

Ceruloplasmin Interferes with the Assessment of Blood Lipid Hydroperoxide Content in Small Ruminants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Blood Sampling
2.2. Analytical Methods
2.3. Statistical Analysis
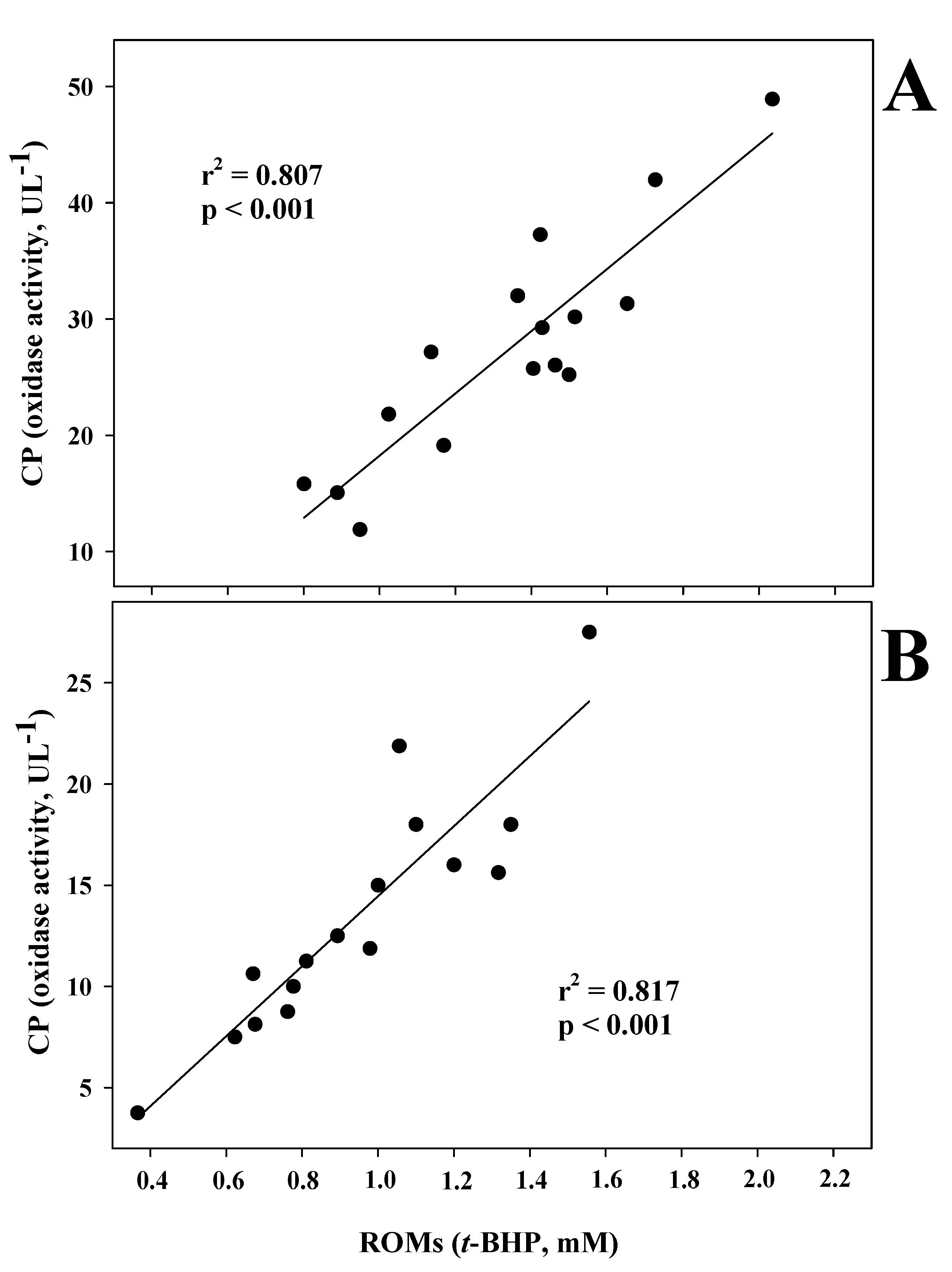
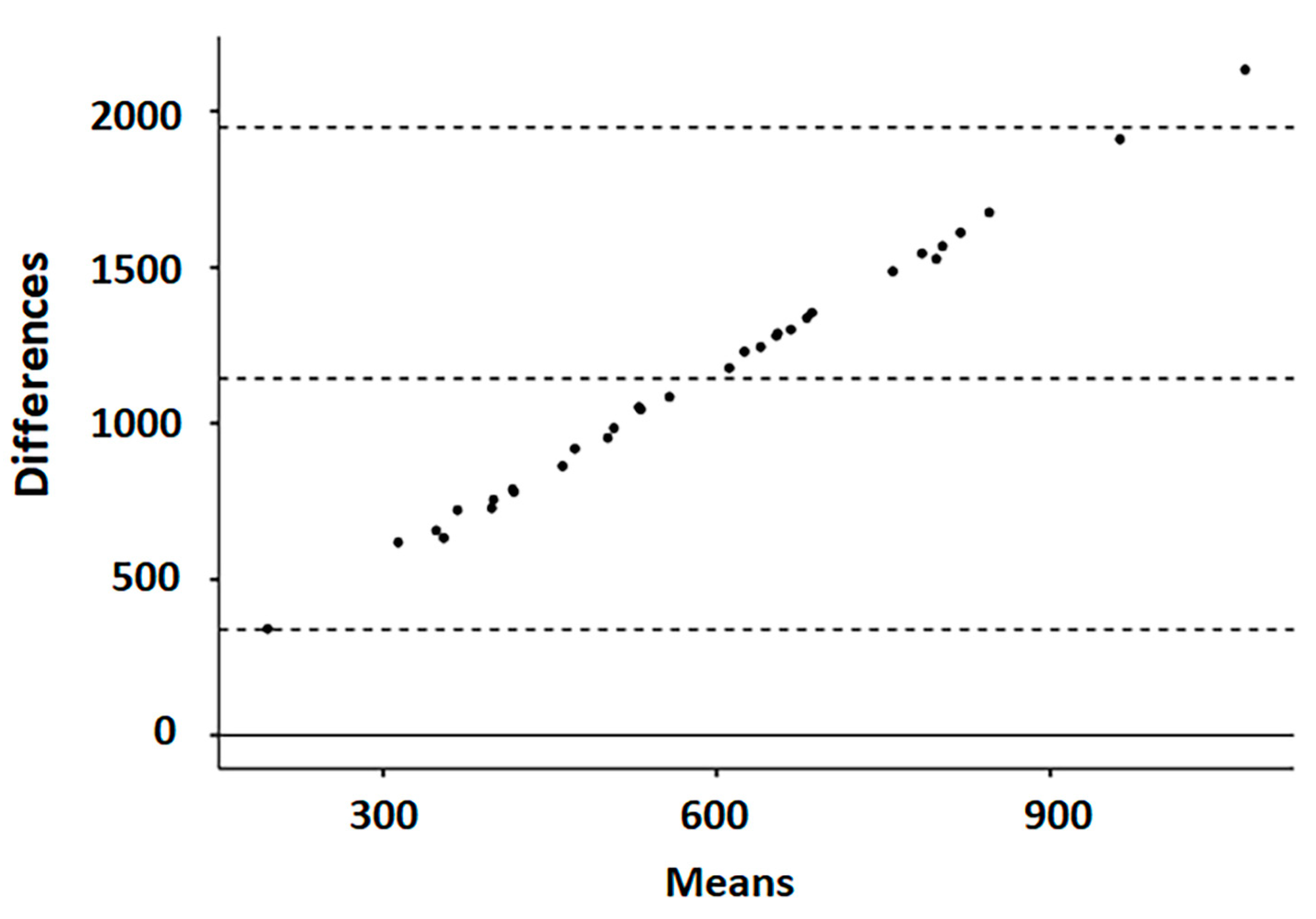
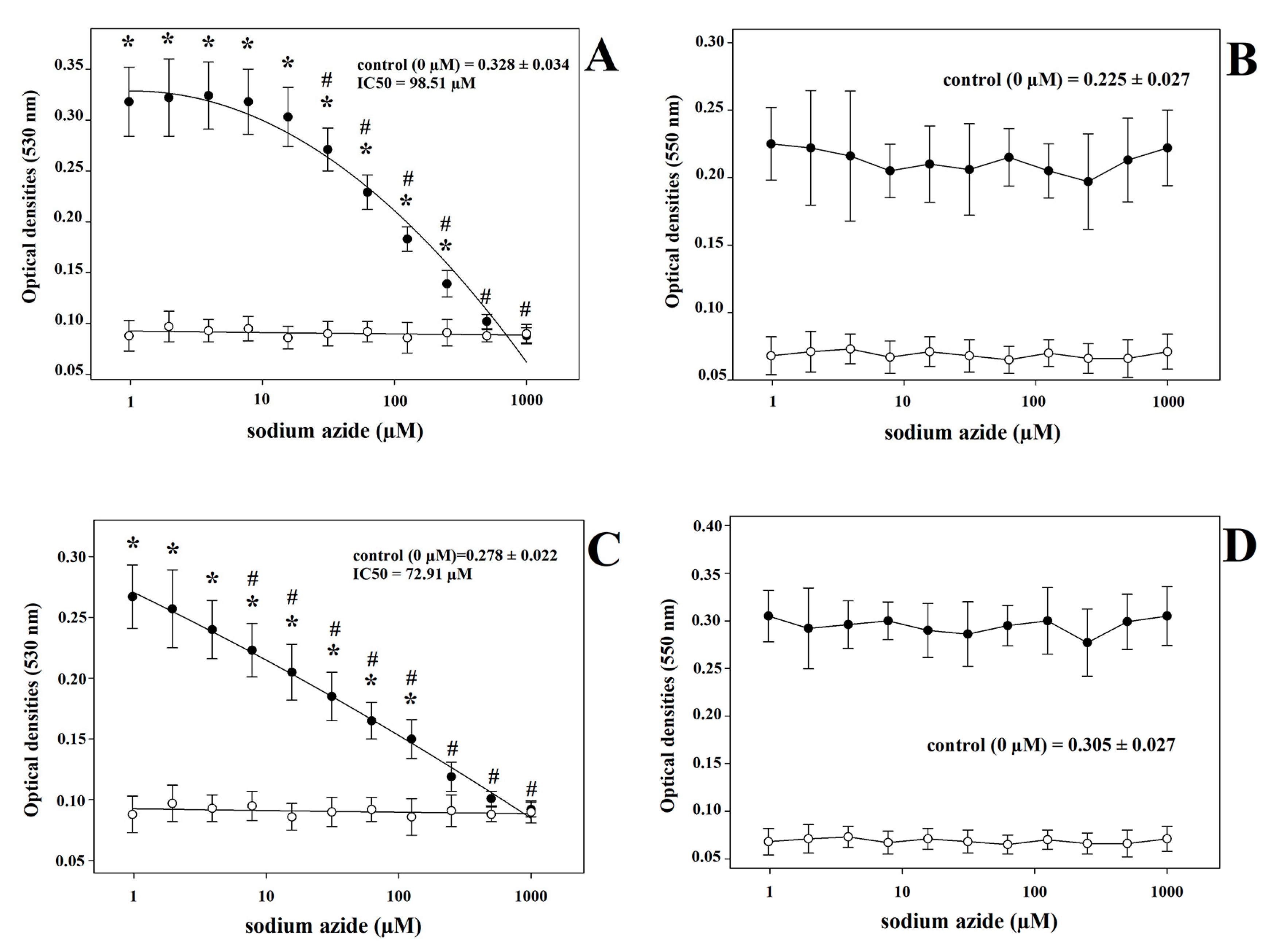
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, C.; Feng, C.; Yan, C.; Yu, Y.; Chen, Z.; Guo, C.; Wang, X. Role of mitochondrial reactive oxygen species in homeostasis regulation. Redox Rep. 2022, 27, 45–52. [Google Scholar] [CrossRef]
- Cadenas, E. Biochemistry of oxygen toxicity. Annu. Rev. Biochem. 1989, 58, 79–110. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef]
- Toyokuni, S. Reactive oxygen species-induced molecular damage and its application in pathology. Pathol. Int. 1999, 49, 91–102. [Google Scholar] [CrossRef]
- Tarpey, M.M.; Wink, D.A.; Grisham, M.B. Methods for detection of reactive metabolites of oxygen and nitrogen: In vitro and in vivo considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R431–R444. [Google Scholar] [CrossRef]
- Pitocco, D.; Zaccardi, F.; Di Stasio, E.; Romitelli, F.; Santini, S.A.; Zuppi, C.; Ghirlanda, G. Oxidative stress, nitric oxide, and diabetes. Rev. Diabet. Stud. 2010, 7, 15. [Google Scholar] [CrossRef]
- Gardner, H.W. Oxygen radical chemistry of polyunsaturated fatty acids. Free Radic. Biol. Med. 1989, 7, 65–86. [Google Scholar] [CrossRef]
- Giustarini, D.; Dalle-Donne, I.; Tsikas, D.; Rossi, R. Oxidative stress and human diseases: Origin, link, measurement, mechanisms, and biomarkers. Crit. Rev. Clin. Lab. Sci. 2009, 46, 241–281. [Google Scholar] [CrossRef]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2011, 33, 233–240. [Google Scholar] [CrossRef]
- Alberti, A.; Bolognini, L.; Macciantelli, D.; Caratelli, M. The radical cation of N, N-diethyl-para-phenylendiamine: A possible indicator of oxidative stress in biological samples. Res. Chem. Intermed. 2000, 26, 253–267. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Jansen, E.H.; Ruskovska, T. Comparative analysis of serum (anti) oxidative status parameters in healthy persons. Int. J. Mol. Sci. 2013, 14, 6106–6115. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, S.; Fazio, F. Assessment of total (anti) oxidant status in goat kids. Arch. Anim. Breed. 2021, 64, 139–146. [Google Scholar] [CrossRef]
- Cecchini Gualandi, S. Relation Between Redox Potential and Natural Antibody Levels in Goat Kid Serum. Vet. Immunol. Immunopathol. 2022, 254, 110517. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Clement, M.V.; Long, L.H. Hydrogen peroxide in the human body. FEBS Lett. 2000, 486, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Kilk, K.; Meitern, R.; Härmson, O.; Soomets, U.; Hõrak, P. Assessment of oxidative stress in serum by d-ROMs test. Free Radic. Res. 2014, 48, 883–889. [Google Scholar] [CrossRef]
- Osaki, S.; Johnson, D.A.; Frieden, E. The possible significance of the ferrous oxidase activity of ceruloplasmin in normal human serum. J. Biol. Chem. 1966, 241, 2746–2751. [Google Scholar] [CrossRef]
- Chapman, A.P.; Mocatta, T.; Shiva, S.; Seidel, A.; Chen, B.; Khalilova, I.; Paumann-Page, M.E.; Jameson, G.L.; Winterbourn, C.; Kettle, A.J. Ceruloplasmin is an endogenous inhibitor of myeloperoxidase. J. Biol. Chem. 2013, 288, 6465–6477. [Google Scholar] [CrossRef]
- Nazligul, Y.; Aslan, M.; Horoz, M.; Celik, Y.; Dulger, A.C.; Celik, H.; Erel, O. The effect on serum myeloperoxidase activity and oxidative status of eradication treatment in patients Helicobacter pylori infected. Clin. Biochem. 2011, 44, 647–649. [Google Scholar] [CrossRef]
- Soriano, V.S.; e Sá, J.; Junior, H.P.R.; Torbitz, V.D.; Moresco, R.N.; Stefani, L.M.; Da Silva, A.S. Postpartum nitric oxide, oxidants and antioxidants levels in ewes and their lambs. Small Rumin. Res. 2015, 123, 13–16. [Google Scholar] [CrossRef]
- Schosinsky, K.H.; Lehmann, H.P.; Beeler, M.F. Measurement of ceruloplasmin from its oxidase activity in serum by use of o-dianisidine dihydrochloride. Clin. Chem. 1974, 20, 1556–1563. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet 1986, 327, 307–310. [Google Scholar] [CrossRef]
- Cecchini, S.; Fazio, F.; Bazzano, M.; Caputo, A.R.; Giannetto, C.; Piccione, G. Redox status and oxidative stress during late pregnancy and postpartum period in mares. Livest. Sci. 2019, 230, 103821. [Google Scholar] [CrossRef]
- Cecchini, S.; Piccione, G.; Saoca, C.; Giangrosso, G.; Caputo, A.; Fazio, F. Redox status, lipid peroxidation and protein oxidation levels in small ruminants. J. Anim. Feed Sci. 2018, 27, 81–85. [Google Scholar] [CrossRef]
- Celi, P.; Di Trana, A.; Claps, S. Effects of plane of nutrition on oxidative stress in goats during the peripartum period. Vet. J. 2010, 184, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Oikonomidis, I.L.; Kiosis, E.A.; Brozos, C.N.; Kritsepi-Konstantinou, M.G. Reference intervals for serum reactive oxygen metabolites, biological antioxidant potential, and oxidative stress index in adult rams. Am. J. Vet. Res. 2017, 78, 274–278. [Google Scholar] [CrossRef]
- Cheng, J.; Ma, H.; Fan, C.; Zhang, Z.; Jia, Z.; Zhu, X.; Wang, L. Effects of different copper sources and levels on plasma superoxide dismutase, lipid peroxidation, and copper status of lambs. Biol. Trace Elem. Res. 2011, 144, 570–579. [Google Scholar] [CrossRef]
- El-khaiat, H.M.; Ghanem, M.M.; Abou-Zeina, H.A.; Abd El-Raof, Y.M.; El-Attar, H.M.; Abd El-Aziem, S.H. Assessment of the immune status and DNA damage in goats with experimentally-induced hypocuprosis. Res. Zool. 2013, 3, 23–30. [Google Scholar]
- Atakisi, O.; Oral, H.; Atakisi, E.; Merhan, O.; Pancarci, S.M.; Ozcan, A.; Marasli, S.; Polat, B.; Colak, A.; Kaya, S. Subclinical mastitis causes alterations in nitric oxide, total oxidant and antioxidant capacity in cow milk. Res. Vet. Sci. 2010, 89, 10–13. [Google Scholar] [CrossRef]
- Padhy, R.; Acharya, S.S.; Devi, N.; Swain, S.; Rattan, R. Association of nitrosative and oxidative stress in young type 2 diabetic patients. Int. J. Res. Med. Sci. 2015, 3, 2694–2697. [Google Scholar] [CrossRef]
- Sayır, F.; Şehitoğulları, A.; Demir, H.; Aslan, M.; Çobanoğlu, U.; Bilgin, C. Serum prolidase activity, total oxidant/antioxidant, and nitric oxide levels in patients with esophageal squamous cell carcinoma. Turk. J. Thorac. Cardiovasc. 2019, 27, 206. [Google Scholar] [CrossRef] [PubMed]
- Akcay, Y.D.; Sagin, F.G.; Aksu, K.; Keser, G.; Taylor, E.; Knight, I.; Winyard, P.G.; Sozmen, E.Y. A panel of oxidative stress assays does not provide supplementary diagnostic information in Behcet’s disease patients. J. Inflamm. 2012, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.K.; Sheybani, B.; Jortzik, E.; Niemann, B.; Wilhelm, J.; Boening, A.; Becker, K. Redox status of patients before cardiac surgery. Redox Rep. 2018, 23, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Colombini, F.; Carratelli, M.; Alberti, A. Oxidative stress, d-ROMs test, and ceruloplasmin. Free Radic. Res. 2016, 50, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.; Rodwell, V.; Bender, D.; Botham, K.M.; Weil, P.A.; Kennelly, P.J. Harper’s Illustrated Biochemistry, 28th ed.; Citeseer: New York, NY, USA, 2009. [Google Scholar]
- Mehdi, W.A.; Yusof, F.; Mehde, A.A.; Zainulabdeen, J.A.; Raus, R.A.; Abdulbari, A.S. Effects of acute lymphoblastic leukemia on ceruloplasmin oxidase, copper and several markers of oxidative damage, in children. Asian Pac. J. Cancer Prev. 2015, 16, 5205–5210. [Google Scholar] [CrossRef]
- Cerón, J.; Martínez-Subiela, S. An automated spectrophotometric method for measuring canine ceruloplasmin in serum. Vet. Res. 2004, 35, 671–679. [Google Scholar] [CrossRef]
- Martinez-Subiela, S.; Tecles, F.; Ceron, J. Comparison of two automated spectrophotometric methods for ceruloplasmin measurement in pigs. Res. Vet. Sci. 2007, 83, 12–19. [Google Scholar] [CrossRef]
- Otomaru, K.; Oishi, S.; Fujimura, Y.; Iwamoto, Y.; Nagai, K.; Ijiri, M. Effects of vitamin C supplementation on the blood oxidative stress and antibody titre against vaccination in calves. J. Vet. Res. 2021, 65, 73–78. [Google Scholar] [CrossRef]
- Contreras, M.; Peres Rubio, C.; de la Fuente, J.; Villar, M.; Merino, O.; Mosqueda, J.; Cerón, J.J. Changes in serum biomarkers of oxidative stress in cattle vaccinated with tick recombinant antigens: A pilot study. Vaccines 2020, 9, 5. [Google Scholar] [CrossRef]
- Rivera-Gomis, J.; Peres Rubio, C.; Martínez Conesa, C.; Otal Salaverri, J.; Cerón, J.J.; Tortosa, D.E.; Cubero Pablo, M.J. Effects of dietary supplementation of garlic and oregano essential oil on biomarkers of oxidative status, stress and inflammation in postweaning piglets. Animals 2020, 10, 2093. [Google Scholar] [CrossRef]
- Mund, M.D.; Riaz, M.; Mirza, M.A.; Rahman, Z.u.; Mahmood, T.; Ahmad, F.; Ammar, A. Effect of dietary tryptophan supplementation on growth performance, immune response and anti-oxidant status of broiler chickens from 7 to 21 days. Vet. Med. Sci. 2020, 6, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Sagkan-Ozturk, A.; Durgut, R.; Ozturk, O. Oxidant/antioxidant status in lambs and sheep with liver and lung cystic echinococcosis diagnosed by ultrasonography and necropsy. Vet. Parasitol. 2015, 208, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Merle, U.; Eisenbach, C.; Weiss, K.H.; Tuma, S.; Stremmel, W. Serum ceruloplasmin oxidase activity is a sensitive and highly specific diagnostic marker for Wilson’s disease. J. Hepatol. 2009, 51, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Rahman, Z.U.; Muhammad, F.; Khaliq, T.; Anwar, H.; Awais, M.M.; Sadaf, S. Oxytocin induced oxidative stress in lactating Bubalis bubalis (Nili Ravi). BMC Vet. Res. 2013, 9, 1–6. [Google Scholar] [CrossRef]
- Stowe, H.D.; Lawler, D.F.; Kealy, R.D. Antioxidant status of pair-fed labrador retrievers is affected by diet restriction and aging. J. Nutr. 2006, 136, 1844–1848. [Google Scholar] [CrossRef]
- Du, Z.; Hemken, R.; Harmon, R. Copper metabolism of Holstein and Jersey cows and heifers fed diets high in cupric sulfate or copper proteinate. J. Dairy Sci. 1996, 79, 1873–1880. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Goat Kids | Lambs | Significance | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | p< | |
| ROMs | 1.342 ± 0.330 | 0.946 ± 0.311 | 0.01 |
| TOS | 13.64 ± 7.68 | 19.72 ± 9.53 | 0.05 |
| CP | 27.42 ± 9.85 | 13.52 ± 5.95 | 0.001 |
| Goats | Sheep | Goat Kids | Lambs | |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |
| ROMs | 0.286 ± 0.049 | 0.481 ± 0.139 | 0.757 ± 0.170 | 0.601 ± 0.086 |
| TOS | 13.46 ± 6.14 | 29.92 ± 16.12 | 17.45 ± 6.12 | 38.69 ± 23.54 |
| CP | 5.38 ± 1.44 | 5.75 ± 2.88 | 10.75 ± 6.43 | 5.38 ± 3.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cecchini Gualandi, S.; Boni, R. Ceruloplasmin Interferes with the Assessment of Blood Lipid Hydroperoxide Content in Small Ruminants. Antioxidants 2023, 12, 701. https://doi.org/10.3390/antiox12030701
Cecchini Gualandi S, Boni R. Ceruloplasmin Interferes with the Assessment of Blood Lipid Hydroperoxide Content in Small Ruminants. Antioxidants. 2023; 12(3):701. https://doi.org/10.3390/antiox12030701
Chicago/Turabian StyleCecchini Gualandi, Stefano, and Raffaele Boni. 2023. "Ceruloplasmin Interferes with the Assessment of Blood Lipid Hydroperoxide Content in Small Ruminants" Antioxidants 12, no. 3: 701. https://doi.org/10.3390/antiox12030701