
High-Throughput Phenotyping of Cross-Sectional Morphology to Assess Stalk Mechanical Properties in Sorghum
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
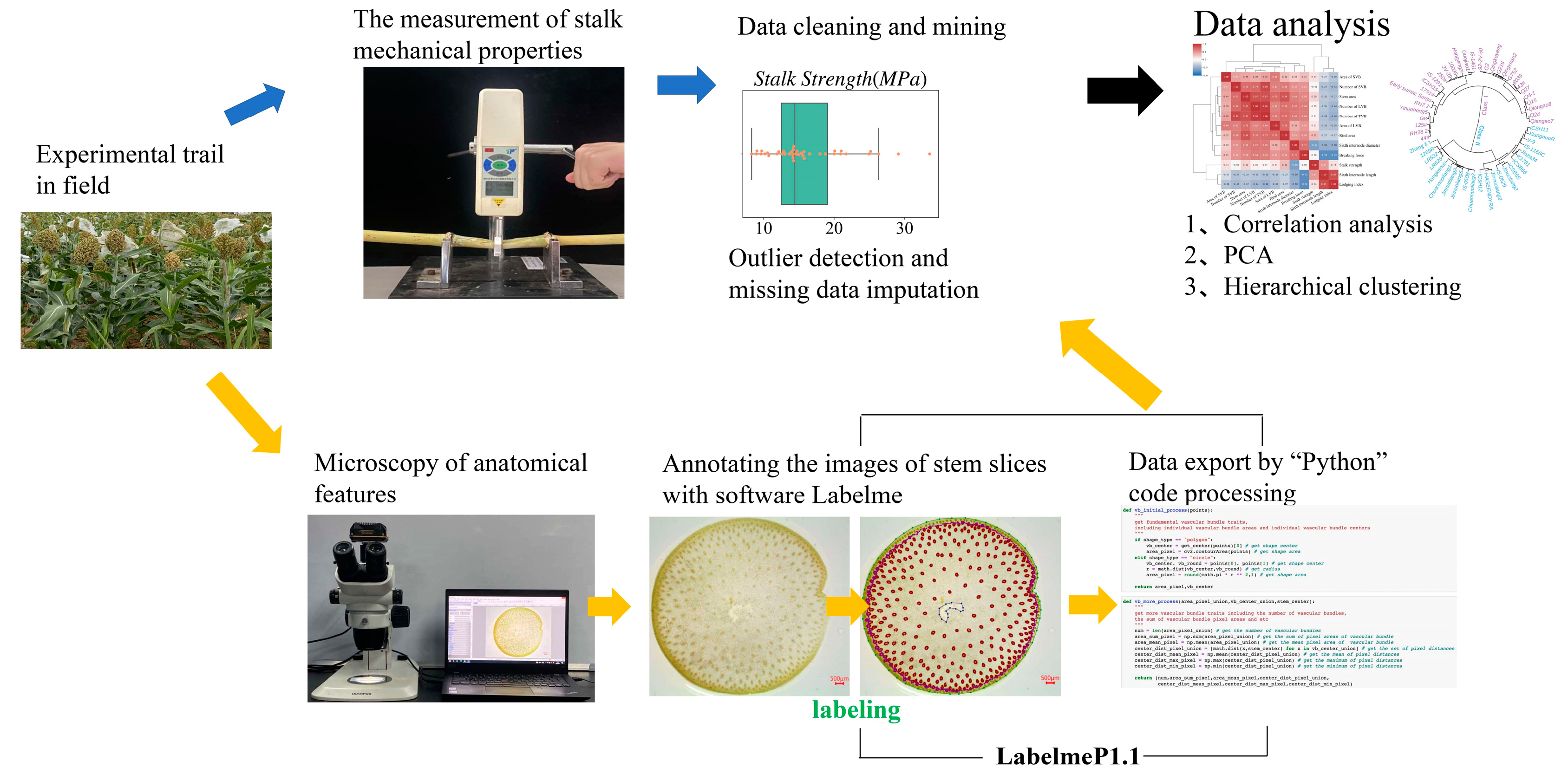
2.2. The Measurement of Plant Morphological Traits and Stem Mechanical Properties
2.3. Microscopy of Anatomical Features
2.4. Annotating the Images of Stem Slices from Sorghum Stalks
2.5. “Python” Code Processing to Extract Stem Anatomical Parameters
2.6. Statistical Analysis and Visualization Methods
3. Results
3.1. Statistics of the Stem Mechanical Properties of Sorghum Germplasm Resources
3.2. Statistics of Stem Cross-Sectional Parameters of Sorghum Germplasm Resources
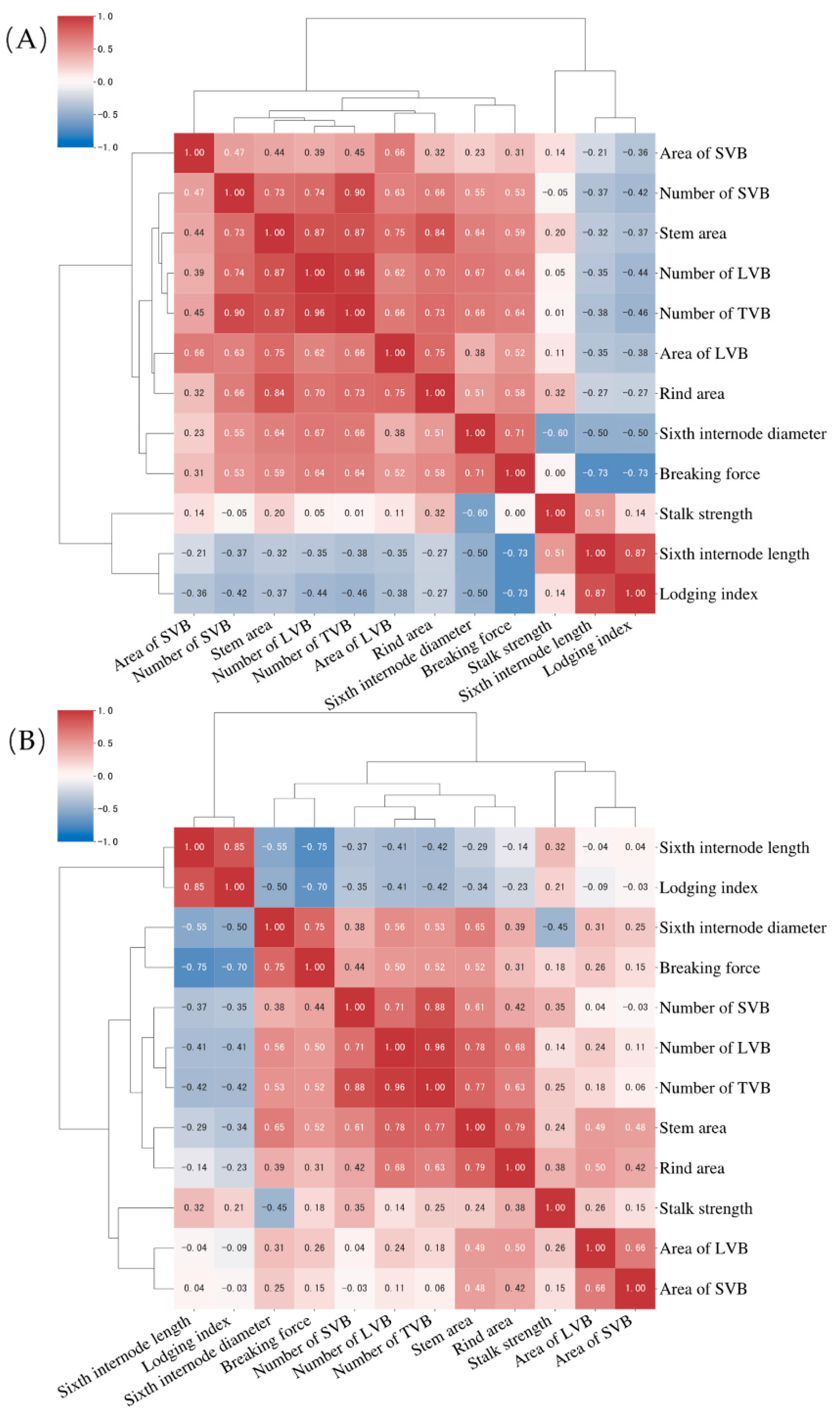
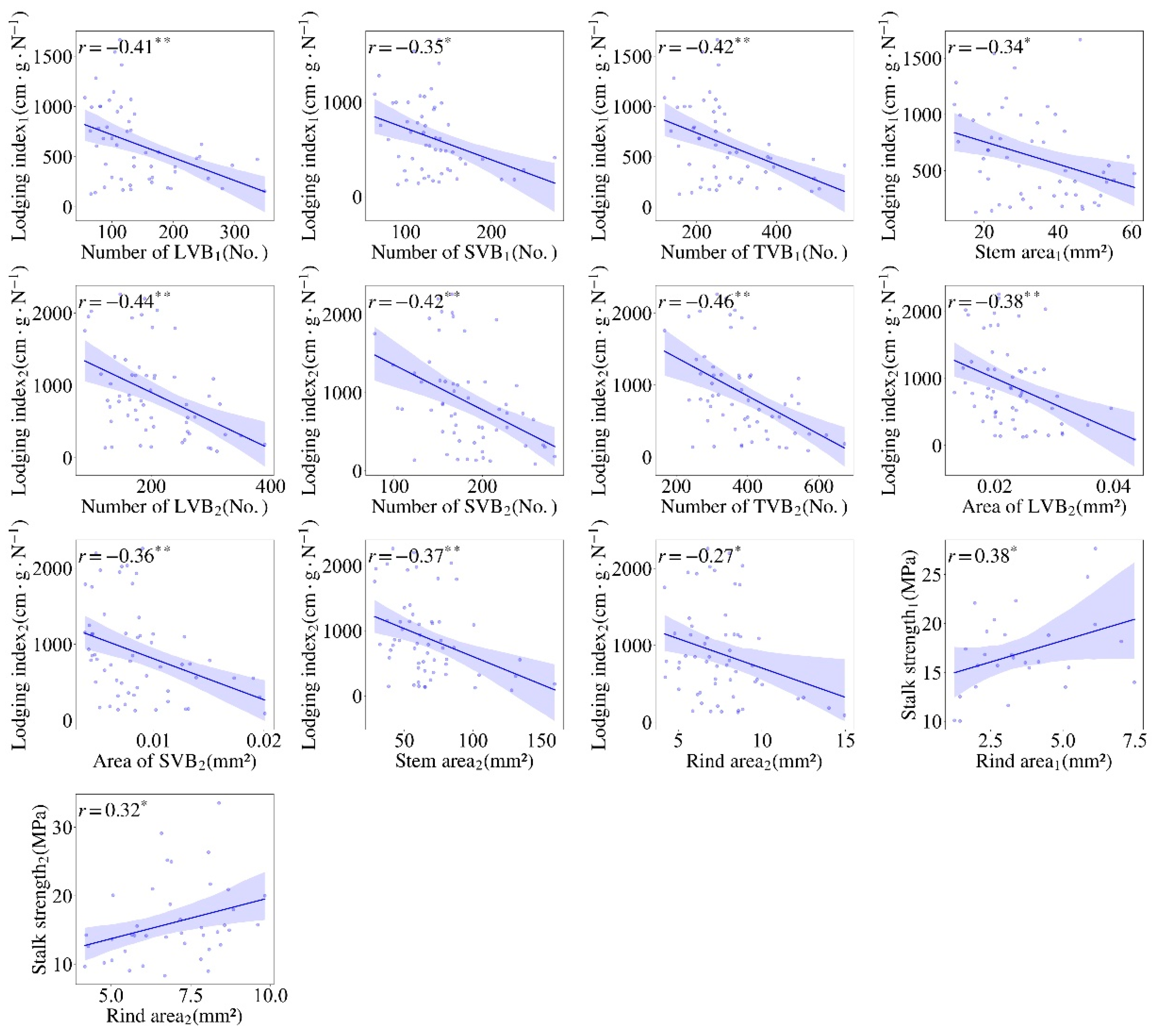
3.3. Correlation of Stalk Mechanical Properties with the Stem Cross-Sectional Parameters in Sorghum Germplasm
3.4. The Principal Components Analysis of the Stalk Mechanical Properties and Stem Cross-Sectional Parameters
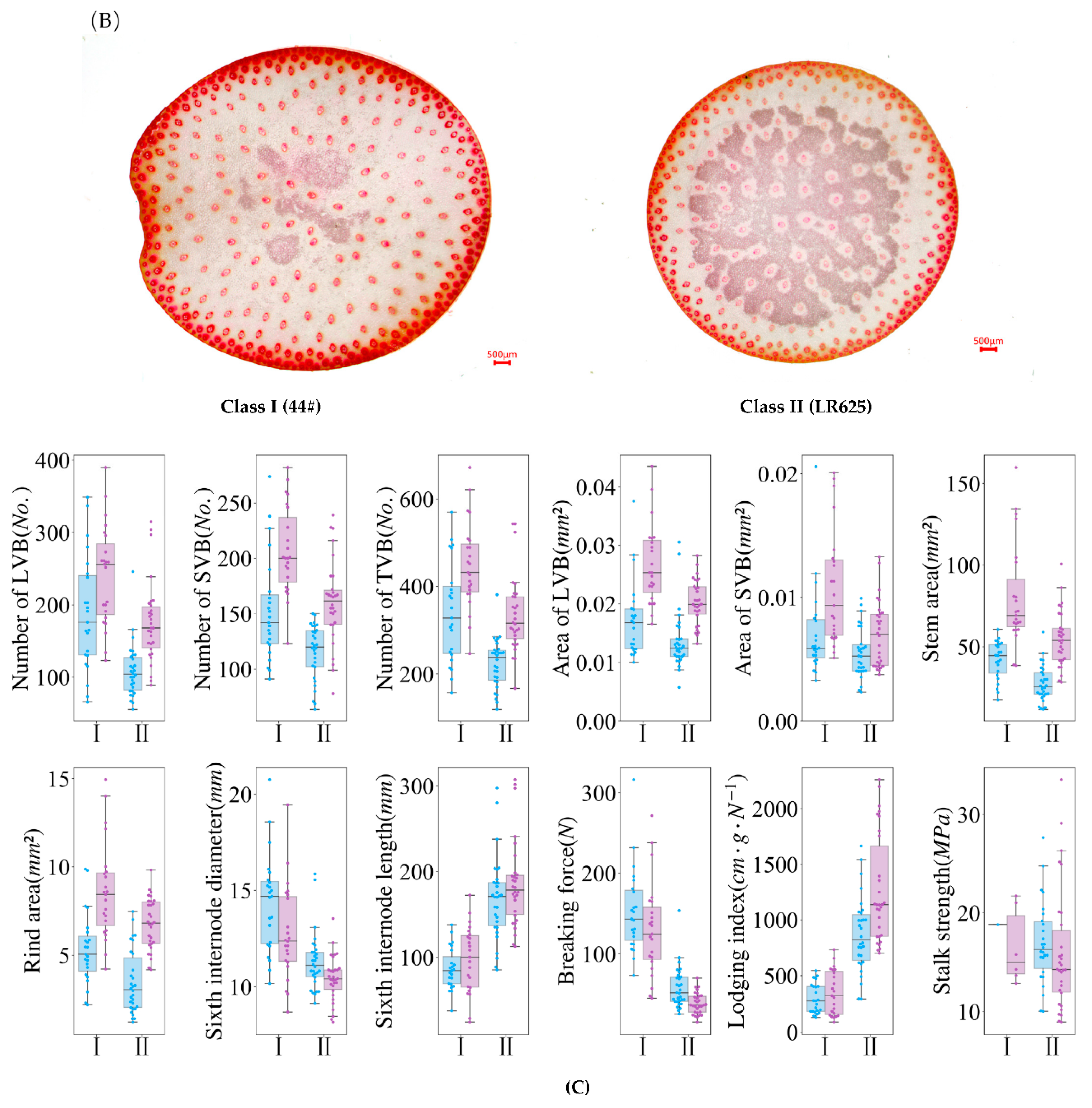
3.5. Two Contrast Categories Were Obtained and Compared
4. Discussion
4.1. The Development of a Novel Procedure Allowed the High-Throughput Extraction of the Anatomical Parameters
4.2. The Relationship of the Traits and Parameters of the Sorghum Stem and the Inspiration for Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berry, P.; Sylvester-Bradley, R.; Berry, S. Ideotype design for lodging-resistant wheat. Euphytica 2007, 154, 165–179. [Google Scholar] [CrossRef]
- Setter, T.; Laureles, E.; Mazaredo, A. Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis. Field Crops Res. 1997, 49, 95–106. [Google Scholar] [CrossRef]
- Berry, P.; Spink, J. Predicting yield losses caused by lodging in wheat. Field Crops Res. 2012, 137, 19–26. [Google Scholar] [CrossRef]
- Rutto, L.K.; Xu, Y.; Brandt, M.; Ren, S.; Kering, M.K. Juice, ethanol, and grain yield potential of five sweet sorghum (Sorghum bicolor [L.] Moench) cultivars. J Sustain. Bioenergy Syst. 2013, 3, 113. [Google Scholar] [CrossRef] [Green Version]
- Byrt, C.S.; Grof, C.P.; Furbank, R.T. C4 plants as biofuel feedstocks: Optimising biomass production and feedstock quality from a lignocellulosic perspective. J. Integr. Plant Biol. 2011, 53, 120–135. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Li, Z.; Leng, C.; Lu, C.; Luo, H.; Liu, Y.; Wu, X.; Liu, Z.; Shang, L.; Jing, H.C. Sorghum breeding in the genomic era: Opportunities and challenges. Theor. Appl. Genet. 2021, 134, 1899–1924. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Sayre, K.; Kaul, J.; Narang, R. Lodging behavior and yield potential of spring wheat (Triticum aestivum L.): Effects of ethephon and genotypes. Field Crops Res. 2004, 87, 207–220. [Google Scholar] [CrossRef]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shi, Y.-h.; Wang, C.-y.; Yin, Y.-p.; Ning, T.-y.; Shi, C.-y.; Li, Y.; Wang, Z.-l. Effects of Nitrogen and PP333 Application on the Lignin Synthesis of Stem in Relation to Lodging Resistance of Wheat (in chinese). Sci. Agric. Sin. 2011, 44, 3529–3536. [Google Scholar]
- Li, G.-h.; Zhong, X.-h.; Tian, K.; Huang, N.-r.; Pan, J.-f.; He, T.-h. Effect of Nitrogen Application on Stem Lodging Resistance of Rice and Its Morphological and Mechanical Mechanisms (in chinese). Sci. Agric. Sin. 2013, 46, 1323–1334. [Google Scholar]
- Liu, C.; Zheng, S.; Gui, J.; Fu, C.; Yu, H.; Song, D.; Shen, J.; Qin, P.; Liu, X.; Han, B. Shortened basal internodes encodes a gibberellin 2-oxidase and contributes to lodging resistance in rice. Mol. Plant 2018, 11, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, T.; Ishimaru, K. Identification and functional analysis of a locus for improvement of lodging resistance in rice. Plant Physiol. 2004, 134, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Hai, L.; Guo, H.; Xiao, S.; Jiang, G.; Zhang, X.; Yan, C.; Xin, Z.; Jia, J. Quantitative trait loci (QTL) of stem strength and related traits in a doubled-haploid population of wheat (Triticum aestivum L.). Euphytica 2005, 141, 1–9. [Google Scholar] [CrossRef]
- Huang, X.; Cloutier, S.; Lycar, L.; Radovanovic, N.; Humphreys, D.; Noll, J.; Somers, D.; Brown, P. Molecular detection of QTLs for agronomic and quality traits in a doubled haploid population derived from two Canadian wheats (Triticum aestivum L.). Theor. Appl. Genet. 2006, 113, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H. The expression of caffeic acid 3-O-methyltransferase in two wheat genotypes differing in lodging resistance. J. Exp. Bot. 2009, 60, 2763–2771. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Ran, W.; Li, L.; Chen, J.; Zheng, H.; Ma, Y.; Mao, J.; Zheng, B.; Shao, R. Correlation and path analysis of lodging resistance with maize stem characters (in chinese). J. Henan Agric. Univ. 2016, 50, 167–170. [Google Scholar]
- Hu, W.; Zhang, Y.e.; Zhao, H.; Wang, X.; Cao, T.; Cao, Y.; Chen, Y.; Yang, J. Comparison and Analysis of the Methods for Evaluating Wheat Lodging Resistance (in chinese). Acta Agric. Boreali-Occident. Sin. 2018, 27, 1780–1788. [Google Scholar]
- Muhammad, A.; Hao, H.; Xue, Y.; Alam, A.; Bai, S.; Hu, W.; Sajid, M.; Hu, Z.; Samad, R.A.; Li, Z.; et al. Survey of wheat straw stem characteristics for enhanced resistance to lodging. Cellulose 2020, 27, 2469–2484. [Google Scholar] [CrossRef]
- Forell, G.V.; Robertson, D.; Lee, S.Y.; Cook, D.D. Preventing lodging in bioenergy crops: A biomechanical analysis of maize stalks suggests a new approach. J. Exp. Bot. 2015, 66, 4367–4371. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Ma, H.; Zhang, P.; Ren, L.; Yang, X.; Yao, G.; Zhang, P.; Zhou, M. Inheritance of stem strength and its correlations with culm morphological traits in wheat (Triticum aestivum L.). Can. J. Plant Sci. 2011, 91, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Kong, E.; Liu, D.; Guo, X.; Yang, W.; Sun, J.; Li, X.; Zhan, K.; Cui, D.; Lin, J.; Zhang, A. Anatomical and chemical characteristics associated with lodging resistance in wheat. Crop J. 2013, 1, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhang, M.; Guo, K.; Hu, Z.; Zhang, R.; Feng, Y.; Yi, X.; Zou, W.; Wang, L.; Wu, C. High-level hemicellulosic arabinose predominately affects lignocellulose crystallinity for genetically enhancing both plant lodging resistance and biomass enzymatic digestibility in rice mutants. Plant Biotechnol. J. 2015, 13, 514–525. [Google Scholar] [CrossRef]
- Li, F.; Xie, G.; Huang, J.; Zhang, R.; Li, Y.; Zhang, M.; Wang, Y.; Li, A.; Li, X.; Xia, T. Os CESA 9 conserved-site mutation leads to largely enhanced plant lodging resistance and biomass enzymatic saccharification by reducing cellulose DP and crystallinity in rice. Plant Biotechnol. J. 2017, 15, 1093–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, D.; Song, Y.; Wu, Q.; Ma, C.; Zhao, J.; Wan, Y.; Zhao, G. Relationship between stem characteristics and lodging resistance of Tartary buckwheat (Fagopyrum tataricum). Plant Prod. Sci. 2019, 22, 202–210. [Google Scholar] [CrossRef]
- Wang, X.; Mace, E.; Tao, Y.; Cruickshank, A.; Hunt, C.; Hammer, G.; Jordan, D. Large-scale genome-wide association study reveals that drought-induced lodging in grain sorghum is associated with plant height and traits linked to carbon remobilisation. Theor. Appl. Genet. 2020, 133, 3201–3215. [Google Scholar] [CrossRef]
- Qunying, W.; Changhao, H. Studies on the anatomical structures of the stalks of maize with different resistance to lodging (in chinese). Acta Agron. Sin. 1991, 17, 72–75. [Google Scholar]
- Zhou, H.-Y.; Jiang, Y.-F.; Yang, M.-C.; Cheng, W.-D.; Qin, L.-Q.; Xie, X.-D.; Xie, H.-X.; Qin, B.-X.; Wang, L.-Q. Evaluation of Stalk Strength, Vascular Bundle and Fiber in Maize. J. Plant Genet. Resour. 2022, 1–8. (In Chinese) [Google Scholar] [CrossRef]
- Robertson, D.J.; Julias, M.; Lee, S.Y.; Cook, D.D. Maize stalk lodging: Morphological determinants of stalk strength. Crop Sci. 2017, 57, 926–934. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, C.J.; McMahan, C.; Seegmiller, W.; Cook, D.D.; Robertson, D.J. Integrated Puncture Score: Force–displacement weighted rind penetration tests improve stalk lodging resistance estimations in maize. Plant Methods 2020, 16, 113. [Google Scholar] [CrossRef]
- Stubbs, C.J.; Seegmiller, K.; McMahan, C.; Sekhon, R.S.; Robertson, D.J. Diverse maize hybrids are structurally inefficient at resisting wind induced bending forces that cause stalk lodging. Plant Methods 2020, 16, 67. [Google Scholar] [CrossRef]
- van Heerden, P.D.; Donaldson, R.A.; Watt, D.A.; Singels, A. Biomass accumulation in sugarcane: Unravelling the factors underpinning reduced growth phenomena. J. Exp. Bot. 2010, 61, 2877–2887. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.; Wu, Y.; Wang, X.; Yu, X.; Meng, X.; Zhang, J.; Ma, X. Differences of Stem Main Inclusion Analysis of Wheat Varieties with Different Height and Lodging Resistance (in chinese). J. Triticeae Crops 2017, 37, 1187–1194. [Google Scholar]
- Gomez, F.E.; Mullet, J.E.; Muliana, A.H.; Niklas, K.J.; Rooney, W.L. The genetic architecture of biomechanical traits in sorghum. Crop Sci. 2020, 60, 82–99. [Google Scholar] [CrossRef]
- Tanaka, K.; Murata, K.; Yamazaki, M.; Onosato, K.; Miyao, A.; Hirochika, H. Three distinct rice cellulose synthase catalytic subunit genes required for cellulose synthesis in the secondary wall. Plant Physiol. 2003, 133, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Zhou, Y. Rice brittleness mutants: A way to open the ‘Black Box’of monocot cell wall biosynthesis free access. J. Integr. Plant Biol. 2011, 53, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xia, T.; Li, G.; Li, X.; Li, Y.; Wang, Y.; Wang, Y.; Chen, Y.; Xie, G.; Bai, F.-W. Overproduction of native endo-β-1, 4-glucanases leads to largely enhanced biomass saccharification and bioethanol production by specific modification of cellulose features in transgenic rice. Biotechnol. Biofuels 2019, 12, 11. [Google Scholar] [CrossRef]
- Oduntan, Y.A.; Stubbs, C.J.; Robertson, D.J. High throughput phenotyping of cross-sectional morphology to assess stalk lodging resistance. Plant Methods 2022, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Peng, S.; Visperas, R.M.; Ereful, N.; Bhuiya, M.S.U.; Julfiquar, A. Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem. Field Crops Res. 2007, 101, 240–248. [Google Scholar] [CrossRef]
- Gomez, F.E.; Carvalho, G., Jr.; Shi, F.; Muliana, A.H.; Rooney, W.L. High throughput phenotyping of morpho-anatomical stem properties using X-ray computed tomography in sorghum. Plant Methods 2018, 14, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, A.-N.; Yan, J.-Q.; Hai-Bo, L.; Zhao, H.-C.; Huang, Z.-H. Response of Stem Microstructure of Different Spring Maize Varieties to Bending Strength. J. Maize Sci. 2020, 28, 53–59. (In Chinese) [Google Scholar]
- Niklas, K.J. Plant Biomechanics: An Engineering Approach to Plant Form and Function; University of Chicago Press: Chicago, IL, USA, 1992. [Google Scholar]
- Crook, M.; Ennos, A. Stem and root characteristics associated with lodging resistance in four winter wheat cultivars. J. Agric. Sci. 1994, 123, 167–174. [Google Scholar] [CrossRef]
- Fan, C.; Feng, S.; Huang, J.; Wang, Y.; Wu, L.; Li, X.; Wang, L.; Tu, Y.; Xia, T.; Li, J. AtCesA8-driven OsSUS3 expression leads to largely enhanced biomass saccharification and lodging resistance by distinctively altering lignocellulose features in rice. Biotechnol. Biofuels 2017, 10, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amano, T.; Zhu, Q.; Wang, Y.; Inoue, N.; Tanaka, H. Case studies on high yields of paddy rice in Jiangsu Province, China: I. Characteristics of grain production. Jpn. J. Crop Sci. 1993, 62, 267–274. [Google Scholar]
- Zhu, L.; Zhong, D.; Xu, J.; Yu, S.; Li, Z. Differential expression of lodging resistance related QTLs in rice (Oryza sativa L.). Plant Sci. 2008, 175, 898–905. [Google Scholar] [CrossRef]
- Shen, Y.; Ma, F.; Wang, M.; Li, X.; Zhang, M.; Huang, J. Accurate Evaluation and Mechanism Analysis of Mechanical Strength of Sugarcane Stalk. Chin. J. Trop. Crops 2021, 43, 207. (In Chinese) [Google Scholar]
- Lemloh, M.-L.; Pohl, A.; Weber, E.; Zeiger, M.; Bauer, P.; Weiss, I.M.; Schneider, A.S. Structure-property relationships in mechanically stimulated Sorghum bicolor stalks. Bioinspired Mater. 2014, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gomez, F.E.; Muliana, A.H.; Rooney, W.L. Predicting Stem Strength in Diverse Bioenergy Sorghum Genotypes. Crop Sci. 2018, 58, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Yan, D. Genome-Wide Association Analysis of Lodging-Related Traits in Wheat. Master’s Thesis, Shandong Agricultural University, Taian, China, 2019. [Google Scholar]
- Nan, M. Study Expression of Genes in Stem Lignin Synthesis and Physiological Mechanism of Lodging Resistance in Oat. Ph.D. Thesis, Gansu Agricultural University, Lanzhou, China, 2021. [Google Scholar]
- Shah, L.; Yahya, M.; Shah, S.M.A.; Nadeem, M.; Ali, A.; Ali, A.; Wang, J.; Riaz, M.W.; Rehman, S.; Wu, W.; et al. Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. Int. J. Mol. Sci. 2019, 20, 4211. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Genotype | Source | No. | Genotype | Source | No. | Genotype | Source |
|---|---|---|---|---|---|---|---|---|
| 1 | Chuannuoliang 1 | Sichuan | 21 | 43# | Guizhou | 41 | 1791# | Liaoning |
| 2 | Chuannuoliang 2 | Sichuan | 22 | 44# | Guizhou | 42 | LR622 | Liaoning |
| 3 | Guojiao 1 | Sichuan | 23 | 92-2V-50 | Guizhou | 43 | LR625 | Liaoning |
| 4 | Hongyingzi | Sichuan | 24 | G2 | Guizhou | 44 | Jian LR | Liaoning |
| 5 | Jinuoliang 2 | Sichuan | 25 | G216 | Guizhou | 45 | Zheng♂1 | North China |
| 6 | Jinnuoliang 2 | Sichuan | 26 | G752 | Guizhou | 46 | Early sumac Sorgo | America |
| 7 | Jinnuoliang 5 | Sichuan | 27 | G8 | Guizhou | 47 | HAGEENDYRA | America |
| 8 | Jinnuoliang 9 | Sichuan | 28 | RH7.1 | Guizhou | 48 | Hegari #750 | America |
| 9 | Jinza 34 | Sichuan | 29 | RH28.2 | Guizhou | 49 | IS-0508 | America |
| 10 | Qingkeyang | Sichuan | 30 | V-9 | Guizhou | 50 | IS-0829 | America |
| 11 | Yinuohong 5 | Sichuan | 31 | Q1 | Guizhou | 51 | IS-1166C | America |
| 12 | 10098 | Sichuan | 32 | Q4-1 | Guizhou | 52 | IS-1291 | America |
| 13 | Hongkenuo | Guizhou | 33 | Q14-3 | Guizhou | 53 | IS-1461 | America |
| 14 | Qingxuan 2 | Guizhou | 34 | Q15 | Guizhou | 54 | ICSB55 | India |
| 15 | Qiangao 7 | Guizhou | 35 | Q24 | Guizhou | 55 | ICSB56 | India |
| 16 | Qiangao 8 | Guizhou | 36 | Q27 | Guizhou | 56 | ICSH11 | India |
| 17 | 8039 | Guizhou | 37 | K1781 | Jilin | 57 | ICSH12 | India |
| 18 | 125# | Guizhou | 38 | Xiangnuo R | Jilin | 58 | ICSH15 | India |
| 19 | 260# | Guizhou | 39 | Nuo 3 | Jilin | |||
| 20 | 2V-29 | Guizhou | 40 | 1269R | Liaoning |
| Traits | First Season of 2021 | Second Season of 2021 | r between Two Seasons | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | CV (%) | Skew | Kurt | Mean | Min | Max | CV (%) | Skew | Kurt | ||
| Plant height (cm) | 177.6 | 98.0 | 261.8 | 25.2 | 0.0 | −1.0 | 165.3 | 87.7 | 262.3 | 25.3 | 0.3 | −0.4 | 0.86 ** |
| Plant fresh weight (g) | 253 | 125.3 | 607.0 | 40.5 | 1.6 | 2.4 | 257.8 | 146.7 | 556.7 | 34.1 | 1.7 | 3.0 | 0.54 ** |
| Sixth internode length (cm) | 132.8 | 38.0 | 297.5 | 42.9 | 0.7 | 0.4 | 151.1 | 25.0 | 307.3 | 44.5 | 0.5 | 0.2 | 0.85 ** |
| Sixth internode diameter (cm) | 12.6 | 9.2 | 20.8 | 19.9 | 1.1 | 0.9 | 11.5 | 8.2 | 19.4 | 18.4 | 1.3 | 2.6 | 0.78 ** |
| Breaking force (N) | 99.2 | 25.8 | 316.2 | 62.1 | 1.2 | 1.6 | 76.4 | 15.9 | 271.4 | 76.8 | 1.5 | 1.8 | 0.82 ** |
| Stalk strength (MPa) | 17.0 | 10.0 | 27.6 | 24.4 | 0.6 | 0.6 | 16.0 | 8.3 | 33.5 | 36.0 | 1.2 | 1.2 | 0.8 ** |
| Lodging index (cm·g·N−1) | 620.0 | 129 | 1644.1 | 63.0 | 0.7 | −0.1 | 886.8 | 87.9 | 2260.5 | 68.8 | 0.7 | −0.4 | 0.48 * |
| Traits | First Season of 2021 | Second Season of 2021 | r between Two Seasons | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | CV (%) | Skew | Kurt | Mean | Min | Max | CV (%) | Skew | Kurt | ||
| Number of large vascular bundles (No.) | 143.0 | 56.0 | 349.0 | 49.4 | 1.3 | 1.1 | 204.0 | 89.0 | 390.0 | 33.4 | 0.6 | −0.2 | 0.47 ** |
| Number of small vascular bundles (No.) | 132.0 | 64.0 | 274.0 | 30.7 | 1.3 | 2.8 | 182.0 | 78.0 | 282.0 | 24.7 | 0.2 | −0.1 | 0.36 ** |
| Total number of vascular bundles (No.) | 275.0 | 120.0 | 570.0 | 37.7 | 1.0 | 0.7 | 386.0 | 167.0 | 672.0 | 27.4 | 0.5 | 0.0 | 0.55 ** |
| Area of large vascular bundles (mm2) | 0.015 | 0.006 | 0.038 | 39.4 | 1.7 | 3.4 | 0.023 | 0.013 | 0.043 | 25.9 | 1.2 | 2.0 | 0.48 ** |
| Area of small vascular bundles (mm2) | 0.006 | 0.002 | 0.021 | 47.4 | 2.5 | 10.2 | 0.009 | 0.004 | 0.020 | 47.4 | 1.2 | 1.1 | 0.57 ** |
| Stem diameter (mm) | 6.7 | 4.1 | 9.3 | 20.8 | 0.0 | −1.0 | 9.6 | 6.3 | 14.6 | 18.6 | 0.6 | 0.6 | 0.47 ** |
| Stem area (mm2) | 33.7 | 11.9 | 60.7 | 39.5 | 0.2 | −1.0 | 66.9 | 28.4 | 159.8 | 39.5 | 1.4 | 2.5 | 0.52 ** |
| Rind area (mm2) | 4.3 | 1.2 | 9.9 | 48.9 | 0.7 | 0.1 | 7.6 | 4.2 | 14.9 | 28.8 | 1.2 | 2.2 | 0.31 * |
| Percentage of TALVB to SA-CA (%) | 6.4 | 3.2 | 15.3 | 41.0 | 1.8 | 2.6 | 7.1 | 4.0 | 11.5 | 24.8 | 0.5 | −0.1 | 0.37 ** |
| First Season of 2021 | Second Season of 2021 | |||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Number of large vascular bundles | 11.6 | 0.2 | 7.4 | 10.5 | 1.4 | 1.5 |
| Number of small vascular bundles | 7.2 | 3.0 | 16.1 | 8.8 | 1.0 | 0.0 |
| Total number of vascular bundles | 11.4 | 0.9 | 11.7 | 11.2 | 1.4 | 0.6 |
| Area of large vascular bundles | 3.8 | 19.1 | 12.5 | 8.4 | 1.8 | 13.5 |
| Area of small vascular bundles | 2.0 | 18.6 | 10.1 | 3.9 | 0.3 | 36.8 |
| Stem diameter | 12.0 | 2.9 | 1.5 | 9.6 | 8.3 | 3.0 |
| Stem area | 12.2 | 2.3 | 1.4 | 10.4 | 4.5 | 3.2 |
| Rind area | 8.5 | 6.6 | 2.1 | 8.4 | 5.0 | 2.0 |
| Percentage of TALVB to SA-CA | 5.5 | 4.6 | 2.9 | 5.7 | 0.2 | 23.5 |
| Sixth internode length | 4.7 | 18.7 | 10.0 | 3.9 | 33.4 | 0.1 |
| Sixth internode diameter | 8.4 | 1.5 | 4.8 | 6.7 | 2.5 | 13.6 |
| Breaking force | 7.8 | 7.4 | 9.4 | 7.8 | 11.6 | 1.7 |
| Stalk strength | 5.0 | 14.2 | 10.0 | 4.8 | 28.6 | 0.3 |
| Lodging index | 51.5 | 17.2 | 11.8 | 60.4 | 13.6 | 9.0 |
| the variance contribution rate of the PC | 51.5 | 17.2 | 11.8 | 60.4 | 13.6 | 9.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, H.; Wang, J.; Shen, S.; Yang, M.; Li, S.; Feng, B.; Zhong, R.; Ma, C.; Wang, J.; Xie, R.; et al. High-Throughput Phenotyping of Cross-Sectional Morphology to Assess Stalk Mechanical Properties in Sorghum. Agriculture 2022, 12, 1696. https://doi.org/10.3390/agriculture12101696
Fan H, Wang J, Shen S, Yang M, Li S, Feng B, Zhong R, Ma C, Wang J, Xie R, et al. High-Throughput Phenotyping of Cross-Sectional Morphology to Assess Stalk Mechanical Properties in Sorghum. Agriculture. 2022; 12(10):1696. https://doi.org/10.3390/agriculture12101696
Chicago/Turabian StyleFan, Hongzeng, Jibin Wang, Songhao Shen, Mingchong Yang, Suli Li, Bihong Feng, Ruimin Zhong, Chongjian Ma, Jihong Wang, Ruohan Xie, and et al. 2022. "High-Throughput Phenotyping of Cross-Sectional Morphology to Assess Stalk Mechanical Properties in Sorghum" Agriculture 12, no. 10: 1696. https://doi.org/10.3390/agriculture12101696