
Thioredoxin-2 Regulates SqrR-Mediated Polysulfide-Responsive Transcription via Reduction of a Polysulfide Link in SqrR


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Overexpression and Purification of SqrR and TrxC
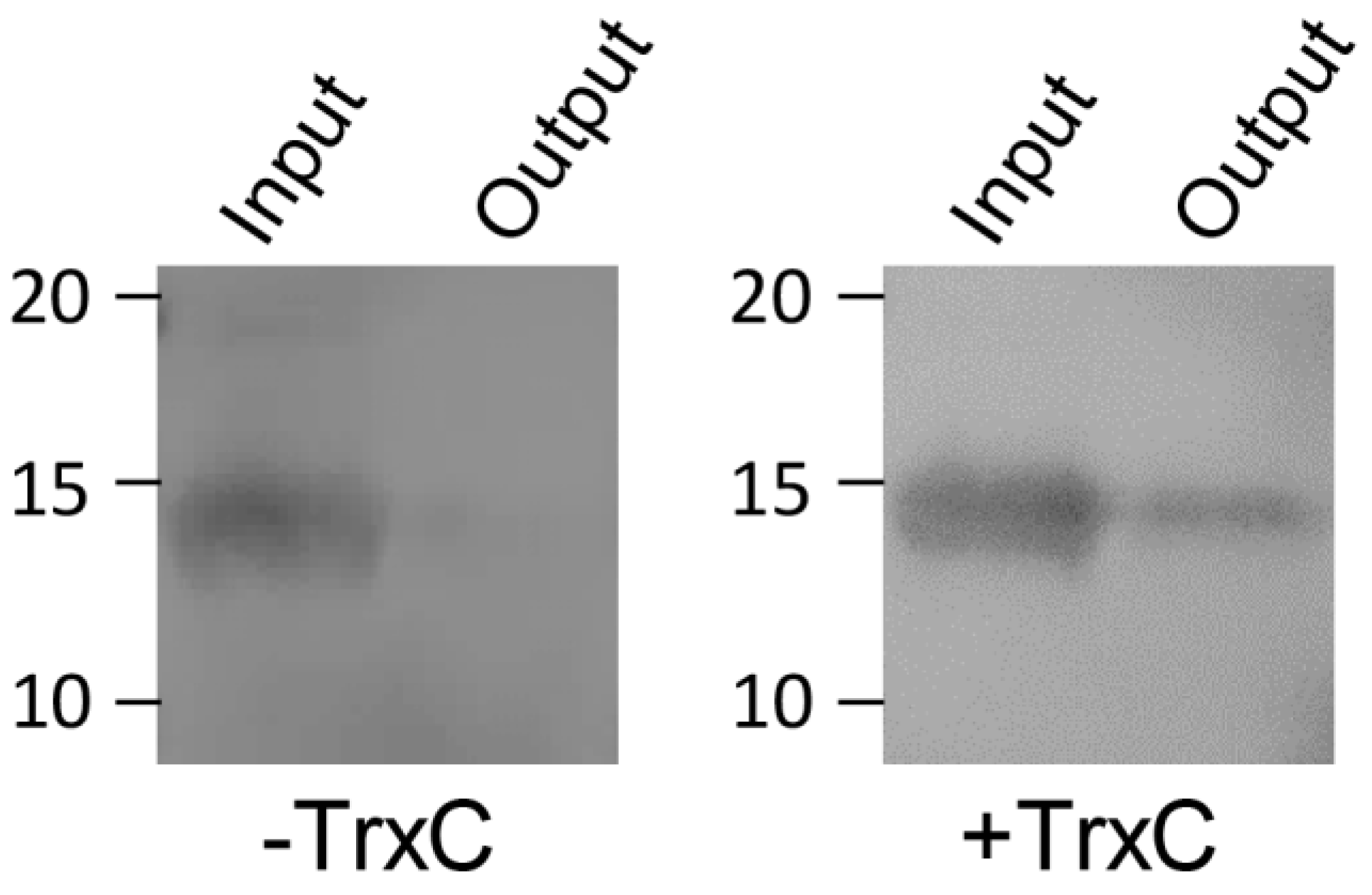
2.3. Pull-Down Assay
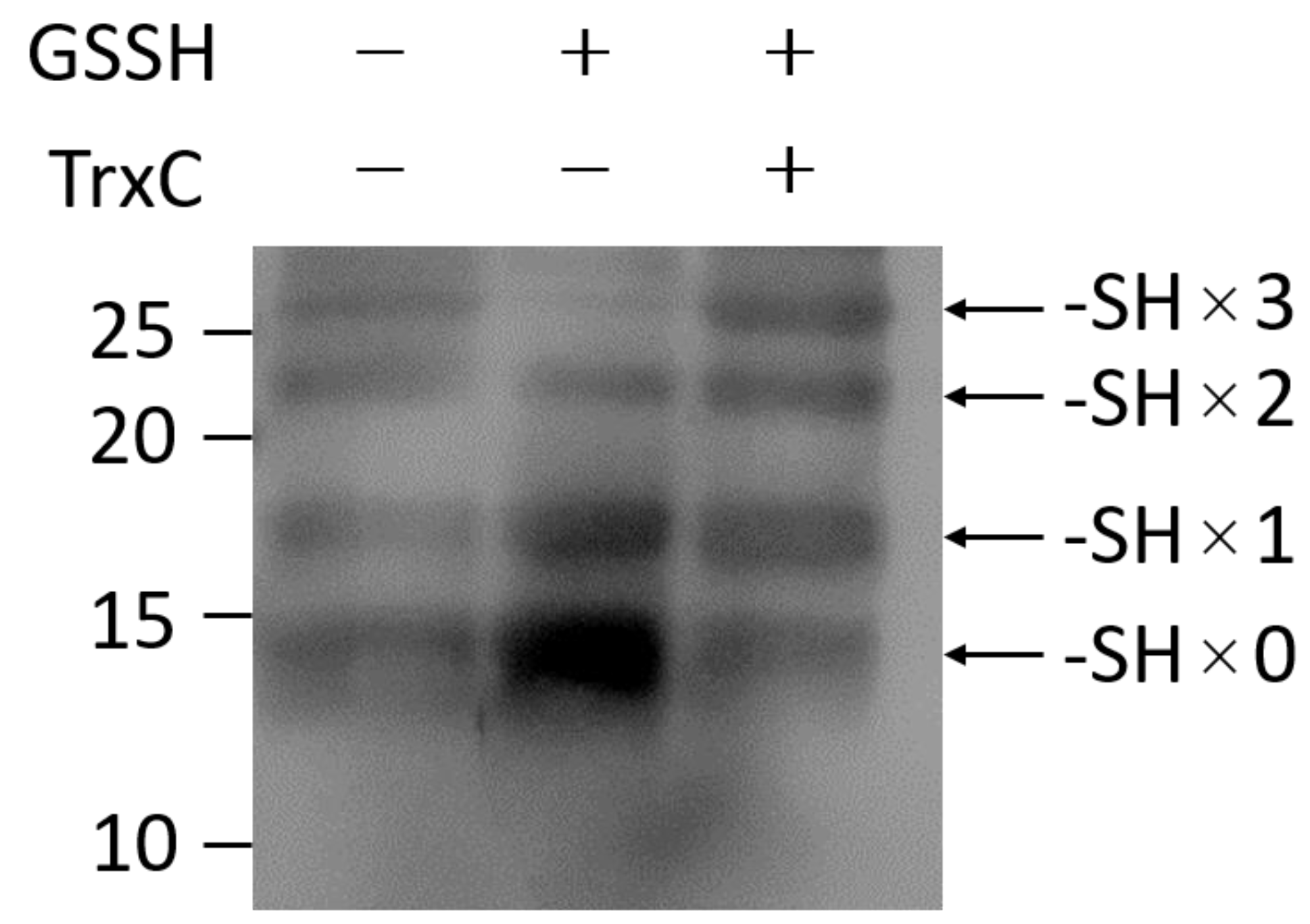
2.4. Analysis of the Redox State of Cysteine Thiols
2.5. Cloning and Mutagenesis
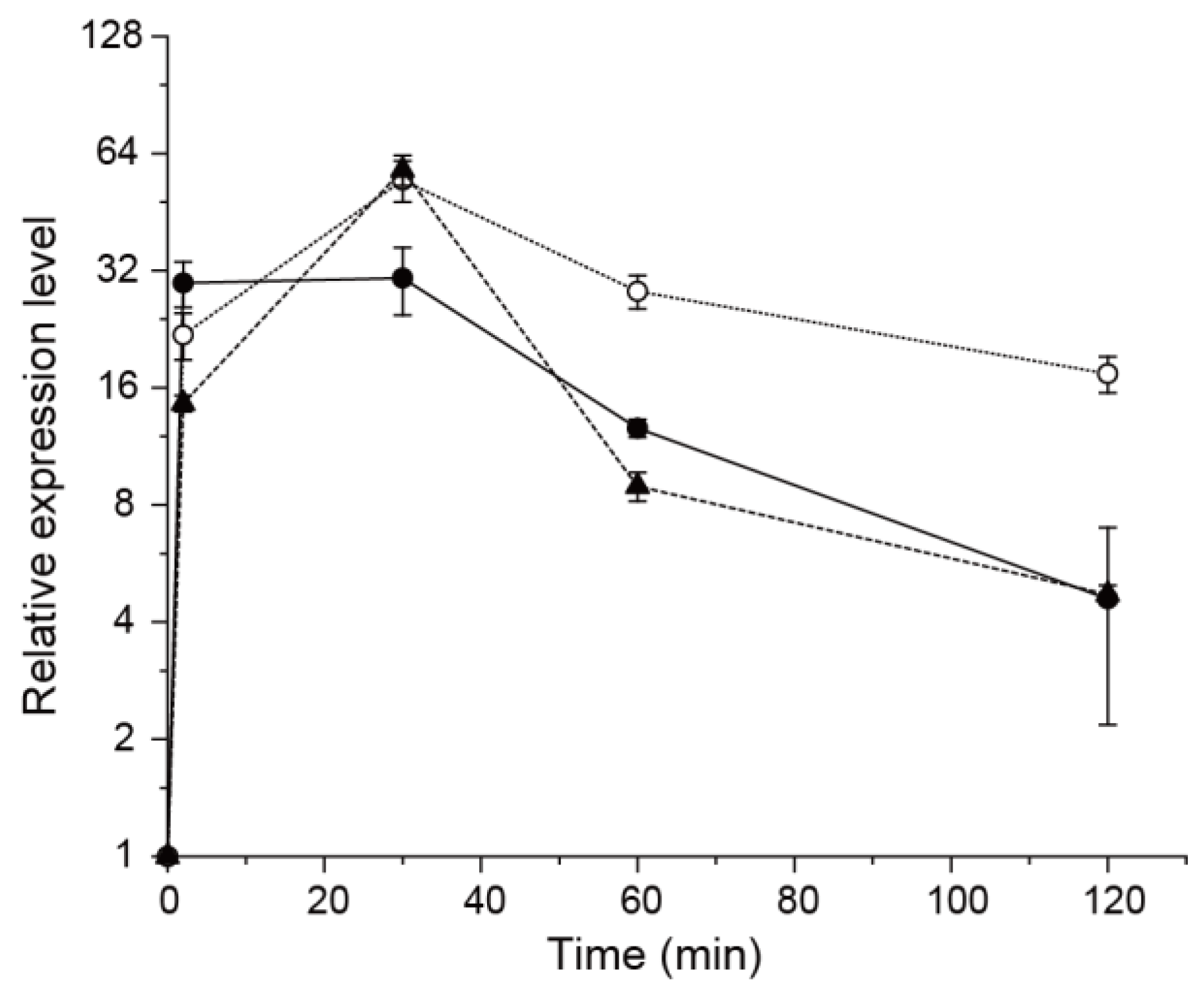
2.6. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
3. Results
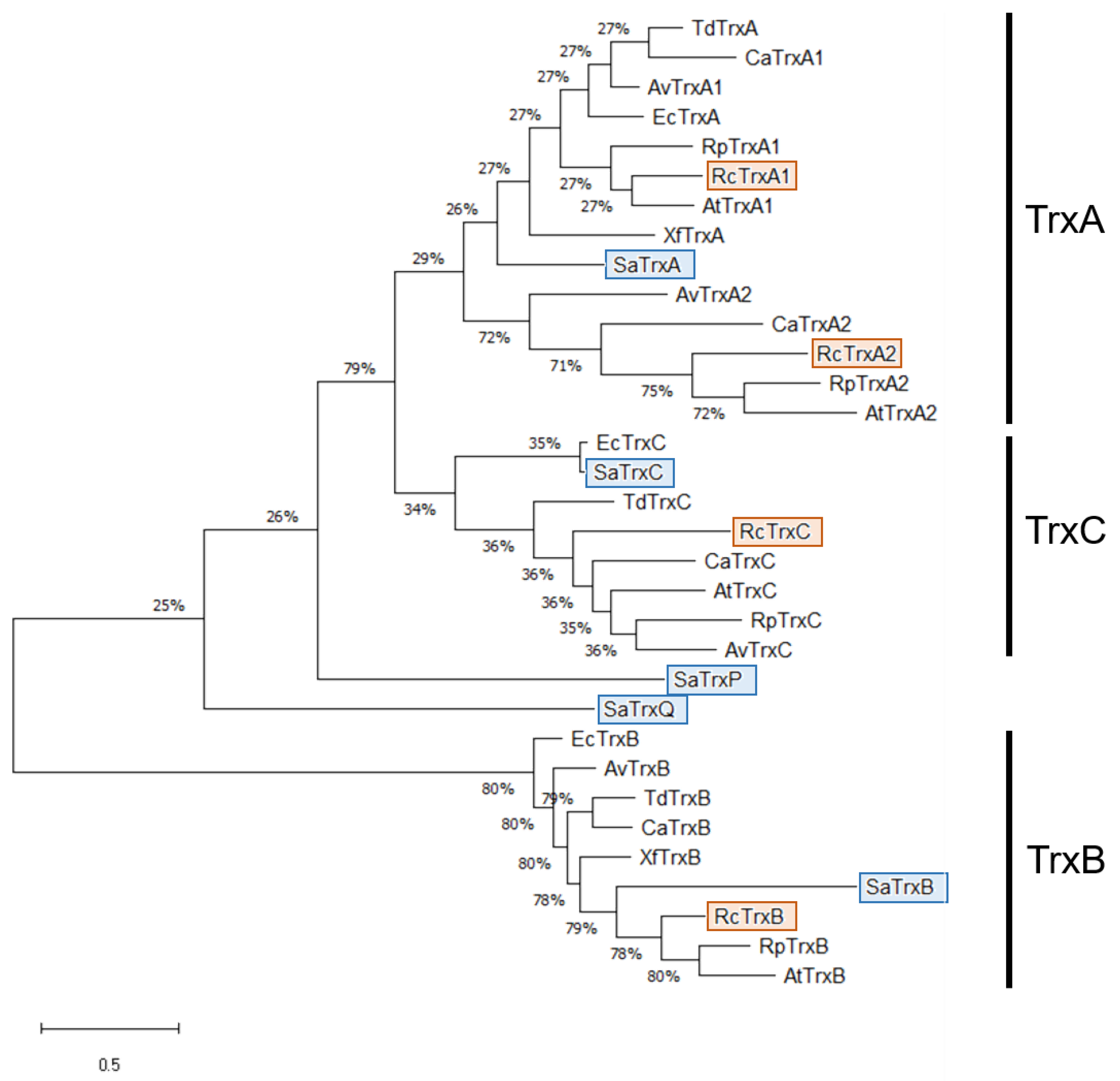
3.1. Identification of TrxC
3.2. Interaction between SqrR and TrxC
3.3. Effect of TrxC on SqrR-Mediated Transcription
4. Discussion

5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive Cysteine Persulfides and S-Polythiolation Regulate Oxidative Stress and Redox Signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, M.; Sawa, T.; Kitajima, N.; Ono, K.; Inoue, H.; Ihara, H.; Motohashi, H.; Yamamoto, M.; Suematsu, M.; Kurose, H.; et al. Hydrogen Sulfide Anion Regulates Redox Signaling via Electrophile Sulfhydration. Nat. Chem. Biol. 2012, 8, 714–724. [Google Scholar] [CrossRef]
- Cuevasanta, E.; Lange, M.; Bonanata, J.; Coitiño, E.L.; Ferrer-Sueta, G.; Filipovic, M.R.; Alvarez, B. Reaction of Hydrogen Sulfide with Disulfide and Sulfenic Acid to Form the Strongly Nucleophilic Persulfide. J. Biol. Chem. 2015, 290, 26866–26880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, P.K.; Martinov, M.; Vitvitsky, V.; Seravalli, J.; Wedmann, R.; Filipovic, M.R.; Banerjee, R. Biosynthesis and Reactivity of Cysteine Persulfides in Signaling. J. Am. Chem. Soc. 2016, 138, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J.; et al. Redox Chemistry and Chemical Biology of H2S, Hydropersulfides, and Derived Species: Implications of Their Possible Biological Activity and Utility. Free. Radic. Biol. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A Novel Persulfide Detection Method Reveals Protein Persulfide- and Polysulfide-Reducing Functions of Thioredoxin and Glutathione Systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.H.; Krokowski, D.; Guan, B.J.; Bederman, I.; Majumder, M.; Parisien, M.; Diatchenko, L.; Kabil, O.; Willard, B.; Banerjee, R.; et al. Quantitative H2S-Mediated Protein Sulfhydration Reveals Metabolic Reprogramming during the Integrated Stress Response. eLife 2015, 4, e10067. [Google Scholar] [CrossRef]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation Proteome Reveals the Regulation of Protein Function by Hydrogen Sulfide in Diverse Biological Processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [Green Version]
- Walsh, B.J.C.; Giedroc, D.P. H2S and Reactive Sulfur Signaling at the Host-Bacterial Pathogen Interface. J. Biol. Chem. 2020, 295, 13150–13168. [Google Scholar] [CrossRef]
- Shimizu, T.; Ida, T.; Antelo, G.T.; Ihara, Y.; Joseph, F.N.; Masuda, S.; Giedroc, D.P.; Akaike, T.; Yamauchi, T.; Tobe, K.; et al. Polysulfide Metabolizing Enzymes Influence SqrR-Mediated Sulfide-Induced Transcription by Impacting Intracellular Polysulfide Dynamics. PNAS Nexus 2023, 2, pgad048. [Google Scholar] [CrossRef]
- Shimizu, T.; Shen, J.; Fang, M.; Zhang, Y.; Hori, K.; Trinidad, J.C.; Bauer, C.E.; Giedroc, D.P.; Masuda, S. SqrR Functions as a Master Regulator of Sulfide-Dependent Photosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, 2355–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capdevila, D.A.; Walsh, B.J.C.; Zhang, Y.; Dietrich, C.; Gonzalez-gutierrez, G.; Giedroc, D.P. Structural Basis for Persulfide-Sensing Specificity in a Transcriptional Regulator. Nat. Chem. Biol. 2020, 17, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Braunstein, I.; Engelman, R.; Yitzhaki, O.; Ziv, T.; Galardon, E.; Benhar, M. Opposing Effects of Polysulfides and Thioredoxin on Apoptosis through Caspase Persulfidation. J. Biol. Chem. 2020, 295, 3590–3600. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Wu, L.; Yang, G. Thioredoxin 1 Regulation of Protein S-Desulfhydration. Biochem. Biophys. Rep. 2016, 5, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Zhang, Y.; Trinidad, J.C.; Giedroc, D.P. Thioredoxin Profiling of Multiple Thioredoxin-like Proteins in Staphylococcus aureus. Front. Microbiol. 2018, 9, 2385. [Google Scholar] [CrossRef]
- Zheng, C.; Guo, S.; Tennant, W.G.; Pradhan, P.K.; Black, K.A.; Dos Santos, P.C. The Thioredoxin System Reduces Protein Persulfide Intermediates Formed during the Synthesis of Thio-Cofactors in Bacillus subtilis. Biochemistry 2019, 58, 1892–1904. [Google Scholar] [CrossRef]
- Wedmann, R.; Onderka, C.; Wei, S.; András Szijártó, I.; Miljkovic, J.; Femic, A.; Lange, M.; Savitsky, S.; Kumar, P.; Torregrossa, R.; et al. Improved Tag-Switch Method Reveals That Thioredoxin Acts as Depersulfidase and Controls the Intracellular Levels of Protein Persulfidation. Chem. Sci. 2016, 7, 3414–3426. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Zhang, Y.; Palmer, L.D.; Kehl-Fie, T.E.; Skaar, E.P.; Trinidad, J.C.; Giedroc, D.P. Hydrogen Sulfide and Reactive Sulfur Species Impact Proteome S-Sulfhydration and Global Virulence Regulation in Staphylococcus aureus. ACS Infect. Dis. 2017, 3, 744–755. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, K.V.; Hiraishi, A.; Shimada, K.; Matsuura, K. Horizontal Transfer of Genes Coding for the Photosynthetic Reaction Centers of Purple Bacteria. J. Mol. Evol. 1997, 45, 131–136. [Google Scholar] [CrossRef]
- Masuda, S.; Bauer, C.E. Null Mutation of HvrA Compensates for Loss of an Essential RelA/SpoT-Like Gene in Rhodobacter capsulatus. J. Bacteriol. 2004, 186, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.Y.; Gest, H.; Bauer, C.E. Chemosensory and Photosensory Perception in Purple Photosynthetic Bacteria Utilize Common Signal Transduction Components. J. Bacteriol. 1997, 179, 5720–5727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iciek, M.; Kowalczyk-Pachel, D.; Bilska-Wilkosz, A.; Kwiecie, I.; Gorny, M.; Włodek, L. S-Sulfhydration as a Cellular Redox Regulation. Biosci. Rep. 2016, 36, e00304. [Google Scholar] [CrossRef] [PubMed]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides Link H2S to Protein Thiol Oxidation. Antioxid. Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R. Reactive Oxygen Species or Reactive Sulfur Species: Why We Should Consider the Latter. J. Exp. Biol. 2020, 223, jeb196352. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ran, M.; Li, X.; Liu, Y.; Xin, Y.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. The Pathway of Sulfide Oxidation to Octasulfur Globules in the Cytoplasm of Aerobic Bacteria. Appl. Environ. Microbiol. 2022, 88, e01941-21. [Google Scholar] [CrossRef]
- Walsh, B.J.C.; Wang, J.; Edmonds, K.A.; Palmer, L.D.; Zhang, Y.; Trinidad, J.C.; Skaar, E.P.; Giedroc, D.P. The Response of Acinetobacter baumannii to Hydrogen Sulfide Reveals Two Independent Persulfide-Sensing Systems and a Connection to Biofilm Regulation. mBio 2020, 11, e01254-20. [Google Scholar] [CrossRef]
- Hou, N.; Yan, Z.; Fan, K.; Li, H.; Zhao, R.; Xia, Y.; Xun, L.; Liu, H. OxyR Senses Sulfane Sulfur and Activates the Genes for Its Removal in Escherichia coli. Redox Biol. 2019, 26, 101293. [Google Scholar] [CrossRef]
- Eklund, H.; Gleason, F.K.; Holmgren, A. Structural and Functional Relations among Thioredoxins of Different Species. Proteins Struct. Funct. Bioinform. 1991, 11, 13–28. [Google Scholar] [CrossRef]
- Ritz, D.; Patel, H.; Doan, B.; Zheng, M.; Åslund, F.; Storz, G.; Beckwith, J. Thioredoxin 2 Is Involved in the Oxidative Stress Response in Escherichia Coli. J. Biol. Chem. 2000, 275, 2505–2512. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Álamo, M.J.; Jurado, J.; Gallardo-Madueño, R.; Monje-Casas, F.; Holmgren, A.; Pueyo, C. Transcriptional Regulation of Glutaredoxin and Thioredoxin Pathways and Related Enzymes in Response to Oxidative Stress. J. Biol. Chem. 2000, 275, 13398–13405. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Pasternak, C.; Klug, G. Expression of the TrxA Gene for Thioredoxin 1 in Rhodobacter sphaeroides during Oxidative Stress. Arch. Microbiol. 2003, 180, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, C.; Haberzettl, K.; Klug, G. Thioredoxin Is Involved in Oxygen-Regulated Formation of the Photosynthetic Apparatus of Rhodobacter sphaeroides. J. Bacteriol. 1999, 181, 100–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Härtig, E.; Klug, G. Thioredoxin 2 Is Involved in Oxidative Stress Defence and Redox-Dependent Expression of Photosynthesis Genes in Rhodobacter capsulatus. Microbiology 2003, 149, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated Software for Molecular Evolutionary Genetics Analysis and Sequence Alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Accession Number) | Sequence 5′–3′ | Purpose |
|---|---|---|
| TrxC-F (ADE86247) | TCGAAGGTAGGCATATGATGGGGGCCAAGATGGCG | Overexpression of recombinant protein |
| TrxC-R | GTACCGAGCTCCATATCAGGCGCGGGCGCCCAGCTTGCCG | |
| trxC F1 | CGACTCTAGAGGATCAAAGATCGGCAGCCGCATCGGCATCTC | Gene disruption |
| trxC R1 | CTTGGCCCCCATCATATTCGCGTTGCGGAATATAT | |
| trxC F2 | ATGATGGGGGCCAAGGGCGCCCGCGCCTGAGAACCCGCGC | |
| trxC R2 | CGGTACCCGGGGATCCCGGCAGGCGTCGCCGACGAAATCGACCGC | |
| rpoZ qF (ADE87042) | GAGATCGCCGATGAAACC | qRT-PCR |
| rpoZ qR | TCGTCGACCTCGATCTGG | |
| sqr qF (ADE84550) | CGCAAGGAAGACAAGGTCAC | |
| sqr qR | CGAGGGCACGAAATGATAC |
| Accession Number | Fold-Change ± lfcSE (with/without Sulfide) | Fold-Change ± lfcSE (ΔsqrR/WT) |
|---|---|---|
| rcc02517 | 22.4 ± 1.2 | 21.6 ± 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, T.; Hashimoto, M.; Masuda, T. Thioredoxin-2 Regulates SqrR-Mediated Polysulfide-Responsive Transcription via Reduction of a Polysulfide Link in SqrR. Antioxidants 2023, 12, 699. https://doi.org/10.3390/antiox12030699
Shimizu T, Hashimoto M, Masuda T. Thioredoxin-2 Regulates SqrR-Mediated Polysulfide-Responsive Transcription via Reduction of a Polysulfide Link in SqrR. Antioxidants. 2023; 12(3):699. https://doi.org/10.3390/antiox12030699
Chicago/Turabian StyleShimizu, Takayuki, Masaru Hashimoto, and Tatsuru Masuda. 2023. "Thioredoxin-2 Regulates SqrR-Mediated Polysulfide-Responsive Transcription via Reduction of a Polysulfide Link in SqrR" Antioxidants 12, no. 3: 699. https://doi.org/10.3390/antiox12030699