
Transcriptional Inhibition of AGPAT2 Induces Abnormal Lipid Metabolism and Oxidative Stress in the Liver of Nile Tilapia Oreochromis niloticus
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of AGPAT2 Knock-Down Model
2.1.1. Experimental Fish
2.1.2. Design of Antisense RNA Sequences
- GCGGTATCATCCACAGCACATCCATAGCGAAACAGCCGCAGAAACTAGCAGCAGCACTGACTCACGCGGGGTGTTTCTGGATGAGGAAGCCGCTAACGGTTT.
- TAGCGCGGGCTAGCTGTCAGACGGCGCCGCGAGCGACATTTCTGCAAAACTCACCTCATGTTTTCAATGTCTCTGCCTCCGCTCTTTAGTATGCACAGAGGTATAGC.
2.1.3. PCR Amplification
2.1.4. Preparation of Transfection Reagent
2.1.5. Artificial Insemination and Incubation
2.1.6. Management of Experimental Fish
2.1.7. Detection of Positive Transfection Efficiency
2.2. Sampling
2.3. Measurement of Indexes
2.3.1. Calculation of Growth Indexes
2.3.2. Serum Index Determination
2.3.3. Determination of Liver Lipid Indexes
2.4. Determination of Liver Antioxidant Indexes
2.5. Histological Analyses
2.6. RNA Extraction and Reverse Transcription
2.7. Library Construction and Transcriptome Sequencing and Analysis
2.7.1. Library Construction
2.7.2. Assembly and Annotation of Transcripts
2.7.3. Identification of Differentially Expressed Genes
2.8. qRT-PCR Analyses
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
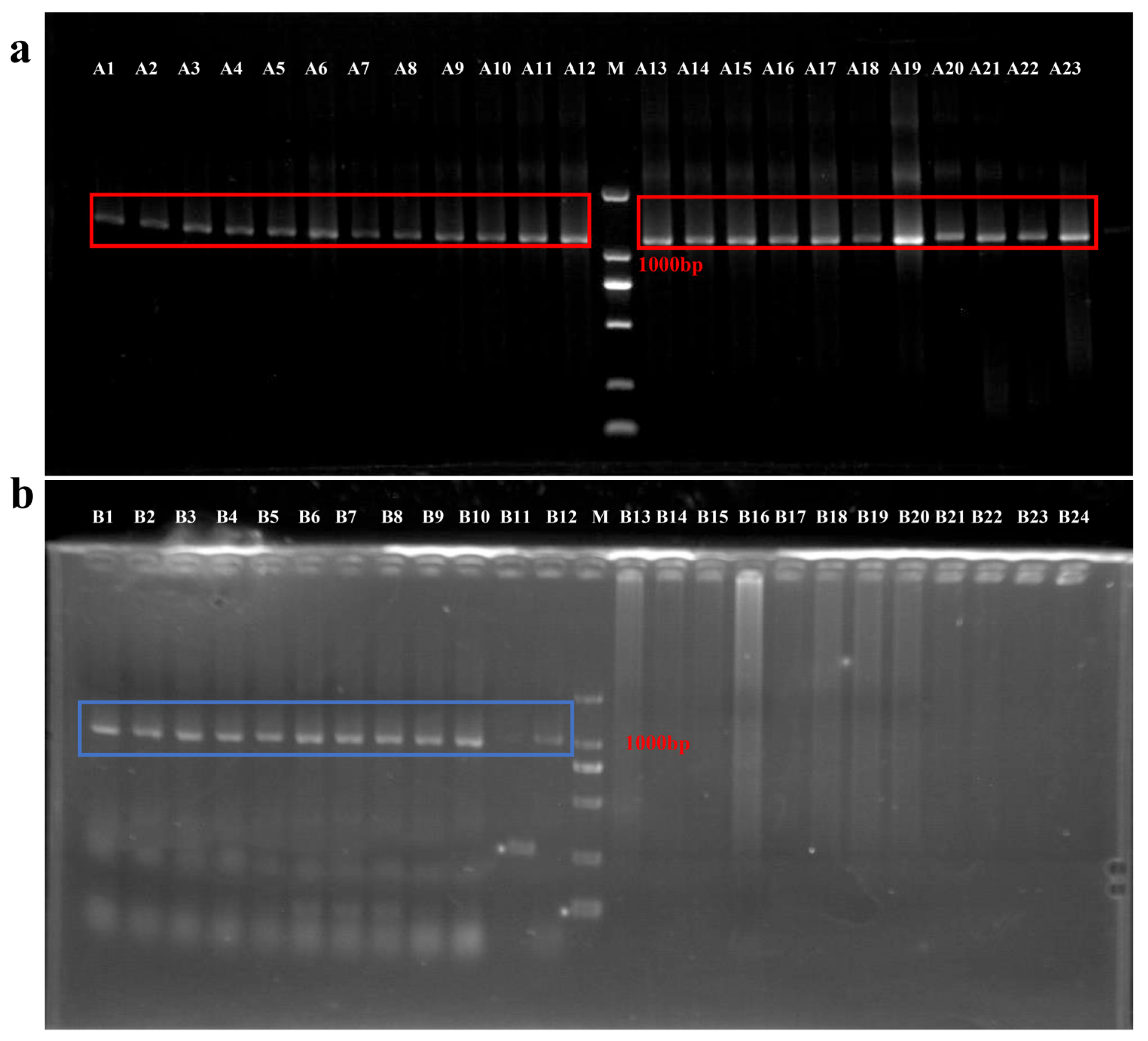
3.1. Positive Transfection Efficiency
3.2. Effects of AGPAT2 Knock-Down on Growth of Nile Tilapia
3.3. Effect of AGPAT2 Knock-Down on Serum Biochemical Parameters of Nile Tilapia
3.4. Effect of AGPAT2 Knock-Down on Liver Biochemical Indexes of Nile Tilapia
3.5. Effect of AGPAT2 Knock-Down on Liver Tissue Structure of Nile Tilapia
3.6. Antisense RNA Downregulated AGPAT2 Transcripts and AGPAT2 Protein in Nile Tilapia Liver
3.7. Transcriptomic Analysis to Determine the Effects of AGPAT2 Knock-Down on Liver Lipid Metabolism
3.7.1. Sequencing of mRNA Libraries and Identification of DEGs
3.7.2. Functional Annotations by GO and KEGG Analyses
3.7.3. qRT-PCR Validation of Gene Expression Patterns
4. Discussion
4.1. AGPAT2 Knock-Down Slows Growth of Nile Tilapia
4.2. AGPAT2 Knock-Down Causes Disordered Lipid Metabolism in Nile Tilapia
4.3. AGPAT2 Knock-Down Increases the Antioxidant Capacity of Nile Tilapia
4.4. AGPAT2 Knock-Down Affects Regulatory Mechanisms of Lipid Metabolism and Oxidative Stress in Nile Tilapia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malik, A.; Abbas, G.; Ghaffar, A.; Ferrando, S.; Gallus, L.; Shah, S.S.A. Effect of Different Salinity Level on Breeding, Fertilization, Hatching and Survival of Nile Tilapia, Oreochromis niloticus (Linnaeus, 1758) in Captivity. Pak. J. Zool. 2018, 50, 539–547. [Google Scholar] [CrossRef]
- Boujard, T.; Gelineau, A.; Coves, D.; Corraze, G.; Dutto, G.; Gasset, E.; Kaushik, S. Regulation of feed intake, growth, nutrient and energy utilisation in European sea bass (Dicentrarchus labrax) fed high fat diets. Aquaculture 2004, 231, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.J.; Cao, X.F.; Zhang, D.D.; Li, X.F.; Liu, W.B.; Jiang, G.Z. Chronic inflammation is a key to inducing liver injury in blunt snout bream (Megalobrama amblycephala) fed with high-fat diet. Dev. Comp. Immunol. 2019, 97, 28–37. [Google Scholar] [CrossRef]
- Agarwal, A.K. Lysophospholipid acyltransferases: 1-acylglycerol-3-phosphate O-acyltransferases. From discovery to disease. Curr. Opin. Lipidol. 2012, 23, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.M.; Duncan, R.E. The lysophosphatidic acid acyltransferases (acylglycerophosphate acyltransferases) family: One reaction, five enzymes, many roles. Curr. Opin. Lipidol. 2018, 29, 110–115. [Google Scholar] [CrossRef]
- Agarwal, A.K.; Arioglu, E.; De Almeida, S.; Akkoc, N.; Taylor, S.I.; Bowcock, A.M.; Barnes, R.I.; Garg, A. AGPAT2 is mutated in congenital generalized lipodystrophy linked to chromosome 9q34. Nat. Genet. 2002, 31, 21–23. [Google Scholar] [CrossRef]
- Haque, W.; Garg, A.; Agarwal, A.K. Enzymatic activity of naturally occurring 1-acylglycerol-3-phosphate-O-acyltransferase 2 mutants associated with congenital generalized lipodystrophy. Biochem. Biophys. Res. Commun. 2005, 327, 446–453. [Google Scholar] [CrossRef]
- Blunsom, N.J.; Cockcroft, S. CDP-Diacylglycerol Synthases (CDS): Gateway to Phosphatidylinositol and Cardiolipin Synthesis. Front. Cell Dev. Biol. 2020, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Gale, S.E.; Frolov, A.; Han, X.L.; Bickel, P.E.; Cao, L.; Bowcock, A.; Schaffer, J.E.; Ory, D.S. A regulatory role for 1-acylglycerol-3-phosphate-O-acyltransferase 2 in adipocyte differentiation. J. Biol. Chem. 2006, 281, 11082–11089. [Google Scholar] [CrossRef] [Green Version]
- Cortes, V.A.; Curtis, D.E.; Sukumaran, S.; Shao, X.L.; Parameswara, V.; Rashid, S.; Smith, A.R.; Ren, J.M.; Esser, V.; Hammer, R.E.; et al. Molecular Mechanisms of Hepatic Steatosis and Insulin Resistance in the AGPAT2-Deficient Mouse Model of Congenital Generalized Lipodystrophy. Cell Metab. 2009, 9, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Mak, H.Y.; Ouyang, Q.; Tumanov, S.; Xu, J.S.; Rong, P.; Dong, F.T.; Lam, S.M.; Wang, X.W.; Lukmantara, I.; Du, X.M.; et al. AGPAT2 interaction with CDP-diacylglycerol synthases promotes the flux of fatty acids through the CDP-diacylglycerol pathway. Nat. Commun. 2021, 12, 6877. [Google Scholar] [CrossRef]
- Bonham, L.; Leung, D.W.; White, T.; Hollenback, D.; Klein, P.; Tulinsky, J.; Coon, M.; de Vries, P.; Singer, J.W. Lysophosphatidic acid acyltransferase-β: A novel target for induction of tumour cell apoptosis. Expert Opin. Ther. Targets 2003, 7, 643–661. [Google Scholar] [CrossRef] [PubMed]
- Nobusue, H.; Kondo, D.; Yamamoto, M.; Kano, K. Effects of lysophosphatidic acid on the in vitro proliferation and differentiation of a novel porcine preadipocyte cell line. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 157, 401–407. [Google Scholar] [CrossRef]
- Lee, J.Y.; Cho, H.K.; Kwon, Y.H. Palmitate induces insulin resistance without significant intracellular triglyceride accumulation in HepG2 cells. Metab. Clin. Exp. 2010, 59, 927–934. [Google Scholar] [CrossRef]
- Cao, J.; Feng, X.X.; Yao, L.; Ning, B.; Yang, Z.X.; Fang, D.L.; Shen, W. Saturated Free Fatty Acid Sodium Palmitate-Induced Lipoapoptosis by Targeting Glycogen Synthase Kinase-3β Activation in Human Liver Cells. Dig. Dis. Sci. 2014, 59, 346–357. [Google Scholar] [CrossRef]
- Cao, Z.M.; Qiang, J.; Zhu, J.H.; Li, H.X.; Tao, Y.F.; He, J.; Xu, P.; Dong, Z.J. Transcriptional inhibition of steroidogenic factor 1 in vivo in Oreochromis niloticus increased weight and suppressed gonad development. Gene 2022, 809, 146203. [Google Scholar] [CrossRef]
- Boonanuntanasarn, S. Gene knockdown: A powerful tool for gene function study in fish. J. World Aquac. Soc. 2008, 39, 311–323. [Google Scholar] [CrossRef]
- Tomizawa, J.; Itoh, T.; Selzer, G.; Som, T. Inhibition of ColE1 RNA primer formation by a plasmid-specified small RNA. Proc. Natl. Acad. Sci. USA 1981, 78, 1421–1425. [Google Scholar] [CrossRef] [Green Version]
- Waki, M.; Kitanaka, A.; Kamano, H.; Tanaka, T.; Kubota, Y.; Ohnishi, H.; Takahara, J.; Irino, S. Antisense src expression inhibits U937 human leukemia cell proliferation in conjunction with reduction of c-myb expression. Biochem. Biophys. Res. Commun. 1994, 201, 1001–1007. [Google Scholar] [CrossRef]
- Tada, Y.; Nakase, M.; Adachi, T.; Nakamura, R.; Shimada, H.; Takahashi, M.; Fujimura, T.; Matsuda, T. Reduction of 14-16 kDa allergenic proteins in transgenic rice plants by antisense gene. FEBS Lett. 1996, 391, 341–345. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.H.; Gao, L.L.; Liang, H.C.; Du, G.H.; Gong, T.; Yang, J.L.; Zhu, P. Downregulation of lanosterol synthase gene expression by antisense RNA technology in Saccharomyces cerevisiae. Acta Pharm. Sin. 2015, 50, 118–122. [Google Scholar]
- Qiang, J.; Cao, Z.M.; Zhu, H.J.; Tao, Y.F.; He, J.; Xu, P. Knock-down of amh transcription by antisense RNA reduces FSH and increases follicular atresia in female. Oreochromis Niloticus Gene 2022, 842, 146792. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tao, Y.F.; Cao, Z.M.; Lu, S.Q.; Xu, P.; Qiang, J. The Effect of Knocked-Down Anti-Mullerian Hormone mRNA on Reproductive Characters of Male Nile Tilapia (Oreochromis niloticus) through Inhibition of the TGF-Beta Signaling Pathway. Fishes 2022, 7, 299. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Tao, Y.F.; Qiang, J.; Bao, J.W.; Chen, D.J.; Yin, G.J.; Xu, P.; Zhu, H.J. Changes in Physiological Parameters, Lipid Metabolism, and Expression of MicroRNAs in Genetically Improved Farmed Tilapia (Oreochromis niloticus) with Fatty Liver Induced by a High-Fat Diet. Front. Physiol. 2018, 9, 1521. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Qiang, J.; Song, C.Y.; Xu, P. Transcriptome profiling reveal Acanthopanax senticosus improves growth performance, immunity and antioxidant capacity by regulating lipid metabolism in GIFT (Oreochromis niloticus). Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 37, 100784. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Qiang, J.; Tao, Y.F.; Bao, J.W.; Chen, D.J.; Li, H.X.; He, J.; Xu, P. High Fat Diet-Induced miR-122 Regulates Lipid Metabolism and Fat Deposition in Genetically Improved Farmed Tilapia (GIFT, Oreochromis niloticus) Liver. Front. Physiol. 2018, 9, 1422. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.Y. Causes of fatty liver in farmed fish: A review and new perspectives. J. Fish. China 2014, 38, 1628–1638. [Google Scholar]
- Mantovani, A.; Gatti, D.; Zoppini, G.; Lippi, G.; Bonora, G.; Byrne, C.D.; Nobili, V.; Targher, G. Association Between Nonalcoholic Fatty Liver Disease and Reduced Bone Mineral Density in Children: A Meta-Analysis. Hepatology 2019, 70, 812–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Tang, Q. Research progress of the mechanism and therapy of non-alcoholic fatty liver disease. Chin. Bull. Life Sci. 2018, 30, 1165–11672. [Google Scholar]
- Suo, X.X.; Yan, X.B.; Tan, B.P.; Pan, S.M.; Li, T.; Liu, H.; Huang, W.B.; Zhang, S.; Yang, Y.Z.; Dong, X.H. Lipid metabolism disorders of hybrid grouper (♀Epinephelus fuscointestinestatus x male ♂E. lanceolatu) induced by high-lipid diet. Front. Mar. Sci. 2022, 9, 990193. [Google Scholar] [CrossRef]
- Chandalia, M.; Garg, A.; Vuitch, F.; Nizzi, F. Postmortem findings in congenital generalized lipodystrophy. J. Clin. Endocrinol. Metab. 1995, 80, 3077–3081. [Google Scholar]
- Eliasen, K.; Patursson, E.J.; Mcadam, B.J.; Pino, e.; Morro, B.; Betancor, M.; Baily, J.; Rey, S. Liver colour scoring index, carotenoids and lipid content assessment as a proxy for lumpfish (Cyclopterus lumpus L.) health and welfare condition. Sci. Rep. 2020, 10, 8927. [Google Scholar] [CrossRef]
- He, W.X.; Xu, Y.J.; Ren, X.H.; Xiang, D.; Lei, K.; Zhang, C.L.; Liu, D. Vitamin E Ameliorates Lipid Metabolism in Mice with Nonalcoholic Fatty Liver Disease via Nrf2/CES1 Signaling Pathway. Dig. Dis. Sci. 2019, 64, 3182–3191. [Google Scholar] [CrossRef]
- Lu, K.L.; Xu, W.N.; Li, X.F.; Liu, W.B.; Wang, L.N.; Zhang, C.N. Hepatic triacylglycerol secretion, lipid transport and tissue lipid uptake in blunt snout bream (Megalobrama amblycephala) fed high-fat diet. Aquaculture 2013, 408, 160–168. [Google Scholar] [CrossRef]
- Kamalam, B.S.; Medale, F.; Kaushik, S.; Polakof, S.; Skiba-Cassy, S.; Panserat, S. Regulation of metabolism by dietary carbohydrates in two lines of rainbow trout divergently selected for muscle fat content. J. Exp. Biol. 2012, 215, 2567–2578. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.Y.; Zhao, X.Y.; Niu, B.; Chang, K. Dietary Sodium Butyrate Supplementation Attenuates the Detrimental Effects of High-Fat Diets on Growth Performance, Liver Health, and Disease Resistance in Grass Carp. N. Am. J. Aquac. 2022, 84, 392–401. [Google Scholar] [CrossRef]
- Lu, K.L.; Wang, L.N.; Zhang, D.D.; Liu, W.B.; Xu, W.N. Berberine attenuates oxidative stress and hepatocytes apoptosis via protecting mitochondria in blunt snout bream Megalobrama amblycephala fed high-fat diets. Fish Physiol. Biochem. 2017, 43, 65–76. [Google Scholar] [CrossRef]
- Chaiyapechara, S.; Casten, M.T.; Hardy, R.W.; Dong, F.M. Fish performance, fillet characteristics, and health assessment index of rainbow trout (Oncorhynchus mykiss) fed diets containing adequate and high concentrations of lipid and vitamin E. Aquaculture 2003, 219, 715–738. [Google Scholar] [CrossRef]
- Jiang, W.D.; Wena, H.L.; Liu, Y.; Jiang, J.; Wu, P.; Zhao, J.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; et al. Enhanced muscle nutrient content and flesh quality, resulting from tryptophan, is associated with anti-oxidative damage referred to the Nrf2 and TOR signalling factors in young grass carp (Ctenopharyngodon idella): Avoid tryptophan deficiency or excess. Food Chem. 2016, 199, 210–219. [Google Scholar] [CrossRef]
- Jia, R.; Du, J.L.; Cao, L.P.; Li, Y.; Johnson, O.; Gu, Z.Y.; Jeney, G.; Xu, P.; Yin, G.J. Antioxidative, inflammatory and immune responses in hydrogen peroxide-induced liver injury of tilapia (GIFT, Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 894–905. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Wang, Z.M.; Pu, X.Z.; Zhou, S.Y. Oxidative stress and oxidative damage in chemical carcinogenesis. Toxicol. Appl. Pharmacol. 2011, 254, 86–99. [Google Scholar] [CrossRef]
- Huang, C.Y.; Fujimura, M.; Chang, Y.Y.; Chan, P.H. Overexpression of copper-zinc superoxide dismutase attenuates acute activation of activator protein-1 after transient focal cerebral ischemia in mice. Stroke 2001, 32, 741–747. [Google Scholar] [CrossRef]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of metabolism and as therapeutic targets in cardiovascular disease. Part II: PPAR-ss/delta and PPAR-gamma. Future Cardiol. 2017, 13, 276–296. [Google Scholar] [CrossRef]
- Qi, C.; Zhu, Y.J.; Reddy, J.K. Peroxisome proliferator-activated receptors, coactivators, and downstream targets. Cell Biochem. Biophys. 2000, 32, 187–204. [Google Scholar] [CrossRef]
- Tailleux, A.; Wouters, K.; Staels, B. Roles of PPARs in NAFLD: Potential therapeutic targets. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012, 1821, 809–818. [Google Scholar] [CrossRef]
- Yu, P.; Li, D.D.; Xiao, W.; Zhao, L.G. Advances in the structure-activity relationships of chlorogenic acid derivatives. Chin. J. Med. Chem. 2018, 28, 144–156+163. [Google Scholar]
- Yun, J.Y.; Jin, H.G.; Cao, Y.; Zhang, L.C.; Zhao, Y.M.; Jin, X.; Yu, Y.S. RNA-Seq Analysis Reveals a Positive Role of HTR2A in Adipogenesis in Yan Yellow Cattle. Int. J. Mol. Sci. 2018, 19, 1760. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.Z.; Mita, R.; Beaulieu, M.; Gao, Z.H.; Godbout, R. Fatty acid binding proteins in brain development and disease. Int. J. Dev. Biol. 2010, 54, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, M.; Owada, Y.; Yoshikawa, T. Role of Polyunsaturated Fatty Acids and Fatty Acid Binding Protein in the Pathogenesis of Schizophrenia. Curr. Pharm. Des. 2011, 17, 168–175. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, D.L.; Yu, D.H.; Lv, C.H.; Luo, H.Y.; Wang, Z.Y. Molecular cloning and expression analysis of scd1 gene from large yellow croaker Larimichthys crocea under cold stress. Gene 2015, 568, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Kim, Y.C.; Gray-Keller, M.P.; Attie, A.D.; Ntambi, J.M. The biosynthesis of hepatic cholesterol esters and triglycerides is impaired in mice with a disruption of the gene for stearoyl-CoA desaturase 1. J. Biol. Chem. 2000, 275, 30132–30138. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Zhou, Y.G.; Sun, D.J.; Gao, Q.F.; Dong, S.L. Influence of Low Temperature and Feeding on Expressions of Unsaturated Fatty Acid Biosynthesis Genes of Rainbow Trout (Oncorhynchus mykiss). Period. Ocean Univ. China 2019, 49, 63–71. [Google Scholar]
- Ayisi, C.L.; Yamei, C.; Zhao, J.L. Genes, transcription factors and enzymes involved in lipid metabolism in fin fish. Agri Gene 2018, 7, 7–14. [Google Scholar] [CrossRef]
- Torres, J.L.; Novo-Veleiro, I.; Manzanedo, L.; Alvela-Suarez, L.; Macias, R.; Laso, F.J.; Marcos, M. Role of microRNAs in alcohol-induced liver disorders and non-alcoholic fatty liver disease. World J. Gastroenterol. 2018, 24, 4104–4118. [Google Scholar] [CrossRef] [PubMed]
- Wepy, J.A.; Galligan, J.J.; Kingsley, P.J.; Xu, S.; Goodman, M.C.; Tallman, K.A.; Rouzer, C.A.; Marnett, L.J. Lysophospholipases cooperate to mediate lipid homeostasis and lysophospholipid signaling. J. Lipid Res. 2019, 60, 360–374. [Google Scholar] [CrossRef] [Green Version]
- Wendel, A.A.; Lewin, T.M.; Coleman, R.A. Glycerol-3-phosphate acyltransferases: Rate limiting enzymes of triacylglycerol biosynthesis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2009, 1791, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, H.Y.; Zhu, Q. Progress in the study on mammalian diacylgycerol acyltransgerase (DGAT) gene and its biological function. Hereditas 2007, 29, 1167–1172. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Q.; Cheng, S.W.; Huang, J.; Diao, G.; Han, J. Atgl gene deletion predisposes to proximal tubule damage by impairing the fatty acid metabolism. Biochem. Biophys. Res. Commun. 2017, 487, 160–166. [Google Scholar] [CrossRef]
- Taxiarchis, A.; Mahdessian, H.; Silveira, A. PNPLA2 influences secretion of triglyceride-rich lipoproteins by human hepatoma cells. J. Lipid Res. 2019, 60, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Tavasoli, M.; Lahire, S.; Reid, T.; Brodovsky, M.; McMaster, C.R. Genetic diseases of the Kennedy pathways for membrane synthesis. J. Biol. Chem. 2020, 295, 17877–17886. [Google Scholar] [CrossRef]
- Menniti, M.; Iuliano, R.; Foeller, M.; Sopjani, M.; Alesutan, I.; Mariggio, S.; Nofziger, C.; Perri, A.M.; Amato, R.; Blazer-Yost, B.; et al. 60 kDa Lysophospholipase, a New Sgk1 Molecular Partner Involved in the Regulation of EnaC. Cell. Physiol. Biochem. 2010, 26, 587–596. [Google Scholar] [CrossRef]
- Mayr, J.A. Lipid metabolism in mitochondrial membranes. J. Inherit. Metab. Dis. 2015, 38, 137–144. [Google Scholar] [CrossRef]
- Keenan, K.; Hoffman, M.; Dullen, K.; O’Brien, K.M. Molecular drivers of mitochondrial membrane proliferation in response to cold acclimation in threespine stickleback. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 203, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.P.S.; Liu, W.H.; Au, D.W.T.; Anderson, D.M.; Wu, R.S.S. Antioxidant responses and lipid peroxidation in gills and erythrocytes of fish (Rhabdosarga sarba) upon exposure to Chattonella marina and hydrogen peroxide: Implications on the cause of fish kills. J. Exp. Mar. Biol. Ecol. 2006, 336, 230–241. [Google Scholar] [CrossRef]
- Tapia, P.J.; Figueroa, A.M.; Eisner, V.; Gonzalez-Hodar, L.; Robledo, F.; Agarwal, A.K.; Garg, A.; Cortes, V. Absence of AGPAT2 impairs brown adipogenesis, increases IFN stimulated gene expression and alters mitochondrial morphology. Metab. Clin. Exp. 2020, 111, 154341. [Google Scholar] [CrossRef]
- Cheng, L.; Sebo, T.J.; Cheville, J.C.; Pisansky, T.M.; Slezak, J.; Bergstralh, E.J.; Pacelli, A.; Neumann, R.M.; Zincke, H.; Bostwick, D.G. p53 protein overexpression is associated with increased cell proliferation in patients with locally recurrent prostate carcinoma after radiation therapy. Cancer 1999, 85, 1293–1299. [Google Scholar] [CrossRef]
- Dulic, V.; Kaufmann, W.K.; Wilson, S.J.; Tlsty, T.D.; Lees, E.; Harper, J.W.; Elledge, S.J.; Reed, S.I. p53-dependent inhibition of cyclin-dependent kinase activities in human fibroblasts during radiation-induced G1 arrest. Cell 1994, 76, 1013–1023. [Google Scholar] [CrossRef]
- Kim, J.H.; Qu, A.; Reddy, J.K.; Gao, B.; Gonzalez, F.J. Hepatic Oxidative Stress Activates the Gadd45b Gene by Way of Degradation of the Transcriptional Repressor STAT3. Hepatology 2014, 59, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | GenBank Number |
|---|---|---|
| agpat2 | F:GCAGGCACAAGTACCCATCA | XM_005454514.4 |
| R:CAGCACGCATCAGGTCAAAG | ||
| scd | F:ACAAGCTCTCCGTGCTGGTCAT | XM_005471382.2 |
| R:GCAGAGTTGGGACGAAGTAGGC | ||
| elovl6 | F:ACAGTTCAACGAGGACGAAGC | XM_003443399.5 |
| R:AGCAAGGGTGAGTGACCACAG | ||
| fasn | F:CCAGAATCAGCCTGTGGAGTA | XM_003454056.5 |
| R:GTTTCAGCCTCAGACTCGTTG | ||
| chkb | F:TGTGGAAGATGAACCTCGCC | XM_003440553.5 |
| R:CAGGGGGAAGACGCCATAAA | ||
| pnpla2 | F:GCCTCATTCGACGAGCAGAT | XM_003440346.5 |
| R:TCCAGGGTGAGAGTGTAGGG | ||
| lypla2 | F:TCGTGAACTGGGACAGCAAAG | XM_003458537.4 |
| R:CCGTCAAAGTATCCGCCCAC | ||
| fabp7a | F:TGCAGAAGTGGGATGGCAAA | XM_003442929.5 |
| R:GGCTTTCTCATACGTGCGGA | ||
| aspg | F:AGAAAGACCGGAGCCCATTT | XM_005476978.4 |
| R:TGGATCACTCCGCTCACTTG | ||
| pisd | F:CTGCCCTTCCTATTGGTGACT | XM_013276568.3 |
| R:GACTCACGCCACCCTTGA | ||
| pparαb | F:AACCATCTCCTTCTGAGCGG | XM_019361775.2 |
| R:CACTGGACAGACACCAAAGC | ||
| gadd45β | F:CTGCTCAGAAACAAGGCTGC | XM_003442905.5 |
| R:CCCAGAGACTCGCAAGATGG | ||
| cdk2 | F: CCCCGGTGACTCGGAAATAG | XM_003441494.5 |
| R: GATAAATCCTGCCGAGCCCA | ||
| gpat4 | F: AGGCATCTGTGTCGCTAACC | XM_003452655.2 |
| R: GAATGACCCCCATCAAGCCA | ||
| β-actin | F:CCACACAGTGCCCATCTACGA | EU887951.1 |
| R:CCACGCTCTGTCAGGATCTTCA |
| Control Group (n = 12) | Negative Control Group (n = 12) | AGPAT2 Knock-Down Group (n = 12) | |
|---|---|---|---|
| Initial body weight (g) | 2.99 ± 0.03 | 3.01 ± 0.02 | 3.01 ± 0.04 |
| Final body weight (g) | 144.91 ± 4.86 b | 148.44 ± 3.56 b | 107.28 ± 12.54 a |
| WGR (%) | 47.42 ± 1.63 b | 48.33 ± 1.23 b | 38.55 ± 3.02 a |
| SGR (%) | 4.31 ± 0.12 b | 4.33 ± 0.09 b | 4.07 ± 0.08 a |
| VSI (%) | 9.52 ± 0.36 b | 9.25 ± 0.38 b | 12.41 ± 0.56 a |
| HSI (%) | 0.86 ± 0.04 b | 0.84 ± 0.04 b | 1.30 ± 0.02 a |
| Control Group | Negative Control Group | AGPAT2 Knock-Down Group | |
|---|---|---|---|
| TC (nmol/g) | 1.84 ± 0.07 b | 1.97 ± 0.04 b | 3.10 ± 0.21 a |
| TG (nmol/g) | 0.23 ± 0.01 b | 0.30 ± 0.01 b | 0.65 ± 0.11 a |
| Glu (nmol/g) | 4.58 ± 0.26 b | 4.44 ± 0.03 b | 6.40 ± 0.54 a |
| HDL-C (mmol/L) | 1.13 ± 0.04 | 1.13 ± 0.00 | 1.04 ± 0.03 |
| LDL-C (mmol/L) | 0.35 ± 0.01 b | 0.35 ± 0.00 b | 0.64 ± 0.02 a |
| INS (mmol/L) | 13.01 ± 0.48 b | 13.00 ± 0.05 b | 18.13 ± 0.86 a |
| Control Group | Negative Control Group | AGPAT2 Knock-Down Group | |
|---|---|---|---|
| TC (nmol/g liver) | 1.02 ± 0.06 b | 1.07 ± 0.06 b | 2.58 ± 0.12 a |
| TG (nmol/g liver) | 4.41 ± 0.23 b | 4.52 ± 0.39 b | 8.05 ± 0.29 a |
| SOD (U/mg prot) | 7.71 ± 0.38 b | 7.73 ± 0.40 b | 9.75 ± 0.41 a |
| CAT (U/mg prot) | 43.83 ± 1.25 b | 42.57 ± 1.11 b | 64.53 ± 2.50 a |
| MDA (mmol/mg prot) | 24.03 ± 1.28 b | 25.05 ± 1.79 b | 14.63 ± 1.07 a |
| GSH-PX (U/mg prot) | 156.99 ± 11.59 b | 159.36 ± 4.30 b | 338.26 ± 13.67 a |
| Gene Name | KEGG Pathway | Fold Change | p Value |
|---|---|---|---|
| scd | Biosynthesis of unsaturated fatty acids; PPAR signaling pathway | 0.03 | 0.00 |
| elovl5 | Biosynthesis of unsaturated fatty acids | 0.14 | 0.00 |
| elovl6 | Biosynthesis of unsaturated fatty acids | 0.17 | 0.00 |
| fads | Biosynthesis of unsaturated fatty acids; PPAR signaling pathway | 0.21 | 0.00 |
| fabp7a | PPAR signaling pathway | 0.01 | 0.00 |
| cyp7a1 | PPAR signaling pathway | 5.58 | 0.00 |
| angptl4 | PPAR signaling pathway | 0.00 | 0.00 |
| fabp3 | PPAR signaling pathway | 0.15 | 0.00 |
| pparαb | PPAR signaling pathway | 0.01 | 0.00 |
| acsl4a | PPAR signaling pathway | 0.27 | 0.01 |
| cpt2 | PPAR signaling pathway | 0.10 | 0.04 |
| abca5 | ABC transporters | 0.13 | 0.01 |
| abca1b | ABC transporters | 0.08 | 0.00 |
| acat2 | Terpenoid backbone biosynthesis; Two-component system | 3.55 | 0.00 |
| cyp51 | Steroid biosynthesis | 3.22 | 0.00 |
| aspg | Glycerophospholipid metabolism | 4.95 | 0.00 |
| pisd | Glycerophospholipid metabolism | 3.37 | 0.00 |
| chkb | Glycerolipid metabolism; Glycerophospholipid metabolism | 0.07 | 0.01 |
| gpat4 | Glycerolipid metabolism; Glycerophospholipid metabolism | 0.08 | 0.00 |
| lypla2 | Glycerophospholipid metabolism | 0.24 | 0.01 |
| pnpla2 | Glycerolipid metabolism | 5.10 | 0.02 |
| fasn | Fatty acid biosynthesis; Insulin signaling pathway | 0.04 | 0.01 |
| cdk2 | p53 signaling pathway | 0.22 | 0.00 |
| gadd45β | p53 signaling pathway | 28.78 | 0.03 |
| sod1 | Peroxisome; Amyotrophic lateral sclerosis (ALS) | 2.15 | 0.04 |
| wars | Aminoacyl-tRNA biosynthesis | 0.09 | 0.01 |
| igfbp3 | p53 signaling pathway | 24.97 | 0.00 |
| nansa | Amino sugar and nucleotide sugar metabolism | 0.17 | 0.02 |
| dmgdh | Glycine, serine and threonine metabolism | 2.17 | 0.01 |
| hk1 | Fructose and mannose metabolism; Amino sugar and nucleotide sugar metabolism; Starch and sucrose metabolism | 0.03 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, T.; Tao, Y.; Yan, Y.; Lu, S.; Li, Y.; Zhang, X.; Qiang, J. Transcriptional Inhibition of AGPAT2 Induces Abnormal Lipid Metabolism and Oxidative Stress in the Liver of Nile Tilapia Oreochromis niloticus. Antioxidants 2023, 12, 700. https://doi.org/10.3390/antiox12030700
Feng T, Tao Y, Yan Y, Lu S, Li Y, Zhang X, Qiang J. Transcriptional Inhibition of AGPAT2 Induces Abnormal Lipid Metabolism and Oxidative Stress in the Liver of Nile Tilapia Oreochromis niloticus. Antioxidants. 2023; 12(3):700. https://doi.org/10.3390/antiox12030700
Chicago/Turabian StyleFeng, Tiantian, Yifan Tao, Yue Yan, Siqi Lu, Yan Li, Xing Zhang, and Jun Qiang. 2023. "Transcriptional Inhibition of AGPAT2 Induces Abnormal Lipid Metabolism and Oxidative Stress in the Liver of Nile Tilapia Oreochromis niloticus" Antioxidants 12, no. 3: 700. https://doi.org/10.3390/antiox12030700