
Different Effects and Mechanisms of Selenium Compounds in Improving Pathology in Alzheimer’s Disease
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
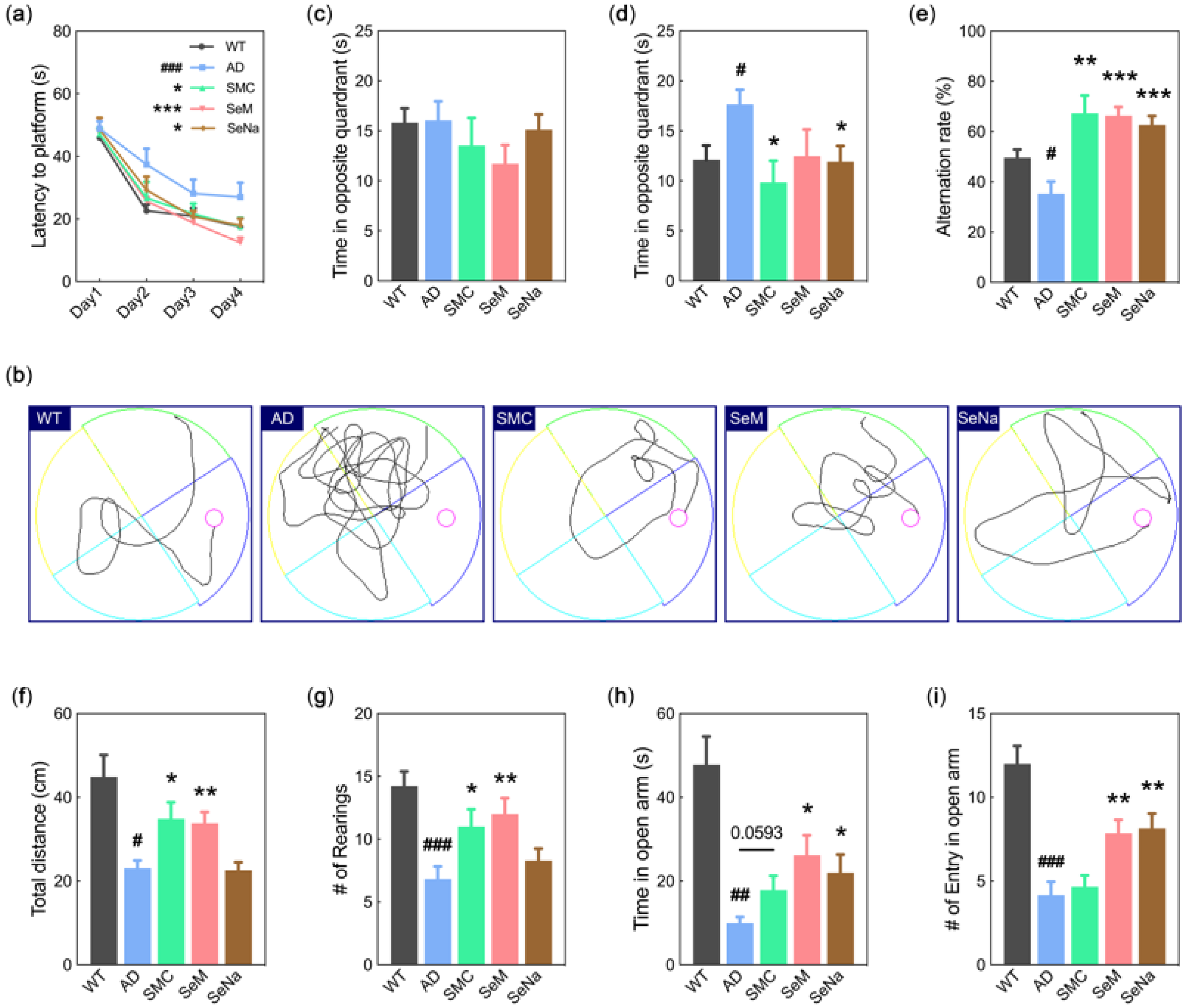
2.2. Morris Water Maze
2.3. Y-Maze Test
2.4. Open-Field Test
2.5. Elevated plus Maze
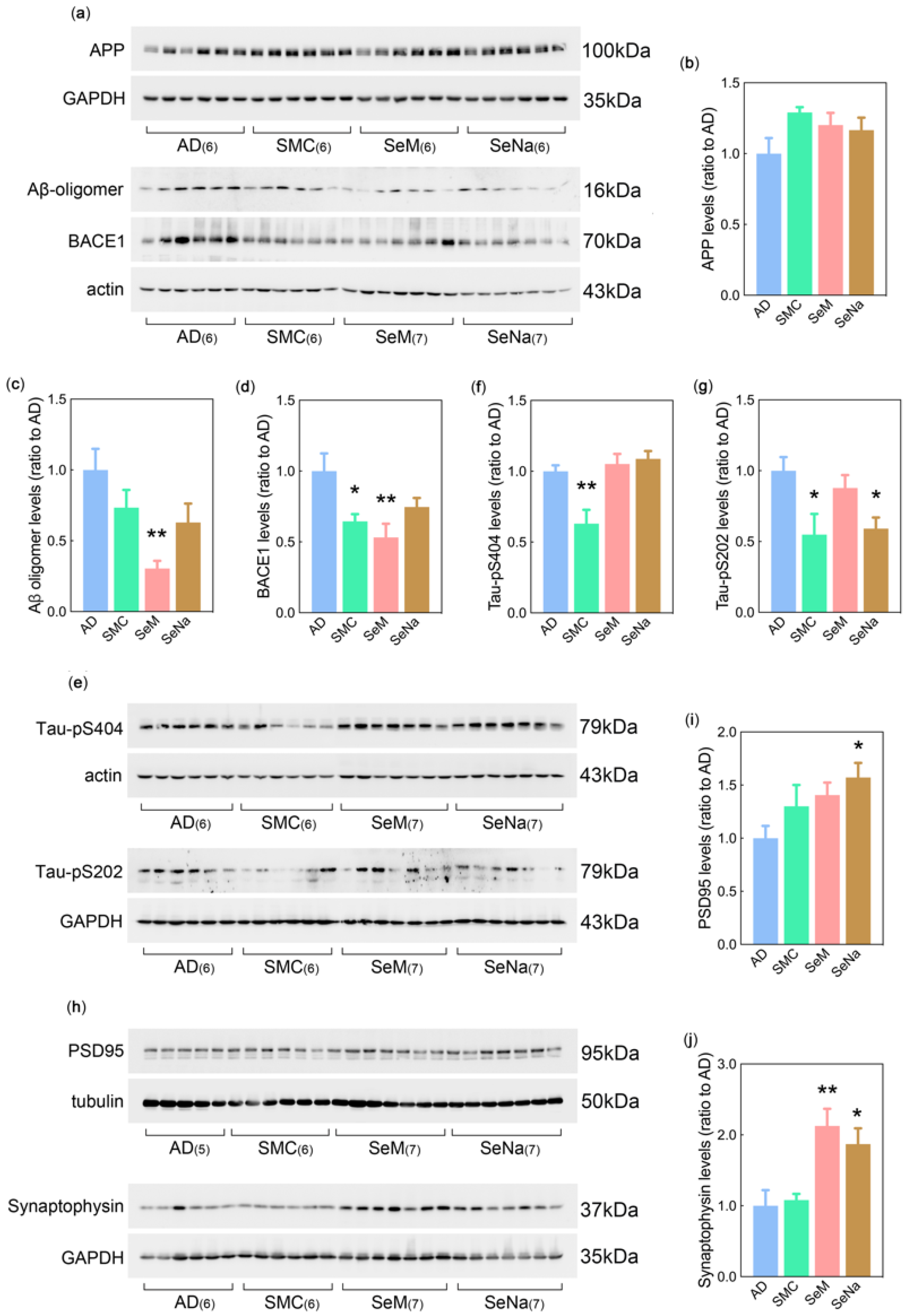
2.6. Western Blot Analysis
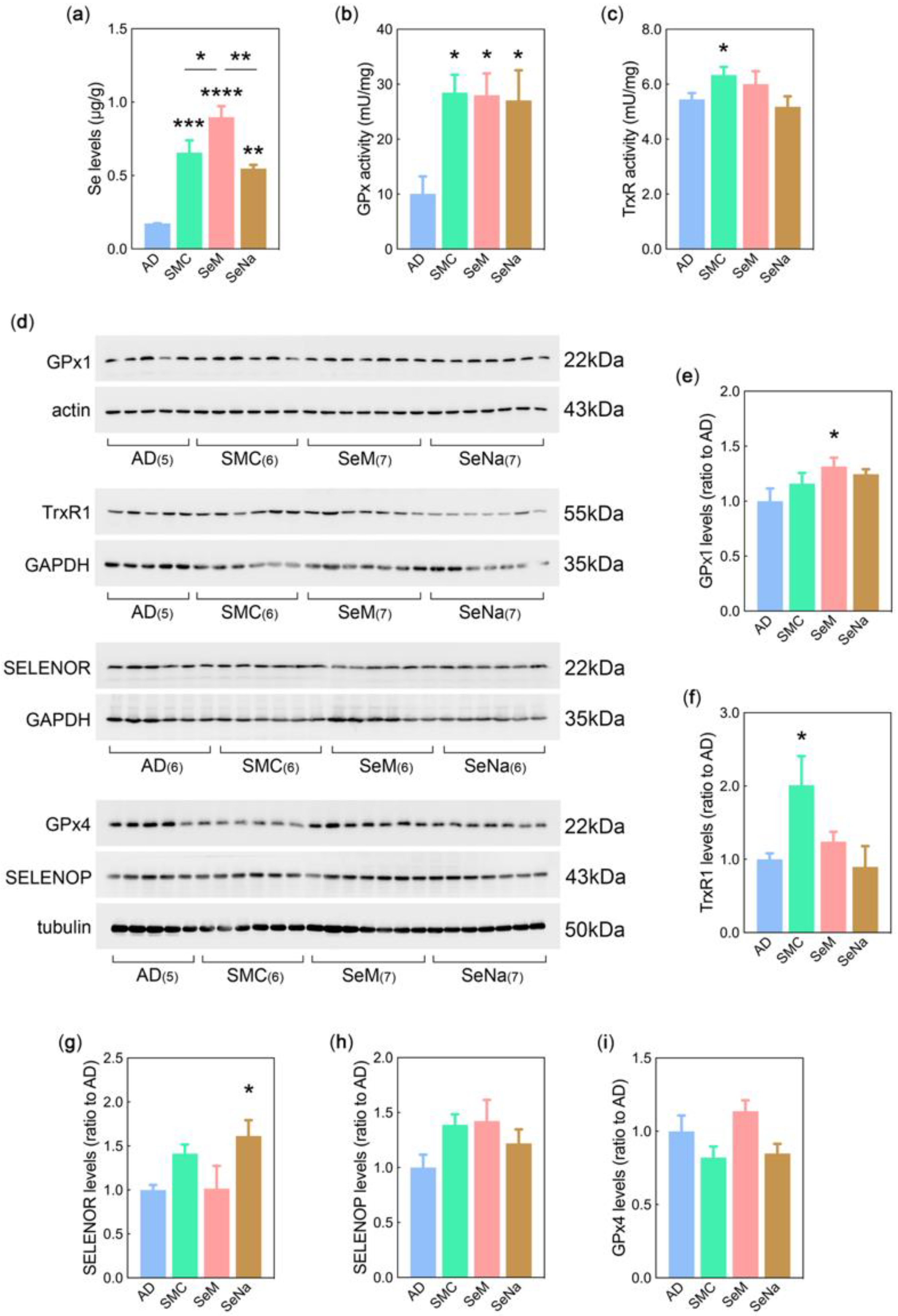
2.7. Se Levels and ELISA
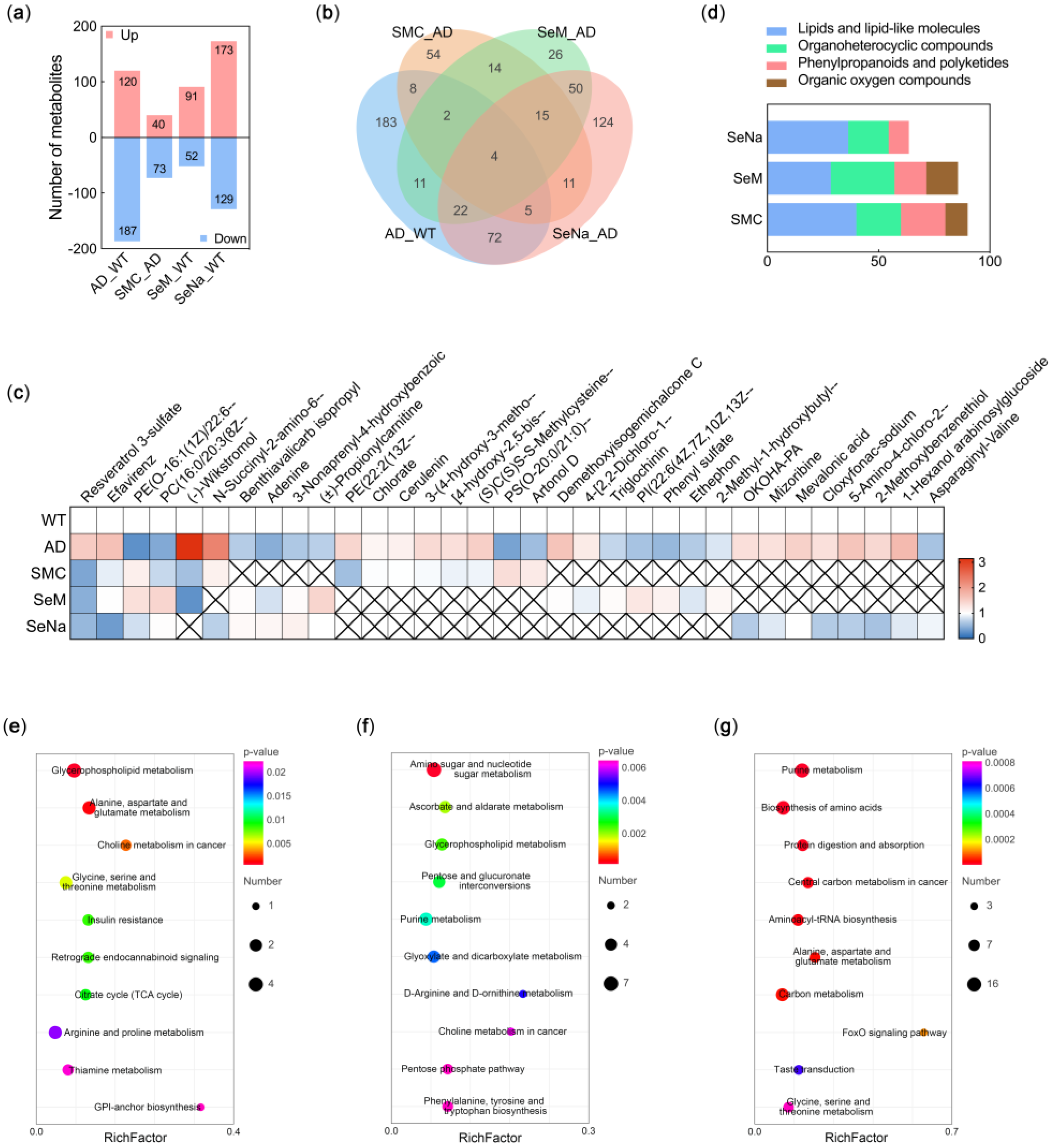
2.8. LC-MS Metabolomics
2.9. Statistical Analysis
3. Results
3.1. Cognitive Function
3.2. AD Pathology
3.3. Se Levels, Selenoenzyme Activity, and Selenoprotein Expression
3.4. Metabolic Profile in the Brain
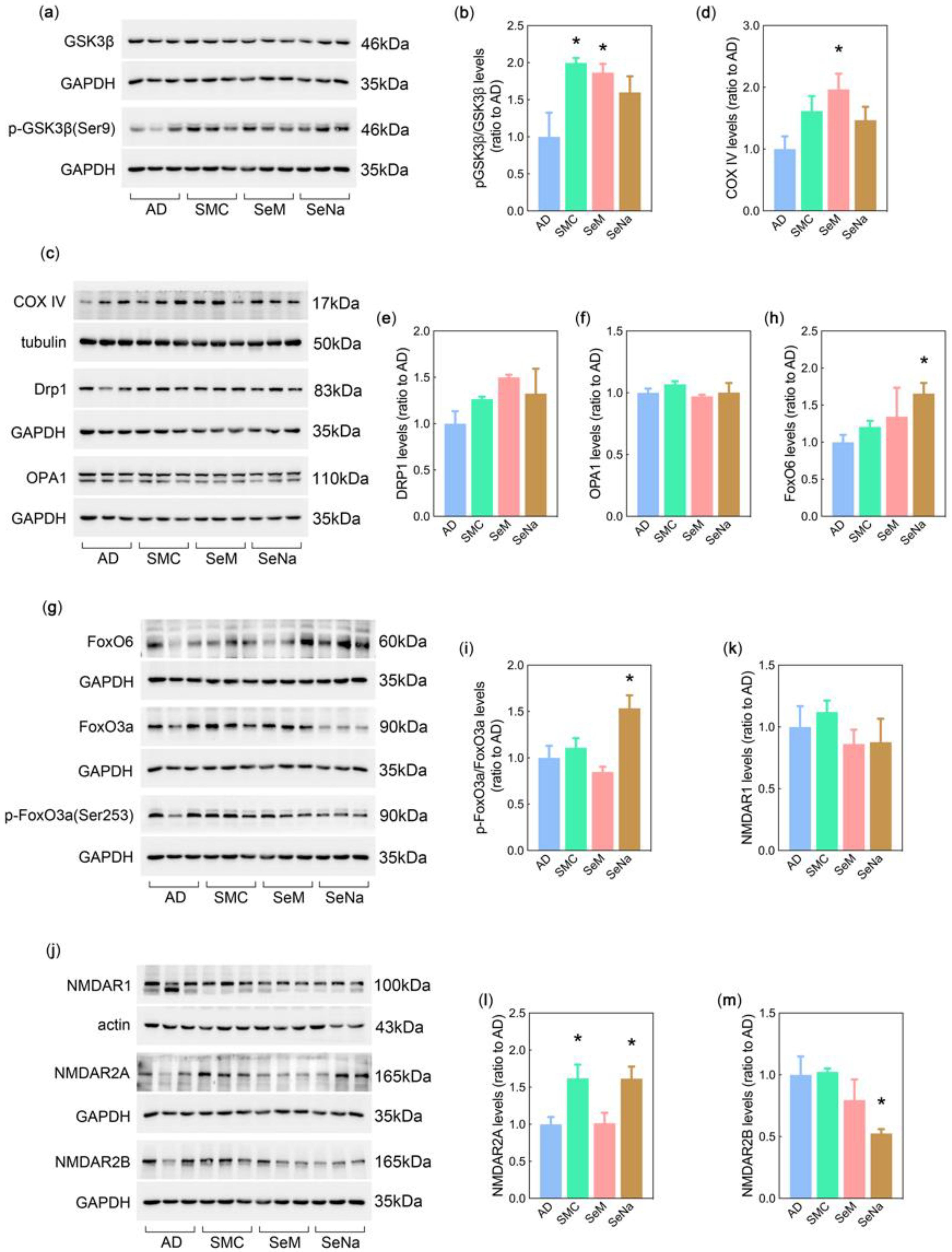
3.5. Mechanisms of Se Compounds in Improving AD Pathology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, T.; Zhang, D.; Zeng, Y.; Huang, T.Y.; Xu, H.; Zhao, Y. Molecular and cellular mechanisms underlying the pathogenesis of Alzheimer’s disease. Mol. Neurodegener. 2020, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, N.D. Importance of selenium and selenoprotein for brain function: From antioxidant protection to neuronal signalling. J. Inorg. Biochem. 2015, 153, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Sun, W.; Zhang, D. Association of Zinc, Iron, Copper, and Selenium Intakes with Low Cognitive Performance in Older Adults: A Cross-Sectional Study from National Health and Nutrition Examination Survey (NHANES). J. Alzheimers Dis. 2019, 72, 1145–1157. [Google Scholar] [CrossRef]
- Zhang, S.; Rocourt, C.; Cheng, W.H. Selenoproteins and the aging brain. Mech. Ageing Dev. 2010, 131, 253–260. [Google Scholar] [CrossRef]
- Pillai, R.; Uyehara-Lock, J.H.; Bellinger, F.P. Selenium and selenoprotein function in brain disorders. IUBMB Life 2014, 66, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Wang, C.; Liu, Q. Potential Roles of Selenium and Selenoproteins in the Prevention of Alzheimer’s Disease. Curr. Top Med. Chem. 2016, 16, 835–848. [Google Scholar] [CrossRef]
- Gwon, A.R.; Park, J.-S.; Park, J.-H.; Baik, S.-H.; Jeong, H.-Y.; Hyun, D.-H.; Park, K.W.; Jo, D.-G. Selenium attenuates A beta production and A beta-induced neuronal death. Neurosci. Lett. 2010, 469, 391–395. [Google Scholar] [CrossRef]
- Ishrat, T.; Parveen, K.; Khan, M.M.; Khuwaja, G.; Yousuf, S.; Ahmad, A.; Shrivastav, P.; Islam, F. Selenium prevents cognitive decline and oxidative damage in rat model of streptozotocin-induced experimental dementia of Alzheimer’s type. Brain Res. 2009, 1281, 117–127. [Google Scholar] [CrossRef]
- Van Eersel, J.; Ke, Y.; Liu, X.; Delerue, F.; Kril, J.; Götz, J.; Ittner, L. Sodium selenate mitigates tau pathology, neurodegeneration, and functional deficits in Alzheimer’s disease models. Proc. Natl. Acad. Sci. USA 2010, 107, 13888–13893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehdi, Y.; Hornick, J.-L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Zhang, Z.; Wen, L.; Chen, C.; Shi, Q.; Zhang, Y.; Ni, J.; Liu, Q. Selenomethionine ameliorates cognitive decline, reduces tau hyperphosphorylation, and reverses synaptic deficit in the triple transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2014, 41, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, Q.; Zheng, L.; Wang, B.; Qu, X.; Ni, J.; Zhang, Y.; Du, X. Se-Methylselenocysteine Ameliorates Neuropathology and Cognitive Deficits by Attenuating Oxidative Stress and Metal Dyshomeostasis in Alzheimer Model Mice. Mol. Nutr. Food Res. 2018, 62, e1800107. [Google Scholar] [CrossRef]
- Xie, Y.; Tan, Y.; Zheng, Y.; Du, X.; Liu, Q. Ebselen ameliorates beta-amyloid pathology, tau pathology, and cognitive impairment in triple-transgenic Alzheimer’s disease mice. J. Biol. Inorg. Chem. 2017, 22, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wu, Q.-Y.; Chen, C.; Zheng, R.; Chen, Y.; Liu, Q.; Ni, J.-Z.; Song, G.-L. Selenomethionine Attenuates the Amyloid-Beta Level by Both Inhibiting Amyloid-Beta Production and Modulating Autophagy in Neuron-2a/AbetaPPswe Cells. J. Alzheimers Dis. 2017, 59, 591–602. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Wu, Q.-Y.; Zheng, R.; Chen, C.; Chen, Y.; Liu, Q.; Hoffmann, P.R.; Ni, J.-Z.; Song, G.-L. Selenomethionine Mitigates Cognitive Decline by Targeting Both Tau Hyperphosphorylation and Autophagic Clearance in an Alzheimer’s Disease Mouse Model. J. Neurosci. 2017, 37, 2449–2462. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Shi, Q.; Zhao, Y.; Xie, Y.; Li, X.; Liu, Q.; Iqbal, J.; Zhang, H.; Liu, X.; Shen, L. Se-Methylselenocysteine (SMC) Improves Cognitive Deficits by Attenuating Synaptic and Metabolic Abnormalities in Alzheimer’s Mice Model: A Proteomic Study. ACS Chem. Neurosci. 2021, 12, 1112–1132. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Cao, X.-C.; Peng, J.-Y.; Huang, S.-L.; Chen, C.; Jia, S.-Z.; Ni, J.-Z.; Song, G.-L. Reversal of Lipid Metabolism Dysregulation by Selenium and Folic Acid Co-Supplementation to Mitigate Pathology in Alzheimer’s Disease. Antioxidants 2022, 11, 829. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, Y.; Shi, G.; Zhang, X. Selenium delays tomato fruit ripening by inhibiting ethylene biosynthesis and enhancing the antioxidant defense system. Food Chem. 2017, 219, 179–184. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Chen, C.; Wu, Q.-Y.; Zheng, R.; Liu, Q.; Ni, J.-Z.; Hoffmann, P.; Song, G.-L. Selenomethionine reduces the deposition of beta-amyloid plaques by modulating beta-secretase and enhancing selenoenzymatic activity in a mouse model of Alzheimer’s disease. Metallomics 2016, 8, 782–789. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Song, G.L. Roles of Selenoproteins in Brain Function and the Potential Mechanism of Selenium in Alzheimer’s Disease. Front. Neurosci. 2021, 15, 646518. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Deng, X.; Liu, N.; Li, M.; Liu, B.; Fu, Q.; Qu, R.; Ma, S. Quercetin attenuates tau hyperphosphorylation and improves cognitive disorder via suppression of ER stress in a manner dependent on AMPK pathway. J. Funct. Foods 2016, 22, 463–476. [Google Scholar] [CrossRef]
- Singh, P.K.; Kawasaki, M.; Berk-Rauch, H.; Nishida, G.; Yamasaki, T.; Foley, M.; Norris, E.; Strickland, S.; Aso, K.; Ahn, H.J. Aminopyrimidine Class Aggregation Inhibitor Effectively Blocks Abeta-Fibrinogen Interaction and Abeta-Induced Contact System Activation. Biochemistry 2018, 57, 1399–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Liu, Y.; Tian, D.; Tian, L.; Ju, X.; Qi, L.; Wang, Y.; Liang, C. Overview of cannabidiol (CBD) and its analogues: Structures, biological activities, and neuroprotective mechanisms in epilepsy and Alzheimer’s disease. Eur. J. Med. Chem. 2020, 192, 112163. [Google Scholar] [CrossRef] [PubMed]
- Fosso, M.Y.; Levine, H., 3rd; Green, K.D.; Tsodikov, O.V.; Garneau-Tsodikova, S. Effects of structural modifications on the metal binding, anti-amyloid activity, and cholinesterase inhibitory activity of chalcones. Org. Biomol. Chem. 2015, 13, 9418–9426. [Google Scholar] [CrossRef]
- Imperato, A.; Ramacci, M.T.; Angelucci, L. Acetyl-l-carnitine enhances acetylcholine release in the striatum and hippocampus of awake freely moving rats. Neurosci. Lett. 1989, 107, 251–255. [Google Scholar] [CrossRef]
- Ando, S.; Tadenuma, T.; Tanaka, Y.; Fukui, F.; Kobayashi, S.; Ohashi, Y.; Kawabata, T. Enhancement of learning capacity and cholinergic synaptic function by carnitine in aging rats. J. Neurosci. Res. 2001, 66, 266–271. [Google Scholar] [CrossRef]
- O’Malley, M. Clinical evaluation of pesticide exposure and poisonings. Lancet 1997, 349, 1161–1166. [Google Scholar] [CrossRef]
- Markova, N.; Bazhenova, N.; Anthony, D.C.; Vignisse, J.; Svistunov, A.; Lesch, K.-P.; Bettendorff, L.; Strekalova, T. Thiamine and benfotiamine improve cognition and ameliorate GSK-3β-associated stress-induced behaviours in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 75, 148–156. [Google Scholar] [CrossRef]
- Hanger, D.P.; Hughes, K.; Woodgett, J.R.; Brion, J.-P.; Anderton, B.H. Glycogen synthase kinase-3 induces Alzheimer’s disease-like phosphorylation of tau: Generation of paired helical filament epitopes and neuronal localisation of the kinase. Neurosci. Lett. 1992, 147, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Paudel, H.K.; Lew, J.; Ali, Z.; Wang, J.H. Brain proline-directed protein kinase phosphorylates tau on sites that are abnormally phosphorylated in tau associated with Alzheimer’s paired helical filaments. J. Biol. Chem. 1993, 268, 23512–23518. [Google Scholar] [CrossRef]
- Swerdlow, R.H. Mitochondria and Mitochondrial Cascades in Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1403–1416. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Song, J.; Wang, H.; Shi, F.; Zhou, N.; Jiang, J.; Xu, Y.; Zhang, L.; Yang, L.; Zhou, M. Chronic paradoxical sleep deprivation-induced depression-like behavior, energy metabolism and microbial changes in rats. Life Sci. 2019, 225, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Farhan, M.; Wang, H.; Gaur, U.; Little, P.; Xu, J.; Zheng, W. FOXO Signaling Pathways as Therapeutic Targets in Cancer. Int. J. Biol. Sci. 2017, 13, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Dong, H.H. FoxO integration of insulin signaling with glucose and lipid metabolism. J Endocrinol 2017, 233, R67–R79. [Google Scholar] [CrossRef] [Green Version]
- Salih, D.A.; Rashid, A.J.; Colas, D.; de la Torre-Ubieta, L.; Zhu, R.P.; Morgan, A.A.; Santo, E.E.; Ucar, D.; Devarajan, K.; Cole, C.J.; et al. FoxO6 regulates memory consolidation and synaptic function. Genes Dev. 2012, 26, 2780–2801. [Google Scholar] [CrossRef] [Green Version]
- Soriano, F.X.; Papadia, S.; Hofmann, F.; Hardingham, N.R.; Bading, H.; Hardingham, G.E. Preconditioning doses of NMDA promote neuroprotection by enhancing neuronal excitability. J. Neurosci. 2006, 26, 4509–4518. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, B.R.; Ong, T.P.; Jacob-Filho, W.; Jaluul, O.; Freitas, M.I.D.; Cozzolino, S.M.F. Nutritional status of selenium in Alzheimer’s disease patients. Br. J. Nutr. 2010, 103, 803–806. [Google Scholar] [CrossRef] [Green Version]
- Jin, N.; Zhu, H.; Liang, X.; Huang, W.; Xie, Q.; Xiao, P.; Ni, J.; Liu, Q. Sodium selenate activated Wnt/β-catenin signaling and repressed amyloid-β formation in a triple transgenic mouse model of Alzheimer’s disease. Exp. Neurol. 2017, 297, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, N.M.; Martin, D.; Hutter-Paier, B.; Windisch, M.; Nguyen, T.; Nheu, L.; Sundstrom, L.; Costello, A.; Hovens, C. Sodium selenate specifically activates PP2A phosphatase, dephosphorylates tau and reverses memory deficits in an Alzheimer’s disease model. J. Clin. Neurosci. 2010, 17, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Chung, Y.H.; Sharma, N.; Nguyen, B.T.; Lee, S.H.; Kang, S.W.; Nah, S.-Y.; Wie, M.B.; Nabeshima, T.; Jeong, J.H.; et al. Glutathione Peroxidase-1 Knockout Facilitates Memory Impairment Induced by beta-Amyloid (1-42) in Mice via Inhibition of PKC betaII-Mediated ERK Signaling; Application with Glutathione Peroxidase-1 Gene-Encoded Adenovirus Vector. Neurochem. Res. 2020, 45, 2991–3002. [Google Scholar] [CrossRef]
- Shin, E.J.; Lee, S.H.; Sharma, N.; Nguyen, B.T.; Chung, Y.H.; Kang, S.W.; Nah, S.-Y.; Lee, Y.J.; Nabeshima, T.; Jeong, J.H.; et al. An adenoviral vector encoded with the GPx-1 gene attenuates memory impairments induced by beta-amyloid (1-42) in GPx-1 KO mice via activation of M1 mAChR-mediated signalling. Free Radic. Res. 2021, 55, 11–25. [Google Scholar] [CrossRef]
- Lacerda, D.; Ortiz, V.; Türck, P.; Campos-Carraro, C.; Zimmer, A.; Teixeira, R.; Bianchi, S.; de Castro, A.L.; Schenkel, P.C.; Belló-Klein, A.; et al. Stilbenoid pterostilbene complexed with cyclodextrin preserves left ventricular function after myocardial infarction in rats: Possible involvement of thiol proteins and modulation of phosphorylated GSK-3beta. Free Radic. Res. 2018, 52, 988–999. [Google Scholar] [CrossRef]
- Yu, C.; Pan, S.; Dong, M.; Niu, Y. Astragaloside IV attenuates lead acetate-induced inhibition of neurite outgrowth through activation of Akt-dependent Nrf2 pathway in vitro. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Yang, Y.; Zhang, Z.; Zhang, L.; Song, J.; Ping, Y.; Du, X.; Song, G.; Liu, Q.; Li, N. Loss of MsrB1 perturbs spatial learning and long-term potentiation/long-term depression in mice. Neurobiol. Learn Mem. 2019, 166, 107104. [Google Scholar] [CrossRef]
- Papouin, T.; Ladépêche, L.; Ruel, J.; Sacchi, S.; Labasque, M.; Hanini, M.; Groc, L.; Pollegioni, L.; Mothet, J.-P.; Oliet, S.H. Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell 2012, 150, 633–646. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent | Source | Identifier |
|---|---|---|
| Rabbit anti-BACE1 | CST | 5606 |
| Mouse anti-Aβ | Biolegend | SIG-39300 |
| Rabbit anti-APP | Abcam | ab32136 |
| Mouse anti-Tau | Invitrogen | MN1000 |
| Mouse anti-pSTau (Ser202) | Invitrogen | MN1020 |
| Rabbit anti-pSTau (Ser202) | Abcam | Ab92676 |
| Rabbit anti-PSD95 | Abcam | ab76115 |
| Rabbit anti-Synaptophysin | CST | 4329s |
| Mouse anti-TrxR1 | Santa Cruz | sc-28321 |
| Rabbit anti-GPX4 | Abcam | ab125066 |
| Rabbit anti-GPX1 | GeneTex | GTX116040 |
| Rabbit anti-SELENOP | Abcam | ab109514 |
| Rabbit anti-MSRB1 (SELENOR) | Invitrogen | Cat# PA5-77009 |
| Mouse anti-GSK3β | CST | 9832 |
| Rabbit anti-pGSK3β (Ser9) | CST | 9323 |
| Rabbit anti-Foxo3a | CST | 2497 |
| Rabbit anti-pFoxo3a (Ser253) | CST | 9466 |
| Rabbit anti-Foxo6 | Proteintech | 19122 |
| Rabbit anti-β-actin | Proteintech | 81115 |
| Mouse anti-α-tubulin | Proteintech | 11224 |
| Rabbit anti-GAPDH | Proteintech | 10494 |
| Goat anti-Rabbit IgG | CST | 7074 |
| Goat anti-Mouse IgG | CST | 7076 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-H.; Peng, J.-Y.; Chen, Y.-B.; Wang, C.; Chen, C.; Song, G.-L. Different Effects and Mechanisms of Selenium Compounds in Improving Pathology in Alzheimer’s Disease. Antioxidants 2023, 12, 702. https://doi.org/10.3390/antiox12030702
Zhang Z-H, Peng J-Y, Chen Y-B, Wang C, Chen C, Song G-L. Different Effects and Mechanisms of Selenium Compounds in Improving Pathology in Alzheimer’s Disease. Antioxidants. 2023; 12(3):702. https://doi.org/10.3390/antiox12030702
Chicago/Turabian StyleZhang, Zhong-Hao, Jia-Ying Peng, Yu-Bin Chen, Chao Wang, Chen Chen, and Guo-Li Song. 2023. "Different Effects and Mechanisms of Selenium Compounds in Improving Pathology in Alzheimer’s Disease" Antioxidants 12, no. 3: 702. https://doi.org/10.3390/antiox12030702