michelle, O.F. Innovando en la cocina andina. Saberes y sabores en movimiento. In Recetario con Productos Tradicionales Andinos de la Provincia de Imbabura; Programa de Desarrollo Territorial Rural; Oxfam Italia: Cotacachi, Ecuador, 2015; p. 67. [Google Scholar]
- Suquilanda, M. Producción orgánica de cultivos andinos. Capítulo 9. In Producción Orgánica del Amaranto (Amaranthus caudatus); Suquilanda, M., Ed.; UNOCANC: Cotopaxi, Ecuador, 2011; pp. 118–134. [Google Scholar]
- Ramírez, J. ‘Un siglo de ausencias’: Historia incompleta de la migración ecuatoriana. Mashkana 2021, 12, 47–64. [Google Scholar] [CrossRef]
- Malaghan, S.N.; Revanappa, S.; Ajjappalavar, P.S.; Nagaraja., M.S.; Raghavendra, S. Genetic Variability, Heritability and Genetic Advance in Grain Amaranth (Amaranthus spp.). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1485–1494. [Google Scholar] [CrossRef]
- Nguyen, D.C.; Tran, D.S.; Tran, T.T.H.; Ohsawa, R.; Yoshioka, Y. Genetic diversity of leafy amaranth (Amaranthus tricolor L.) resources in Vietnam. Breed. Sci. 2019, 69, 640–650. [Google Scholar] [CrossRef]
- Nyasulu, M.; Sefasi, A.; Chimzinga, S.; Maliro, M. Agromophological characterisation of Amaranth Accessions from Malawi. Am. J. Plant Sci. 2021, 12, 1528–1542. [Google Scholar] [CrossRef]
- Mazón, N.; Peralta, E.; Rivera, M.; Subía, C.; Tapia, C. Catálogo del Banco de Germoplasma de Amaranto (Amaranthus spp.) de INIAP-Ecuador; Estación Experimental Santa Catalina: Quito, Ecuador, 2003; p. 53. [Google Scholar]
- Tapia, C.; Peralta, E.; Mazón, N. Colección núcleo de amaranto del Banco de Germoplasma del INIAP, Ecuador. In Avances, Desarrollo y Sustentabilidad Agroambiental en Ecuador y Venezuela; En Sandía, L., Rivas, F., Recalde, E., S. Mafla, S., Eds.; ULA/PUCESI: Mérida, Venezuela, 2017; pp. 200–206. [Google Scholar]
- Delgado, H.; Tapia, C.; Borja, E.; Naranjo, E.; Martín, J.P. Phenotypic diversity of Amaranthus quitensis Kunth landraces: A millenary crop of Ecuadorian Andean region. Sci. Agropecu. 2022, 13, 381–393. [Google Scholar] [CrossRef]
- Park, Y.J.; Nishikawa, T.; Matsushima, K.; Minami, M.; Nemoto, K. A rapid and reliable PCR-restriction fragment length polymorphism (RFLP) marker for the identification of Amaranthus cruentus species. Breed. Sci. 2014, 64, 422–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Transue, D.K.; Fairbanks, D.J.; Robison, L.R.; Andersen, W.R. Species identification by RAPD analysis of grain amaranth genetic resources. Crop. Sci. 1994, 34, 1385–1389. [Google Scholar] [CrossRef]
- Lymanskaya, S.V. Estimation of the Genetic Variability of an Amaranth Collection (Amaranthus L.) by RAPD Analysis. Cytol. Genet. 2012, 46, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Akin-Idowu, P.E.; Gbadegesin, M.A.; Orkpeh, U.; Ibitoye, D.O.; Odunola, O.A. Characterization of Grain Amaranth (Amaranthus spp.) Germplasm in South West Nigeria Using Morphological, Nutritional, and Random Amplified Polymorphic DNA (RAPD). Anal. Resour. 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Sun, M. Comparative analysis of phylogenetic relationships of grain amaranths and their wild relatives (Amaranthus;Amaranthaceae) using internal transcribed spacer, amplified fragment length polymorphism, and double-primer fluorescent intersimple sequence repeat markers. Mol. Phylogenetics Evol. 2001, 21, 372–387. [Google Scholar] [CrossRef]
- Wasson, J.J.; Tranel, P.J. Amplified Fragment Length Polymorphism-Based Genetic Relationships Among Weedy Amaranthus Species. J. Hered. 2005, 96, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Raut, V.R.; Dodake, S.S.; Chimote, V.P. Evaluation of Genetic Diversity in Grain Amaranth (Amaranthus hypochondriacus) at Molecular Level using ISSR Markers. Indian J. Agric. Biochem. 2014, 27, 60–65. [Google Scholar]
- Štefúnová, V.; Bezo, M.; Labajová, M.; Senková, S. Genetic analysis of three Amaranth species using ISSR markers. Emir. J. Food Agric. 2014, 26, 35–44. [Google Scholar] [CrossRef]
- Gelotar, M.J.; Dharajiya, D.T.; Solanki, S.D.; Prajapati, N.N.; Tiwari, K.K. Genetic diversity analysis and molecular characterization of grain amaranth genotypes using inter simple sequence repeat (ISSR) markers. Bull Natl. Res. Cent. 2019, 43, 103. [Google Scholar] [CrossRef]
- Suresh, S.; Chung, J.W.; Cho, G.T.; Sung, J.S.; Park, J.H.; Gwag, J.G.; Baek, H.J. Analysis of molecular genetic diversity and population structure in Amaranthus germplasm using SSR markers. Plant Biosyst. 2014, 148, 635–644. [Google Scholar] [CrossRef]
- Maughan, P.J.; Yourstone, S.M.; Jellen, E.N.; Udall, J.A. SNP discovery via genomic reduction, barcoding, and 454-pyrosequencing in amaranth. Plant Genome 2009, 2, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Maughan, P.J.; Smith, S.M.; Fairbanks, D.J.; Jellen, E.N. Development, Characterization, and Linkage Mapping of Single Nucleotide Polymorphisms in the Grain Amaranths (Amaranthus sp.). Plant Genome 2011, 4, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Blair, M.W. Diversity in Grain Amaranths and Relatives Distinguished by Genotyping by Sequencing (GBS). Front. Plant Sci. 2017, 8, 1960. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; Munhoz, C.D. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Mallory, M.A.; Hall, R.V.; McNabb, A.R.; Pratt, D.B.; Jellen, E.N.; Maughan, P.J. Development and characterization of microsatellite markers for the grain amaranths. Crop. Sci. 2008, 48, 1098–1106. [Google Scholar] [CrossRef]
- Lee, J.R.; Hong., G.Y.; Dixit, A.; Chung, J.W.; Ma, K.H.; Lee, J.H.; Kang, H.K.; Cho, H.Y.; Gwang, J.G.; Park, Y.J. Characterization of microsatellite loci developed for Amaranthus hypochondriacus and their cross-amplification in wild species. Conserv. Genet. 2008, 9, 243–246. [Google Scholar] [CrossRef]
- Khaing, A.A.; Moe, K.T.; Chung, J.W.; Baek, H.J.; Park, Y.J. Genetic diversity and population structure of the selected core set in Amaranthus using SSR markers. Plant Breed. 2013, 132, 165–173. [Google Scholar] [CrossRef]
- Wang, X.Q.; Park, J.Y. Comparison of genetic diversity among amaranth accessions from South and Southeast Asia using SSR markers. Korean J. Crop. Sci. 2013, 21, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Nieto, C. El Cultivo de Amaranto Amaranthus spp. Una Alternativa Agronómica para Ecuador; Publicación Miscelánea No.52 Estación Experimental Santa Catalina: Quito, Ecuador, 1989; p. 25. [Google Scholar]
- Peakal, R.; Smouse, P. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.A. Blood samples: Probability of discrimination. J. Forensic. Sci. Soc. 1972, 12, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Martín del Puerto, M.; Martínez García, F.; Mohanty, A.; Martín, J.P. Genetic Diversity in Relict and Fragmented Populations of Ulmus glabra Hudson in the Central System of the Iberian Peninsula. Forests 2017, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A. The Use of Multiple Measurements in Taxonomy Problems. Ann. Eugen. 1936, 7, 179–188. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Amer. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction sites. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. Available online: http://www.alz.org/what-is-dementia.asp (accessed on 20 October 2022). [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Worthington, M.; Soleri, D.; Aragón-Cuevas, F.; Gepts, P. Genetic composition and spatial distribution of farmer-managed Phaseolus bean plantings: An example from a village in Oaxaca, Mexico. Crop. Sci. 2012, 52, 1721–1735. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy System (Ver. 2.2); Exeter Publishing, Ltd.: New York, NY, USA, 2008. [Google Scholar]
- Espitia, R.E.; Mapes, S.C.; Escobedo, L.D.; De la O. Olán, M.; Rivas, V.P.; Martínez, T.G.; Cortés, E.L.; Hernández, C.J. Conservación y uso de Los Recursos Genéticos de Amaranto en México; INIFAP, Centro de Investigación Regional Centro: Celaya, México, 2010; p. 200. [Google Scholar]
- Basantes, F.; Aragón, J.P.; Albuja, M. Cultivos andinos de importancia agro productiva y comercial en la zona 1 de Ecuador; Editorial UTN: Ibarra, Ecuador, 2022; p. 192. [Google Scholar]
- Informe Final. Proyecto Recolección de varios cultivos andinos en Ecuador; Instituto Nacional de Investigaciones Agropecuarias (INIAP): Quito, Ecuador, 1985; p. 100. [Google Scholar]
- Jurado, E.O. Estudio de la Producción y Comercialización del Amaranto (Amaranthus sp) en la Provincia de Imbabura; Trabajo de Grado; Facultad de Universidad Técnica del Norte, Ingeniería en Ciencias Agropecuarias y Ambientales: Ibarra, Ecuador, 2019. [Google Scholar]
- Cevallos, M.; Urdaneta, F.; Jaimes, E.; Rodriguez, M. Transición agroecológica de los sistemas de producción agrícola de la provincia de Imbabura Ecuador. Rev. Fac Agron. Luz. 2020, 37, 69–94. [Google Scholar]
- Monteros, C.; Nieto, C.; Caicedo, C.; Rivera, M.; Vimos, C. INIAP-ALEGRÍA; Primera variedad mejorada de amaranto para la sierra ecuatoriana, Boletín Divulgativo No.246, Programa de Cultivos Andinos; Estación Experimental Santa Catalina, INIAP: Quito, Ecuador, 1994; p. 24. [Google Scholar]
- Zimmerer, K.S. Geographies of seed networks for food plants (potato, ulloco) and approaches to agrobiodiversity conservation in the Andean countries. Soc. Natur. Resour. 2003, 16, 583–601. [Google Scholar] [CrossRef]
- Salazar, J.; De Lourdes, M.; Gutiérrez, B.; Torres, A.F. Molecular characterization of Ecuadorian quinoa (Chenopodium quinoa Willd.) diversity: Implications for conservation and breeding. Euphytica 2019, 215, 60. [Google Scholar] [CrossRef]
- Peralta, E.; Mazón, N.; Murillo, Á.; Villacrés, E.; Rivera, M. Catalogó de Variedades Mejoradas de Granos Andino: Chocho, Quinua, Amaranto, Sangorache, Para la Sierra Ecuatoriana; Publicación Miscelánea No. 151.Tercera Edición, Programa Nacional de Leguminosas y Granos Andinos, Estación; Experimental Santa Catalina INIAP: Quito, Ecuador, 2013; p. 63. [Google Scholar]
- González, M.S. Estudio de Flujo de Genes en Quinua (Chenopodium quinoa w.) en Campo de Agricultores Mediante el uso de Marcadores Microssatélites; Trabajo de Grado; Departamento de Ciencias de la Vida Ingeniería en Biotecnología, Escuela Politécnica del Ejercito: Pichincha, Ecuador, 2009. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No._Locus a | Forward Sequence (5′-3′) Reverse Sequence (5′-3′) | Repeat Motif | Fluorescent Dye b | Annealing Temp. (°C) |
|---|---|---|---|---|
| 1_GB-AMM-051 | GAGGAGACTTGGTGGCCT TCGGGAGCAATGTAGCAC | (AGA)5 | FAM | 50 |
| 2_GB-AMM-099 | AAATTGACAATGCGCAGC TTCCTCACCAAAATTGCC | (TCA)12(TCT)5 | FAM | 50 |
| 3_GB-AMM-136 | TCAGCAAAACATGATCAACAA GTTGCTGCATTGGTGGTT | (GAA)6(CCA)6 | CFR590 | 50 |
| 4_AHAAC011 | CCGTCTGTGCTGTATTGAGG GGCCACTTGGGTTTATTCCT | (GTT)8 | FAM | 50 |
| 5_AHAAC021 | GAGTTATGGCCGAATTTCCA TTGGTGTTGTTCAACATTTGG | (CAA)9 | CFG540 | 50 |
| 6_AHAAT030 | CCAGATGCCAGATGTGCTTA CCAAACAAGGTCGATTTCAGA | (ATT)11 | Q570 | 52 |
| 7_AHAAT051 | TGTAACACTGCGCTACAAATCA CCCTCAGAGTTTCCTTCACC | (AAC)7AGC(AAT)AGT(AAT)20 | CFR590 | 50 |
| 8_AHAAT063 | TCGGAAATTAGTCGGAGGTTT CGATGACAATTATGTAACCCAATG | (TTA)25 | CFG540 | 50 |
| 9_AHAC062 | GGCTCCCAAGTCACAGTGTT TCATCTTTATCGTTGATTCGTTTC | (AC)11(AGACACAC)2(AC)5 (AGACACAC)4(AC)4 | Q570 | 52 |
| No._Locus | Na | Ne | Ng | Ho | He | I | C | D |
|---|---|---|---|---|---|---|---|---|
| 1_GB-AMM-051 | 4 | 1.149 | 4 | 0.013 | 0.129 | 0.278 | 0.864 | 0.136 |
| 2_GB-AMM-099 | 2 | 1.140 | 3 | 0.026 | 0.123 | 0.243 | 0.852 | 0.148 |
| 3_GB-AMM-136 | 4 | 1.150 | 5 | 0.013 | 0.130 | 0.292 | 0.863 | 0.137 |
| 4_AHAAC011 | 2 | 1.118 | 3 | 0.007 | 0.106 | 0.216 | 0.888 | 0.112 |
| 5_AHAAC021 | 3 | 1.149 | 4 | 0.013 | 0.129 | 0.273 | 0.864 | 0.136 |
| 6_AHAAT030 | 4 | 1.164 | 4 | 0.013 | 0.141 | 0.295 | 0.853 | 0.147 |
| 7_AHAAT051 | 6 | 1.166 | 5 | 0.026 | 0.143 | 0.344 | 0.840 | 0.160 |
| 8_AHAAT063 | 6 | 1.175 | 5 | 0.007 | 0.149 | 0.358 | 0.851 | 0.149 |
| 9_AHAC062 | 5 | 1.183 | 6 | 0.007 | 0.155 | 0.373 | 0.839 | 0.161 |
| Mean | 4.0 | 1.155 | 4.3 | 0.014 | 0.134 | 0.297 | - | - |
| Cumulative | 36 | - | 38 | - | - | - | 0.249 | 0.751 |
| Provinces (Collection) | % P | G | Na | Ne | Ho | He | I |
|---|---|---|---|---|---|---|---|
| Imbabura (A) | 100 | 6 | 2.11 a | 1.770 a | 0.034 a | 0.435 a | 0.637 a |
| Tungurahua (A) | 33 | 4 | 1.33 b | 1.018 b | 0.009 a,b | 0.017 b,c | 0.039 b |
| Cañar (A) | 44 | 5 | 1.44 b | 1.050 b | 0.023 a,b | 0.043 c | 0.087 b |
| Mean Collection A | 59.3 | 5.0 | 1.63 | 1.279 | 0.022 | 0.165 | 0.254 |
| Imbabura (B) | 100 | 8 | 3.67 c | 1.417 c | 0.029 a | 0.292 d | 0.555 c |
| Tungurahua (B) | 11 | 2 | 1.11 b | 1.007 b | 0.000 b | 0.007 b | 0.016 b |
| Cañar (B) | 11 | 2 | 1.11 b | 1.008 b | 0.000 b | 0.007 b | 0.017 b |
| Mean Collection B | 40.7 | 4.0 | 1.96 | 1.144 | 0.010 | 0.102 | 0.196 |
| Collection A | Collection B | ||||
|---|---|---|---|---|---|
| Imbabura | Tungurahua | Cañar | Imbabura | Tungurahua | Cañar |
| G1 (6) | G1 (23) | G1 (15) | G1 (25) | G1 (30) | G1 (28) |
| G2 (3) | G2 (3) | ||||
| G3 (1) | G3 (1) | G3 (1) | |||
| G4 (1) | G4 (1) | ||||
| G5 (1) | G5 (1) | ||||
| G6 (1) | |||||
| G7 (1) | |||||
| G8 (1) | |||||
| G9 (1) | |||||
| G10 (1) | |||||
| G11 (1) | |||||
| G12 (1) | |||||
| G13 (1) | |||||
| G14 (1) | |||||
| G15 (1) | |||||
| G16 (1) | |||||
| G17 (1) | |||||
| Imbabura_A | Tungurahua_A | Cañar_A | Imbabura_B | Tungurahua_B | Cañar_B | |
|---|---|---|---|---|---|---|
| Imbabura_A | --- | |||||
| Tungurahua_A | 0.094 | --- | ||||
| Cañar_A | 0.095 | 0.002 | --- | |||
| Imbabura_B | 0.037 | 0.015 | 0.016 | --- | ||
| Tungurahua_B | 0.095 | 0.000 | 0.001 | 0.016 | --- | |
| Cañar_B | 0.095 | 0.000 | 0.001 | 0.016 | 0.000 | --- |
| Na | Ne | Ho | He | I | |
|---|---|---|---|---|---|
| Cluster 1 | 2.00 a | 1.018 a | 0.009 a | 0.018 a | 0.049 a |
| Cluster 2 | 2.89 b | 1.401 b | 0.089 b | 0.264 b | 0.510 b |
| Mean | 2,44 | 1,210 | 0,049 | 0,141 | 0,279 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, H.; Martín, J.P. Genetic Diversity of Black Amaranth (Amaranthus quitensis Kunth) Landraces of Ecuadorian Highlands: Association Genotypes—Color Morphotypes. Agriculture 2023, 13, 34. https://doi.org/10.3390/agriculture13010034
Delgado H, Martín JP. Genetic Diversity of Black Amaranth (Amaranthus quitensis Kunth) Landraces of Ecuadorian Highlands: Association Genotypes—Color Morphotypes. Agriculture. 2023; 13(1):34. https://doi.org/10.3390/agriculture13010034
Chicago/Turabian StyleDelgado, Hipatia, and Juan Pedro Martín. 2023. "Genetic Diversity of Black Amaranth (Amaranthus quitensis Kunth) Landraces of Ecuadorian Highlands: Association Genotypes—Color Morphotypes" Agriculture 13, no. 1: 34. https://doi.org/10.3390/agriculture13010034