
Recent Advances in Biotechnologies for the Treatment of Environmental Pollutants Based on Reactive Sulfur Species

Abstract
:1. Introduction
2. RSS Definition and Relationship with Environmental Management
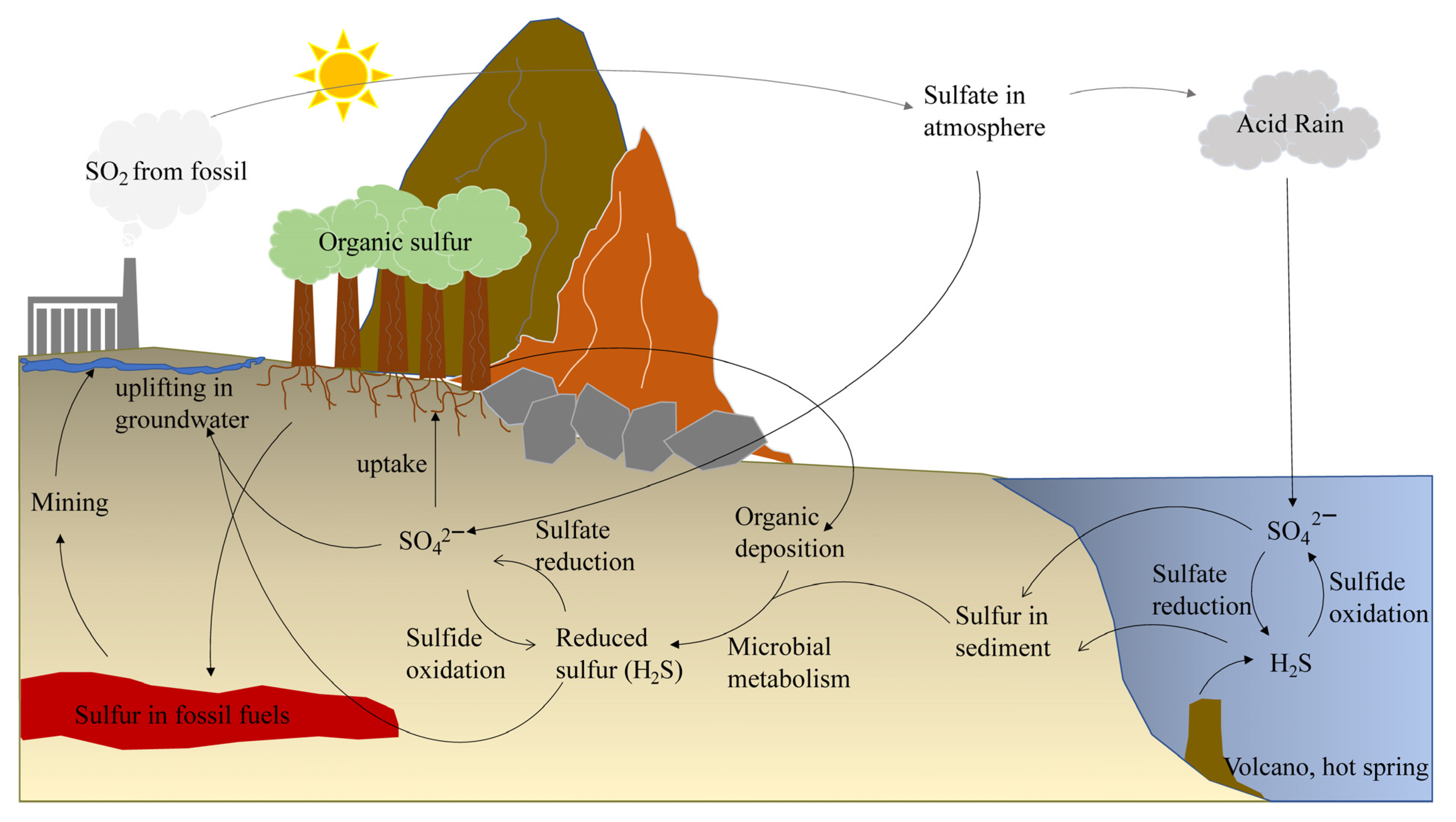
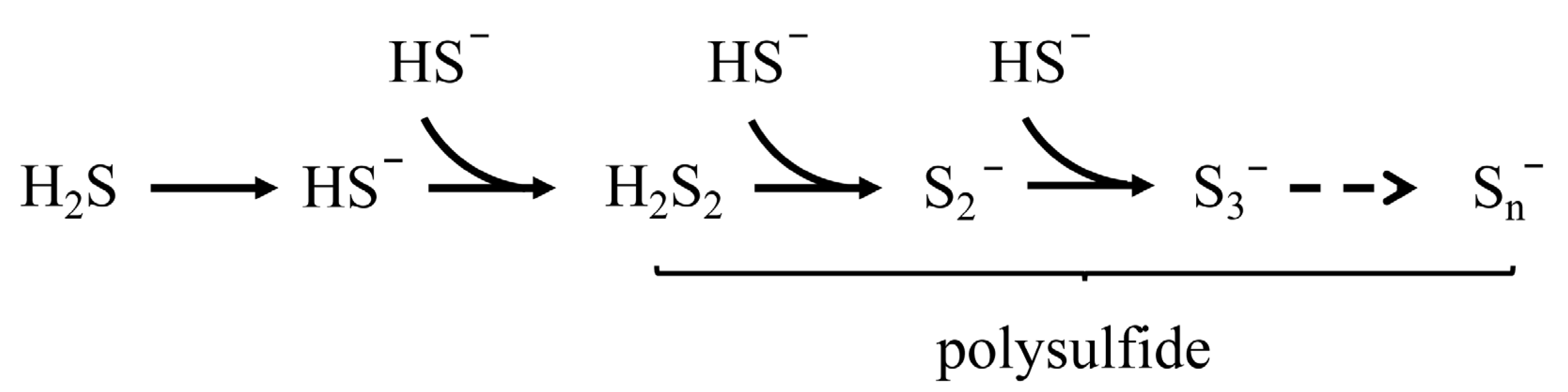
2.1. Origins and Definition of RSS
2.2. Relationship between RSS and Environmental Pollutant Management
3. RSS-Related Bioprocesses for the Treatment of Environmental Pollutants
3.1. Sulfur-Reduction-Based Biological Treatment
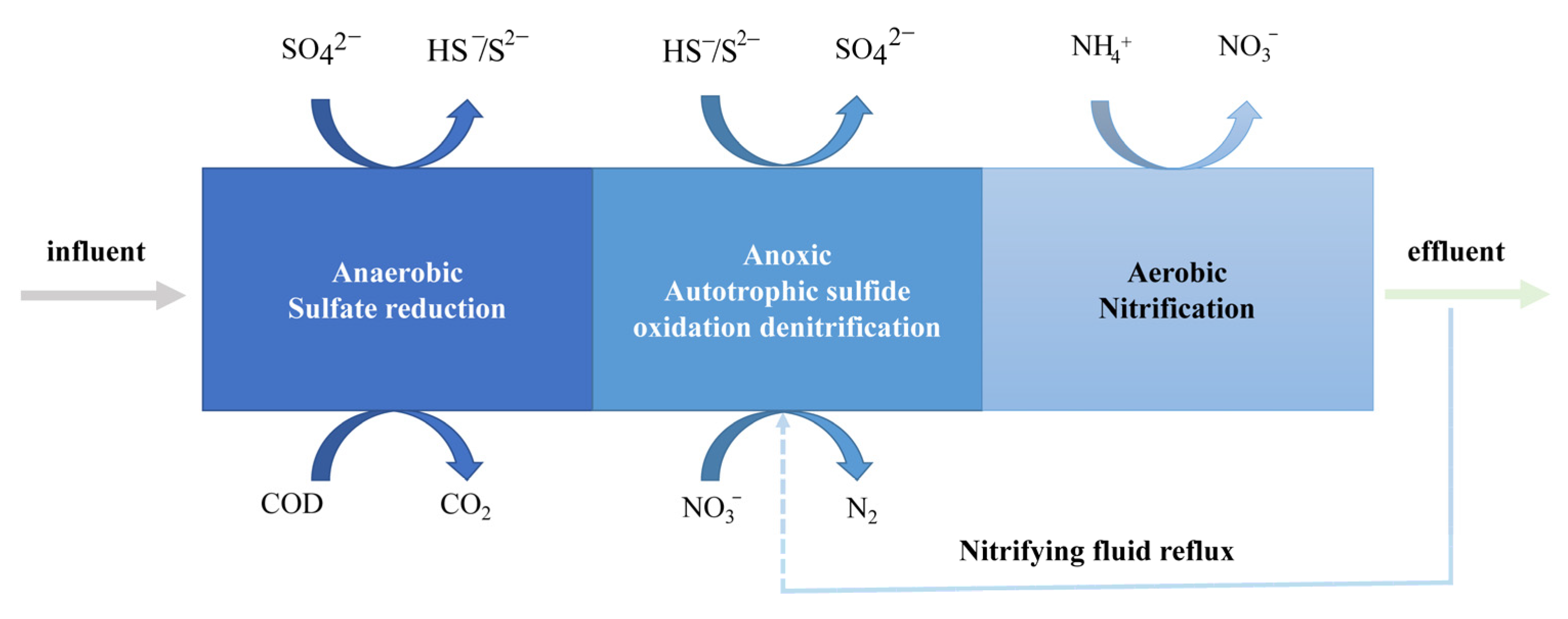
3.1.1. Sulfate-Reduction Bioreactors
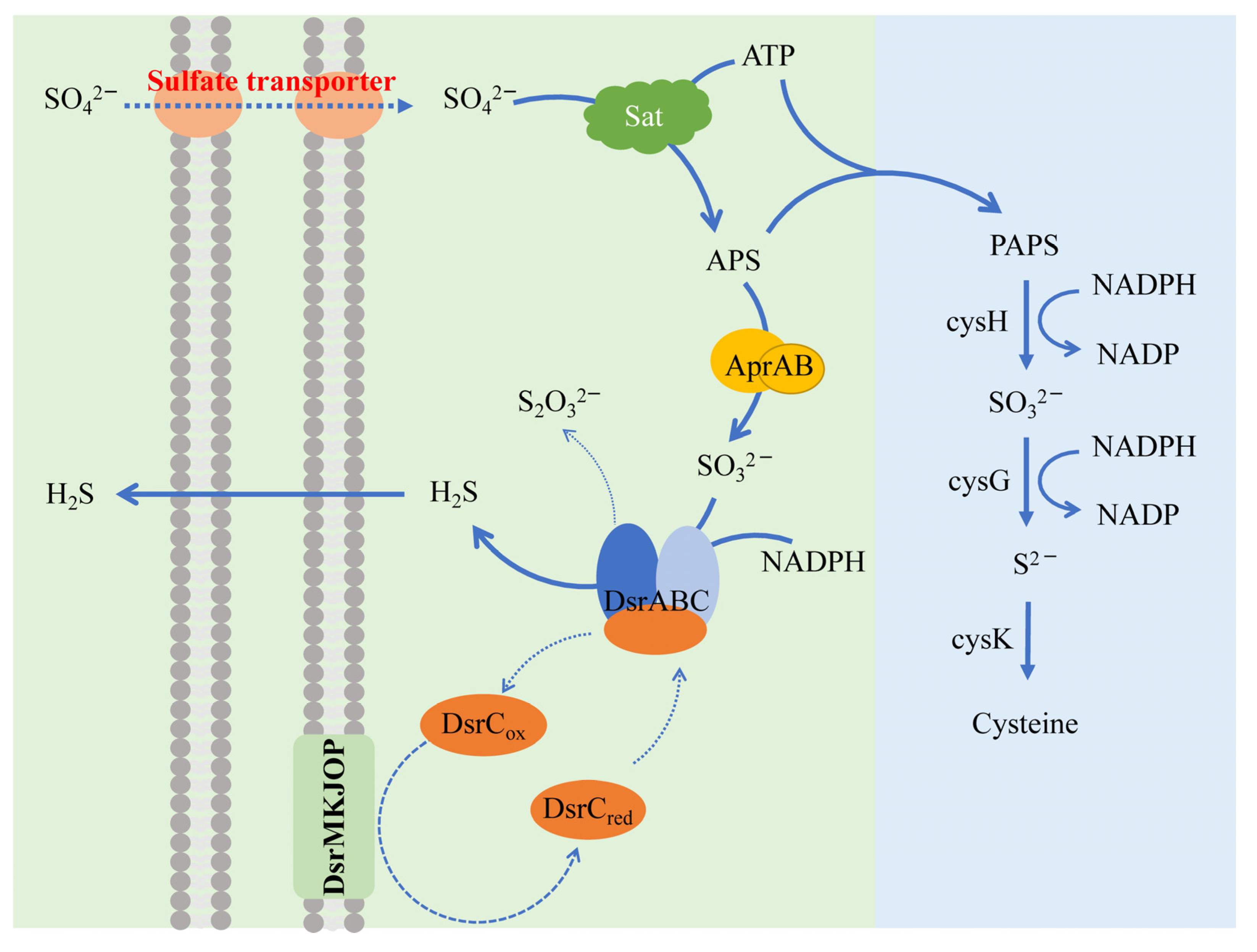
3.1.2. Sulfate-Reducing Bacteria (SRB) and Molecular Mechanisms
3.1.3. S0-Based Reduction Bioreactors
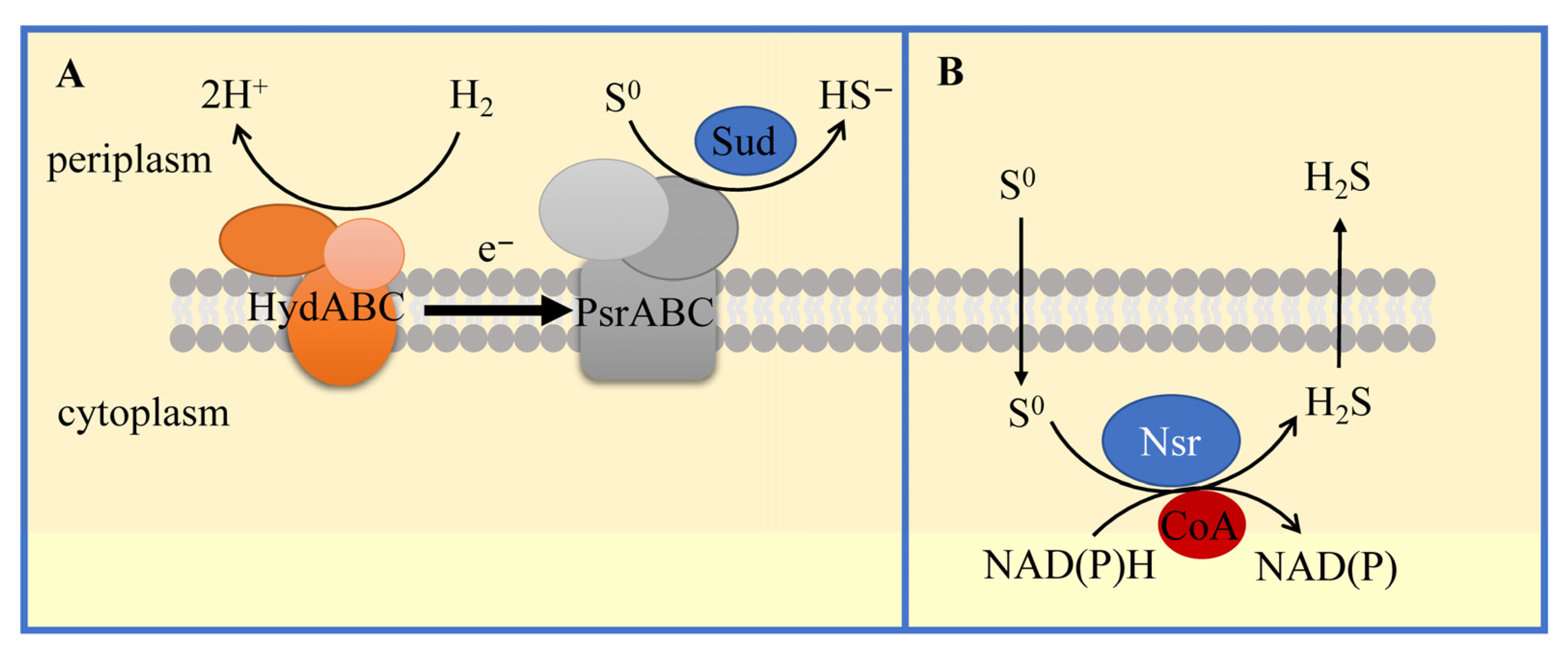
3.1.4. Sulfur-Reducing Bacteria (S0RB) and Molecular Mechanisms
3.2. Sulfur-Oxidation-Based Biological Treatment
3.2.1. Sulfide-Oxidation Bioreactors
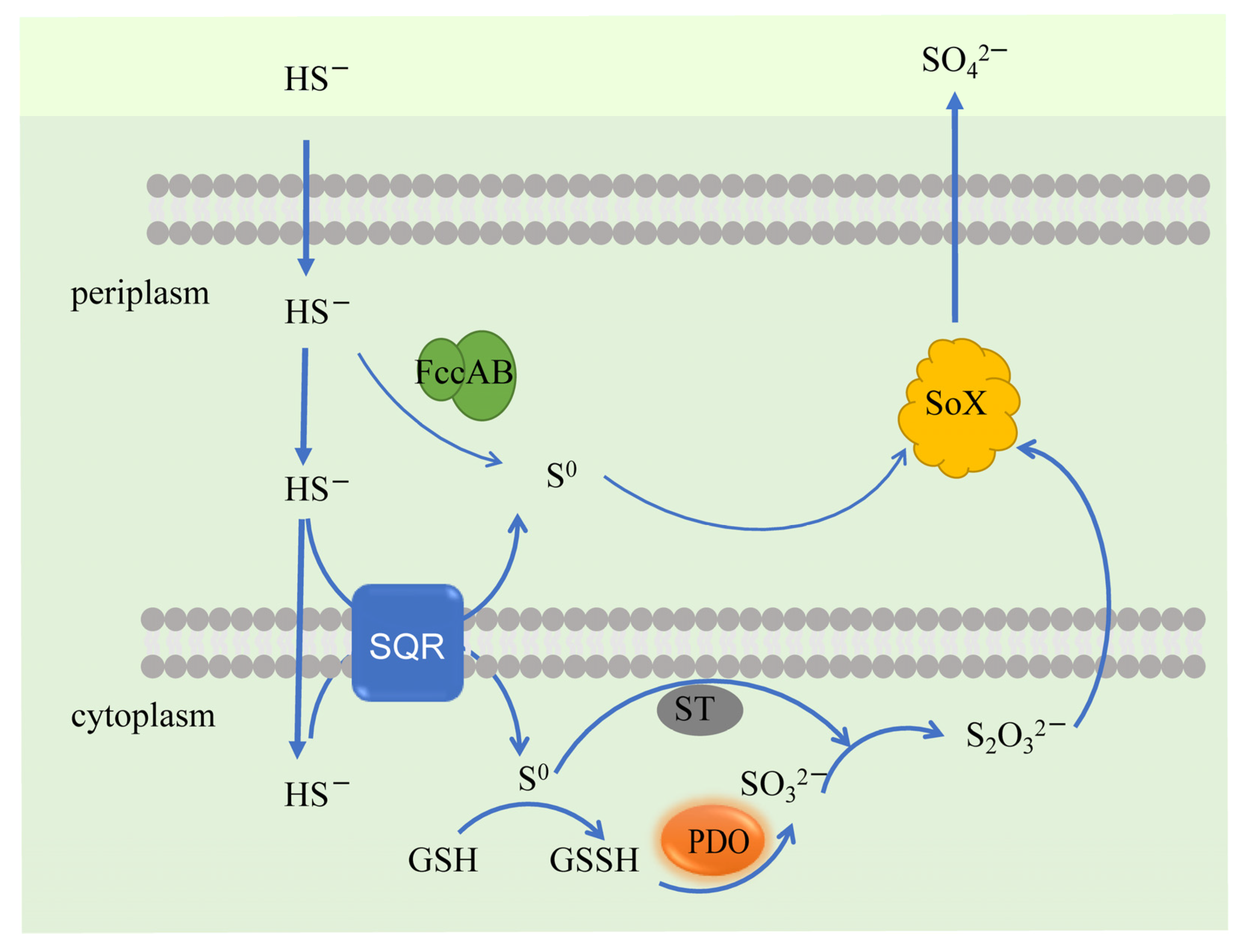
3.2.2. Sulfide-Oxidation Bacteria (SOB) and Molecular Mechanisms
3.2.3. S0-Based Oxidation Bioreactors
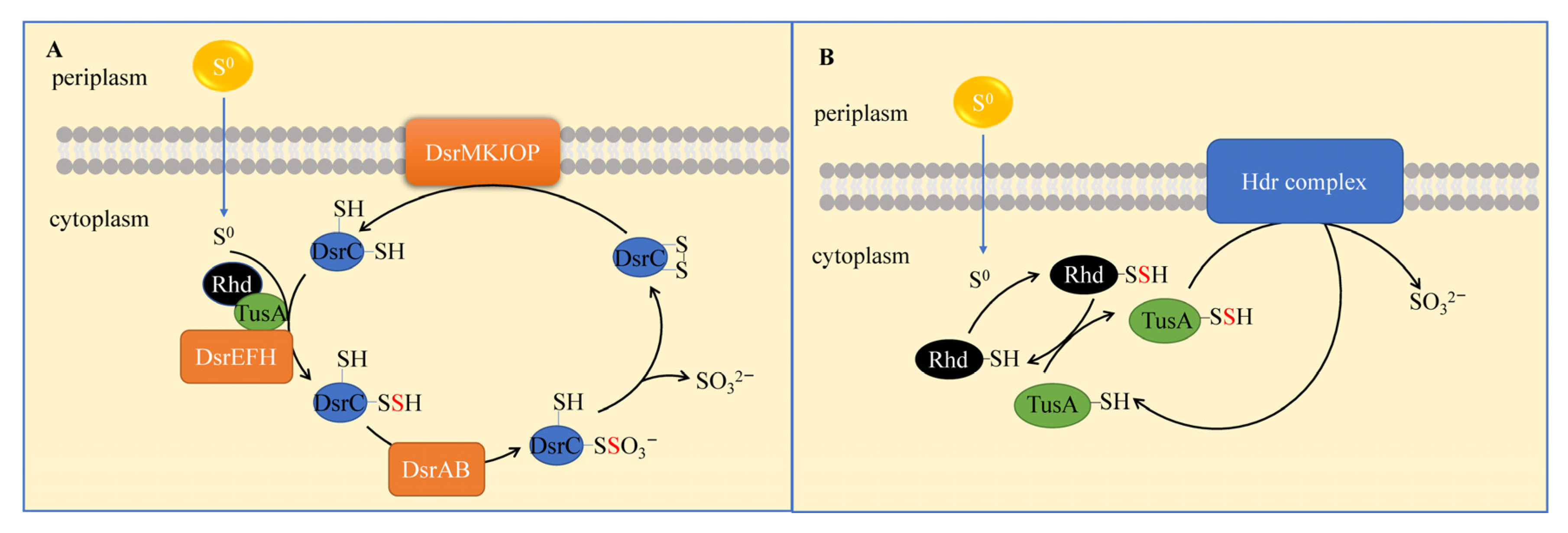
3.2.4. Sulfur-Oxidation Bacteria (S0OB) and Molecular Mechanisms
4. Prospects and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raiswell, R.; Canfield, D.E. The iron biogeochemical cycle past and present. Geochem. Perspect. 2012, 1, 1–220. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R.; Straub, K.D. The role of hydrogen sulfide in evolution and the evolution of hydrogen sulfide in metabolism and signaling. Physiology 2016, 31, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopf, J.W. Geological evidence of oxygenic photosynthesis and the biotic response to the 2400-2200 Ma “Great Oxidation Event”. Biochem.-Mosc. 2014, 79, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P.; Sies, H. The Redox Code. Antioxid. Redox Signal. 2015, 23, 734–746. [Google Scholar] [CrossRef] [Green Version]
- Grman, M.; Nasim, M.J.; Leontiev, R.; Misak, A.; Jakusova, V.; Ondrias, K.; Jacob, C. Inorganic reactive sulfur-nitrogen species: Intricate release mechanisms or cacophony in yellow, blue and red? Antioxidants 2017, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Giles, G.I.; Jacob, C. Reactive sulfur species: An emerging concept in oxidative stress. Biol. Chem. 2002, 383, 375–388. [Google Scholar] [CrossRef]
- Olson, K.R. Reactive oxygen species or reactive sulfur species: Why we should consider the latter. J. Exp. Biol. 2020, 223, 196352. [Google Scholar] [CrossRef]
- Czerewko, M.A.; Cripps, J.C.; Reid, J.M.; Duffell, C.G. Sulfur species in geological materials—sources and quantification. Cem. Concr. Compos. 2003, 25, 657–671. [Google Scholar] [CrossRef]
- Fike, D.A.; Bradley, A.S.; Rose, C.V. Rethinking the Ancient Sulfur Cycle. In Annual Review of Earth and Planetary Sciences; Jeanloz, R., Freeman, K.H., Eds.; Annual Reviews: Palo Alto, CA, USA, 2015; Volume 43, pp. 593–622. [Google Scholar]
- Giles, G.I.; Tasker, K.M.; Jacob, C. Hypothesis: The role of reactive sulfur species in oxidative stress. Free Radic. Biol. Med. 2001, 31, 1279–1283. [Google Scholar] [CrossRef]
- Brannan, R.G. Reactive sulfur species act as prooxidants in liposomal and skeletal muscle model systems. J. Agric. Food Chem. 2010, 58, 3767–3771. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Slusarenko, A.J. The biology of reactive sulfur species (RSS). Plant Physiol. Biochem. 2012, 59, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Ingenbleek, Y. The nutritional relationship linking sulfur to nitrogen in living organisms. J. Nutr. 2006, 136S, 1641S–1651S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, A.; Bailey, S.M. Redox biology of hydrogen sulfide: Implications for physiology, pathophysiology, and pharmacology. Redox Biol. 2013, 1, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Czyzewski, B.K.; Wang, D. Identification and characterization of a bacterial hydrosulphide ion channel. Nature 2012, 483, 155–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, J.S. Bacterial intracellular sulfur globules: Structure and function. J. Plant Biochem. Biotechnol. 2013, 23, 270–280. [Google Scholar] [CrossRef]
- Chen, Z.; Xia, Y.; Liu, H.; Liu, H.; Xun, L. The mechanisms of thiosulfate toxicity against Saccharomyces cerevisiae. Antioxidants 2021, 10, 646. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Xuan, G.; Liu, H.; Xia, Y.; Xun, L. Sulfane sulfur is a strong inducer of the multiple antibiotic resistance regulator MarR in Escherichia coli. Antioxidants 2021, 10, 1778. [Google Scholar] [CrossRef]
- Toohey, J.I.; Cooper, A.J.L. Thiosulfoxide (sulfane) sulfur: New chemistry and new regulatory roles in biology. Molecules 2014, 19, 12789–12813. [Google Scholar] [CrossRef] [Green Version]
- Lau, N.; Pluth, M.D. Reactive sulfur species (RSS): Persulfides, polysulfides, potential, and problems. Curr. Opin. Chem. Biol. 2019, 49, 1–8. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, Y.; Chen, Z.; Xia, Y.; Xun, L.; Liu, H. Sulfide-quinone oxidoreductase is required for cysteine synthesis and indispensable to mitochondrial health. Redox Biol. 2021, 47, 102169. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Z.; Zhang, X.; Xin, Y.; Xia, Y.; Xun, L.; Liu, H. Rhodanese Rdl2 produces reactive sulfur species to protect mitochondria from reactive oxygen species. Free Radic. Biol. Med. 2021, 177, 287–298. [Google Scholar] [CrossRef]
- Su, C.; Ran, X.; Hu, J.; Shao, C. Photocatalytic process of simultaneous desulfurization and denitrification of flue gas by tio2-polyacrylonitrile nanofibers. Environ. Sci. Technol. 2013, 47, 11562–11568. [Google Scholar] [CrossRef] [PubMed]
- Guimera, X.; Mora, M.; Dorado, A.D.; Bonsfills, A.; Gabriel, D.; Gamisans, X. Optimization of SO2 and NOx sequential wet absorption in a two-stage bioscrubber for elemental sulphur valorisation. Environ. Sci. Pollut. Res. 2021, 28, 24605–24617. [Google Scholar] [CrossRef] [PubMed]
- Crowe, S.A.; Paris, G.; Katsev, S.; Jones, C.; Kim, S.; Zerkle, A.L.; Nomosatryo, S.; Fowle, D.A.; Adkins, J.F.; Sessions, A.L.; et al. Sulfate was a trace constituent of Archean seawater. Science 2014, 346, 735–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, T.; Xiang, P.; Mackey, H.R.; Chi, K.; Lu, H.; Chui, H.; van Loosdrecht, M.C.M.; Chen, G. A review of biological sulfate conversions in wastewater treatment. Water Res. 2014, 65, 1–21. [Google Scholar] [CrossRef]
- Zhang, L.; Qiu, Y.; Zhou, Y.; Chen, G.; Loosdrecht, M.C.M.V.; Jiang, F. Elemental sulfur as electron donor and/or acceptor: Mechanisms, applications and perspectives for biological water and wastewater treatment. Water Res. 2021, 202, 117373. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Wang, J. Various electron donors for biological nitrate removal: A review. Sci. Total Environ. 2021, 794, 148699. [Google Scholar] [CrossRef]
- Wang, J.; Lu, H.; Chen, G.; Lau, G.N.; Tsang, W.L.; van Loosdrecht, M.C.M. A novel sulfate reduction, autotrophic denitrification, nitrification integrated (SANI) process for saline wastewater treatment. Water Res. 2009, 43, 2363–2372. [Google Scholar] [CrossRef]
- Jin, Q.; Bethke, C.M. Cellular energy conservation and the rate of microbial sulfate reduction. Geology 2009, 37, 1027–1030. [Google Scholar] [CrossRef]
- Sorensen, J.; Christensen, D.; Jorgensen, B.B. Volatile fatty acids and hydrogen as substrates for sulfate-reducing bacteria in anaerobic marine sediment. Appl. Environ. Microbiol. 1981, 42, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Nandi, M.; Pakshirajan, K. Recent advances in heavy metal recovery from wastewater by biogenic sulfide precipitation. J. Environ. Manage. 2021, 278, 111555. [Google Scholar] [CrossRef]
- Mendez-Garcia, C.; Pelaez, A.I.; Mesa, V.; Sanchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xu, Z.; Wu, J.; Mo, P. Efficiency and mechanisms of antimony removal from wastewater using mixed cultures of iron-oxidizing bacteria and sulfate-reducing bacteria based on scrap iron. Sep. Purif. Technol. 2020, 246, 116756. [Google Scholar] [CrossRef]
- Xi, Y.; Lan, S.; Li, X.; Wu, Y.; Yuan, X.; Zhang, C.; Liu, Y.; Huang, Y.; Quan, B.; Wu, S. Bioremediation of antimony from wastewater by sulfate-reducing bacteria: Effect of the coexisting ferrous ion. Int. Biodeterior. Biodegrad. 2020, 148, 104912. [Google Scholar] [CrossRef]
- Xie, P.; Chen, C.; Zhang, C.; Su, G.; Ren, N.; Ho, S. Revealing the role of adsorption in ciprofloxacin and sulfadiazine elimination routes in microalgae. Water Res. 2020, 172, 115475. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Khanal, S.K.; Shu, H.; Zhang, H.; Chen, G.; Lu, H. Ciprofloxacin degradation in anaerobic sulfate-reducing bacteria (SRB) sludge system: Mechanism and pathways. Water Res. 2018, 136, 64–74. [Google Scholar] [CrossRef]
- Zhang, H.; Song, S.; Jia, Y.; Wu, D.; Lu, H. Stress-responses of activated sludge and anaerobic sulfate-reducing bacteria sludge under long-term ciprofloxacin exposure. Water Res. 2019, 164, 114964. [Google Scholar] [CrossRef]
- Boopathy, R. Anaerobic phenol degradation by microorganisms of swine manure. Curr. Microbiol. 1997, 35, 64–67. [Google Scholar] [CrossRef]
- Abu Laban, N.; Selesi, D.; Jobelius, C.; Meckenstock, R.U. Anaerobic benzene degradation by Gram-positive sulfate-reducing bacteria. Fems Microbiol. Ecol. 2009, 68, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.J.; Lu, Z.Y.; Wang, P.; Li, H.; Huang, Z.Z.; Lin, K.F.; Liu, Y.D. Diversity and degradation mechanism of an anaerobic bacterial community treating phenolic wastewater with sulfate as an electron acceptor. Environ. Sci. Pollut. Res. 2015, 22, 16121–16132. [Google Scholar] [CrossRef]
- Xie, X.; Mueller, N. Enzymes involved in the anaerobic degradation of phenol by the sulfate-reducing bacterium Desulfatiglans anilini. BMC Microbiol. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finke, N.; Vandieken, V.; Jorgensen, B.B. Acetate, lactate, propionate, and isobutyrate as electron donors for iron and sulfate reduction in Arctic marine sediments, Svalbard. Fems Microbiol. Ecol. 2007, 59, 10–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Gittel, A.; Mussmann, M.; Sass, H.; Cypionka, H.; Koenneke, M. Identity and abundance of active sulfate-reducing bacteria in deep tidal flat sediments determined by directed cultivation and CARD-FISH analysis. Environ. Microbiol. 2008, 10, 2645–2658. [Google Scholar] [CrossRef]
- Jochum, L.M.; Chen, X.; Lever, M.A.; Loy, A.; Jorgensen, B.B.; Schramm, A.; Kjeldsen, K.U. Depth distribution and assembly of sulfate-reducing microbial communities in marine sediments of Aarhus bay. Appl. Environ. Microbiol. 2017, 83, e01547-17. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Ren, N.; Wang, A.; Yu, Z.; Lee, D. Microbial community of granules in expanded granular sludge bed reactor for simultaneous biological removal of sulfate, nitrate and lactate. Appl. Microbiol. Biotechnol. 2008, 79, 1071–1077. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, C.; Liang, B.; Huang, C.; Zhao, Y.; Xu, X.; Tan, W.; Zhou, X.; Gao, S.; Sun, D.; et al. Fine-tuning key parameters of an integrated reactor system for the simultaneous removal of COD, sulfate and ammonium and elemental sulfur reclamation. J. Hazard. Mater. 2014, 269, 56–67. [Google Scholar] [CrossRef]
- Madani, R.M.; Liang, J.; Cui, L.; Zhang, D.; Otitoju, T.A.; Elsalahi, R.H.; Song, X. Novel simultaneous anaerobic ammonium and sulfate removal process: A review. Environ. Technol. Innov. 2021, 23, 101661. [Google Scholar] [CrossRef]
- Peck, H.D.J. Enzymatic basis for assimilatory and dissimilatory sulfate reduction. J. Bacteriol. 1961, 82, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Prior, A.; Uhrig, J.F.; Heins, L.; Wiesmann, A.; Lillig, C.H.; Stoltze, C.; Soll, J.; Schwenn, J.D. Structural and kinetic properties of adenylyl sulfate reductase from Catharanthus roseus cell cultures. Biochim. Biophys. Acta Protein Struct. Molecul. Enzymol. 1999, 1430, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Fritz, G.; Buchert, T.; Huber, H.; Stetter, K.O.; Kroneck, P. Adenylylsulfate reductases from archaea and bacteria are 1:1 alpha beta-heterodimeric iron-sulfur flavoenzymes—high similarity of molecular properties emphasizes their central role in sulfur metabolism. Febs Lett. 2000, 473, 63–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.A.; Venceslau, S.S.; Grein, F.; Leavitt, W.D.; Dahl, C.; Johnston, D.T.; Pereira, I.A.C. A protein trisulfide couples dissimilatory sulfate reduction to energy conservation. Science 2015, 350, 1541–1545. [Google Scholar] [CrossRef]
- Mueller, A.L.; Kjeldsen, K.U.; Rattei, T.; Pester, M.; Loy, A. Phylogenetic and environmental diversity of DsrAB-type dissimilatory (bi) sulfite reductases. Isme J. 2015, 9, 1152–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, M.M.; Bisher, M.E.; Pratt, L.M.; Fong, J.; Southam, G.; Pfiffner, S.M.; Reches, Z.; Onstott, T.C. Sulfur isotope enrichment during maintenance metabolism in the thermophilic sulfate-reducing bacterium Desulfotomaculum putei. Appl. Environ. Microbiol. 2009, 75, 5621–5630. [Google Scholar] [CrossRef] [Green Version]
- Wing, B.A.; Halevy, I. Intracellular metabolite levels shape sulfur isotope fractionation during microbial sulfate respiration. Proc. Natl. Acad. Sci. USA. 2014, 111, 18116–18125. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, C.; Wang, A.; Fang, N.; Yuan, Y.; Ren, N.; Lee, D. Enhanced elementary sulfur recovery in integrated sulfate-reducing, sulfur-producing rector under micro-aerobic condition. Bioresour. Technol. 2012, 116, 517–521. [Google Scholar] [CrossRef]
- Beulig, F.; Roy, H.; Glombitza, C.; Jorgensen, B.B. Control on rate and pathway of anaerobic organic carbon degradation in the seabed. Proc. Natl. Acad. Sci. USA. 2018, 115, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Das, B.K.; Gauri, S.S.; Bhattacharya, J. Sweetmeat waste fractions as suitable organic carbon source for biological sulfate reduction. Int. Biodeterior. Biodegrad. 2013, 82, 215–223. [Google Scholar] [CrossRef]
- Hussain, A.; Iqbal, M.A.; Javid, A.; Razaq, A.; Aslam, S.; Hasan, A.; Akmal, M.; Qazi, J.I. Application of fruit wastes as cost-effective carbon sources for biological sulphate reduction. Iran. J. Sci. Technol. Trans. A-Sci. 2019, 43, 33–41. [Google Scholar] [CrossRef]
- Jorgensen, B.B.; Findlay, A.J.; Pellerin, A. The biogeochemical sulfur cycle of marine sediments. Front. Microbiol. 2019, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Mardanov, A.V.; Kadnikov, V.V.; Beletsky, A.V.; Ravin, N.V. Sulfur and methane-oxidizing microbial community in a terrestrial mud volcano revealed by metagenomics. Microorganisms 2020, 8, 1333. [Google Scholar] [CrossRef] [PubMed]
- Shahsavari, S.; Seth, R.; Chaganti, S.R.; Biswas, N. Inhibition of anaerobic biological sulfate reduction process by copper precipitates. Chemosphere 2019, 236, 124246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lin, X.; Zhang, Z.; Chen, G.; Jiang, F. Elemental sulfur as an electron acceptor for organic matter removal in a new high-rate anaerobic biological wastewater treatment process. Chem. Eng. J. 2018, 331, 16–22. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, C.; Hu, C.; Liu, H.; Qu, J. Denitrification of groundwater using a sulfur-oxidizing autotrophic denitrifying anaerobic fluidized-bed MBR: Performance and bacterial community structure. Appl. Microbiol. Biotechnol. 2015, 99, 2815–2827. [Google Scholar] [CrossRef]
- Sun, R.; Li, Y.; Lin, N.; Ou, C.; Wang, X.; Zhang, L.; Jiang, F. Removal of heavy metals using a novel sulfidogenic AMD treatment system with sulfur reduction: Configuration, performance, critical parameters and economic analysis. Environ. Int. 2020, 136, 105457. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, L.; Mu, X.; Li, G.; Guan, X.; Hong, J.; Jiang, F. Overlooked pathways of denitrification in a sulfur-based denitrification system with organic supplementation. Water Res. 2020, 169, 115084. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, X.; Zhang, R.; Shao, B.; Fan, K.; Zhao, L.; Ji, X.; Ren, N.; Lee, D.; Chen, C. The mixed/mixotrophic nitrogen removal for the effective and sustainable treatment of wastewater: From treatment process to microbial mechanism. Water Res. 2022, 226, 119269. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; Sun, R.; Liang, S.; Chen, G.; Jiang, F. Self-accelerating sulfur reduction via polysulfide to realize a high-rate sulfidogenic reactor for wastewater treatment. Water Res. 2018, 130, 161–167. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Li, L.; Chen, G.; Jiang, F. A novel elemental sulfur reduction and sulfide oxidation integrated process for wastewater treatment and sulfur recycling. Chem. Eng. J. 2018, 342, 438–445. [Google Scholar] [CrossRef]
- Li, G.; Liang, Z.; Sun, J.; Qiu, Y.; Qiu, C.; Liang, X.; Zhu, Y.; Wang, P.; Li, Y.; Jiang, F. A pilot-scale sulfur-based sulfidogenic system for the treatment of Cu-laden electroplating wastewater using real domestic sewage as electron donor. Water Res. 2021, 195, 116999. [Google Scholar] [CrossRef] [PubMed]
- Gorny, J.; Billon, G.; Lesven, L.; Dumoulin, D.; Made, B.; Noiriel, C. Arsenic behavior in river sediments under redox gradient: A review. Sci. Total Environ. 2015, 505, 423–434. [Google Scholar] [CrossRef] [PubMed]
- de Matos, L.P.; Costa, P.F.; Moreira, M.; Silva Gomes, P.C.; Silva, S.D.Q.; Alves Gurgel, L.V.; Teixeira, M.C. Simultaneous removal of sulfate and arsenic using immobilized non-traditional SRB mixed culture and alternative low-cost carbon sources. Chem. Eng. J. 2018, 334, 1630–1641. [Google Scholar] [CrossRef]
- Sun, J.; Hong, Y.; Guo, J.; Yang, J.; Huang, D.; Lin, Z.; Jiang, F. Arsenite removal without thioarsenite formation in a sulfidogenic system driven by sulfur reducing bacteria under acidic conditions. Water Res. 2019, 151, 362–370. [Google Scholar] [CrossRef]
- King, J.K.; Harmon, S.M.; Fu, T.T.; Gladden, J.B. Mercury removal, methylmercury formation, and sulfate-reducing bacteria profiles in wetland mesocosms. Chemosphere 2002, 46, 859–870. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L.; Kang, Y.; Chen, G.; Jiang, F. Long-term feeding of elemental sulfur alters microbial community structure and eliminates mercury methylation potential in sulfate-reducing bacteria abundant activated sludge. Environ. Sci. Technol. 2018, 52, 4746–4753. [Google Scholar] [CrossRef]
- Sun, J.; Li, L.; Zhou, G.; Wang, X.; Zhang, L.; Liu, Y.; Yang, J.; Lu, X.; Jiang, F. Biological sulfur reduction to generate H2S as a reducing agent to achieve simultaneous catalytic removal of so2 and no and sulfur recovery from flue gas. Environ. Sci. Technol. 2018, 52, 4754–4762. [Google Scholar] [CrossRef]
- Qiu, Y.; Gong, X.; Zhang, L.; Zhou, S.; Li, G.; Jiang, F. Achieving a novel polysulfide-involved sulfur-based autotrophic denitrification process for high-rate nitrogen removal in elemental sulfur-packed bed reactors. ACS EsT Eng. 2022, 2, 1504–1513. [Google Scholar] [CrossRef]
- Hedderich, R.; Klimmek, O.; Kroger, A.; Dirmeier, R.; Keller, M.; Stetter, K.O. Anaerobic respiration with elemental sulfur and with disulfides. Fems Microbiol. Rev. 1998, 22, 353–381. [Google Scholar] [CrossRef]
- Schauder, R.; Kroger, A. Bacterial sulfur respiration. Arch. Microbiol. 1993, 159, 491–497. [Google Scholar] [CrossRef]
- Segerer, A.; Neuner, A.; Kristjansson, J.K.; Stetter, K.O. Acidianus infernus gen-nov, sp-nov, and Acidianus brierleyi comb-nov—facultatively aerobic, extremely acidophilic thermophilic sulfur-metabolizing archaebacteria. Int. J. Syst. Bacteriol. 1986, 36, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Fischer, F.; Zillig, W.; Stetter, K.O.; Schreiber, G. Chemolithoautotrophic metabolism of anaerobic extremely thermophilic archaebacteria. Nature 1983, 301, 511–513. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.M.; Masuchi, Y.; Robb, F.T.; Ammerman, J.W.; Maeder, D.L.; Yanagibayashi, M.; Tamaoka, J.; Kato, C. Pyrococcus horikoshii sp. nov., a hyperthermophilic archaeon isolated from a hydrothermal vent at the Okinawa Trough. Extremophiles 1998, 2, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Deppenmeier, U.; Lienard, T.; Gottschalk, G. Novel reactions involved in energy conservation by methanogenic archaea. Febs Lett. 1999, 457, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, R.; Wilharm, T.; Huber, D.; Trincone, A.; Burggraf, S.; Konig, H.; Rachel, R.; Rockinger, I.; Fricke, H.; Stetter, K.O. Aquifex pyrophilus gen-nov sp-nov represents a novel group of marine hyperthermophilic hydrogen-oxidizing bacteria. Syst. Appl. Microbiol. 1992, 15, 340–351. [Google Scholar] [CrossRef]
- L’Haridon, S.; Cilia, V.; Messner, P.; Raguénès, G.; Gambacorta, A.; Sleytr, U.B.; Prieur, D.; Jeanthon, C. Desulfurobacterium thermolithotrophum gen. nov., sp. nov., a novel autotrophic, sulphur-reducing bacterium isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 1998, 48, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Pfennig, N.; Biebl, H. Desulfuromonas acetoxidans gen. nov. and sp. nov., a new anaerobic, sulfur-reducing, acetate-oxidizing bacterium. Arch. Microbiol. 1976, 110, 3–12. [Google Scholar] [CrossRef]
- Liesack, W.; Finster, K. Phylogenetic analysis of five strains of gram-negative, obligately anaerobic, sulfur-reducing bacteria and description of Desulfuromusa gen. nov., including Desulfuromusa kysingii sp. nov., Desulfuromusa bakii sp. nov., and Desulfuromusa succinoxidans sp. Int. J. Syst. Evol. Microbiol. 1994, 44, 753–758. [Google Scholar] [CrossRef]
- Huber, R.; Woese, C.R.; Langworthy, T.A.; Kristjansson, J.K.; Stetter, K.O. Fervidobacterium-islandicum sp-nov, a new extremely thermophilic eubacterium belonging to the thermotogales. Arch. Microbiol. 1990, 154, 105–111. [Google Scholar] [CrossRef]
- Caccavo, F.; Lonergan, D.J.; Lovley, D.R.; Davis, M.; Stolz, J.F.; Mcinerney, M.J. Geobacter sulfurreducens sp-nov, a hydrogen-oxidizing and acetate-oxidizing dissimilatory metal-reducing microorganism. Appl. Environ. Microbiol. 1994, 60, 3752–3759. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, R.S.; Penning, N. Reduction of sulfur by spirillum 5175 and syntrophism with Chlorobium. Appl. Environ. Microbiol. 1977, 33, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Windberger, E.; Huber, R.; Trincone, A.; Fricke, H.; Stetter, K.O. Thermotoga thermarum sp-nov and Thermotoga neapolitana occurring in African continental solfataric springs. Arch. Microbiol. 1989, 151, 506–512. [Google Scholar] [CrossRef]
- Huber, R.; Woese, C.R.; Langworthy, T.A.; Fricke, H.; Stetter, K.O. Thermosipho africanus gen-nov, represents a new genus of thermophilic eubacteria within the thermotogales. Syst. Appl. Microbiol. 1989, 12, 32–37. [Google Scholar] [CrossRef]
- Macy, J.M.; Schroder, I.; Thauer, R.K.; Kroger, A. Growth the Wolinella succinogenes on H2S plus fumarate and on formate plus sulfur as energy sources. Arch. Microbiol. 1986, 144, 147–150. [Google Scholar] [CrossRef]
- Jelen, B.; Giovannelli, D.; Falkowski, P.G.; Vetriani, C. Elemental sulfur reduction in the deep-sea vent thermophile, Thermovibrio ammonificans. Environ. Microbiol. 2018, 20, 2301–2316. [Google Scholar] [CrossRef] [PubMed]
- Koschorreck, M. Microbial sulphate reduction at a low pH microbial sulphate reduction at a low pH. Fems Microbiol. Ecol. 2008, 64, 329–342. [Google Scholar] [CrossRef]
- Grubba, D.; Yin, Z.; Majtacz, J.; Al-Hazmi, H.E.; Makinia, J. Incorporation of the sulfur cycle in sustainable nitrogen removal systems-A review. J. Clean Prod. 2022, 372, 133495. [Google Scholar] [CrossRef]
- Huang, C.; Liu, Q.; Li, Z.; Ma, X.; Hou, Y.; Ren, N.; Wang, A. Relationship between functional bacteria in a denitrification desulfurization system under autotrophic, heterotrophic, and mixotrophic conditions. Water Res. 2021, 188, 116526. [Google Scholar] [CrossRef]
- Bi, Z.; Zhang, Q.; Xu, X.; Yuan, Y.; Ren, N.; Lee, D.; Chen, C. Perspective on inorganic electron donor-mediated biological denitrification process for low C/N wastewaters. Bioresour. Technol. 2022, 363, 127890. [Google Scholar] [CrossRef]
- Fernandez, M.; Ramirez, M.; Maria Perez, R.; Manuel Gomez, J.; Canter, D. Hydrogen sulphide removal from biogas by an anoxic biotrickling filter packed with Pall rings. Chem. Eng. J. 2013, 225, 456–463. [Google Scholar] [CrossRef]
- Ramos, I.; Perez, R.; Fdz-Polanco, M. Microaerobic desulphurisation unit: A new biological system for the removal of H2S from biogas. Bioresour. Technol. 2013, 142, 633–640. [Google Scholar] [CrossRef]
- Jung, H.; Kim, D.; Choi, H.; Lee, C. A review of technologies for in-situ sulfide control in anaerobic digestion. Renew. Sust. Energ. Rev. 2022, 157, 112068. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, R.; Huang, Z.; Chen, C.; Xu, X.; Zhou, X.; Yin, T.; Wang, A.; Lee, D.; Ren, N. Performance of a novel IAHD-DSR process with methane and sulfide as co-electron donors. J. Hazard. Mater. 2020, 386, 121657. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Avila, J.S.; Razo-Flores, E.; Gomez, J. Simultaneous biological removal of nitrogen, carbon and sulfur by denitrification. Water Res. 2004, 38, 3313–3321. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ren, N.; Wang, A.; Yu, Z.; Lee, D. Simultaneous biological removal of sulfur, nitrogen and carbon using EGSB reactor. Appl. Microbiol. Biotechnol. 2008, 78, 1057–1063. [Google Scholar] [CrossRef]
- Zhang, R.; Xu, X.; Chen, C.; Xing, D.; Shao, B.; Liu, W.; Wang, A.; Lee, D.; Ren, N. Interactions of functional bacteria and their contributions to the performance in integrated autotrophic and heterotrophic denitrification. Water Res. 2018, 143, 355–366. [Google Scholar] [CrossRef]
- Huang, C.; Li, Z.; Chen, F.; Liu, Q.; Zhao, Y.; Gao, L.; Chen, C.; Zhou, J.; Wang, A. Efficient regulation of elemental sulfur recovery through optimizing working height of upflow anaerobic sludge blanket reactor during denitrifying sulfide removal process. Bioresour. Technol. 2016, 200, 1019–1023. [Google Scholar] [CrossRef]
- Huang, C.; Liu, W.; Li, Z.; Zhang, S.; Chen, F.; Yu, H.; Shao, S.; Nan, J.; Wang, A. High recycling efficiency and elemental sulfur purity achieved in a biofilm formed membrane filtration reactor. Water Res. 2018, 130, 1–12. [Google Scholar] [CrossRef]
- Marques, E.L.S.; Dias, J.C.T.; Gross, E.; de Cerqueira E Silva, A.B.; de Moura, S.R.; Rezende, R.P. Purple sulfur bacteria dominate microbial community in Brazilian limestone cave. Microorganisms 2019, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Gregersen, L.H.; Bryant, D.A.; Frigaard, N. Mechanisms and evolution of oxidative sulfur metabolism in green sulfur bacteria. Front. Microbiol. 2011, 2, 116. [Google Scholar] [CrossRef] [Green Version]
- Karr, E.A.; Sattley, W.M.; Jung, D.O.; Madigan, M.T.; Achenbach, L.A. Remarkable diversity of phototrophic purple bacteria in a permanently frozen Antarctic lake. Appl. Environ. Microbiol. 2003, 69, 4910–4914. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.T. Anoxygenic phototrophic bacteria from extreme environments. Photosynth. Res. 2003, 76, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Eckert, C.A.; Freed, E.; Wawrousek, K.; Smolinski, S.; Yu, J.; Maness, P. Inactivation of the uptake hydrogenase in the purple non-sulfur photosynthetic bacterium Rubrivivax gelatinosus CBS enables a biological water-gas shift platform for H-2 production. J. Ind. Microbiol. Biotechnol. 2019, 46, 993–1002. [Google Scholar] [CrossRef]
- Chan, L.; Morgan-Kiss, R.; Hanson, T.E. Sulfur oxidation in Chlorobium tepidum (syn. Chlorobaculum tepidum) Genetic and proteomic analyses. In Microbial Sulfur Metabolism; Springer: Munster, Germany, 2008; p. 117. [Google Scholar]
- Stout, J.; De Smet, L.; Vergauwen, B.; Savvides, S.; Van Beeumen, J. Structural insights into component SoxY of the thiosulfate-oxidizing multienzyme system of Chlorobaculum thiosulfatiphilum. In Microbial Sulfur Metabolism; Springer: Munster, Germany, 2008; p. 127. [Google Scholar]
- Serrano, W.; Schruebbers, J.; Amann, R.; Fischer, U. Allochromatium humboldtianum sp. nov., isolated from soft coastal sediments. Int. J. Syst. Evol. Microbiol. 2015, 65, 2980. [Google Scholar] [CrossRef] [PubMed]
- Caumette, P.; Baulaigue, R.; Matheron, R. Characterization of Chromatium salexigens sp. nov., a Halophilic Chromatiaceae Isolated from Mediterranean Salinas—ScienceDirect. Syst. Appl. Microbiol. 1988, 10, 284–292. [Google Scholar] [CrossRef]
- Bertini, I.; Gaudemer, A.; Luchinat, C.; Piccioli, M. Electron self-exchange in high-potential iron-sulfur proteins. Characterization of protein I from Ectothiorhodospira vacuolata. Biochemistry 1993, 32, 12887–12893. [Google Scholar] [CrossRef] [PubMed]
- Hensen, D.; Sperling, D.; Trüper, H.G.; Brune, D.C.; Dahl, C. Thiosulphate oxidation in the phototrophic sulphur bacterium Allochromatium vinosum. Mol. Microbiol. 2010, 62, 794–810. [Google Scholar] [CrossRef]
- Harwood, C.S.; Gibson, J. Anaerobic and aerobic metabolism of diverse aromatic compounds by the photosynthetic bacterium Rhodopseudomonas palustris. Appl. Environ. Microbiol. 1988, 54, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Pfennig, N. Rhodocyclus purpureus gen. nov. and sp. nov., a Ring-Shaped, Vitamin B12-Requiring Member of the Family Rhodospirillaceae. Int. J. Syst. Bacteriol. 1978, 30, 283–288. [Google Scholar]
- Dziewit, L.; Baj, J.; Szuplewska, M.; Maj, A.; Tabin, M.; Czyzkowska, A.; Skrzypczyk, G.; Adamczuk, M.; Sitarek, T.; Stawinski, P. Insights into the transposable mobilome of Paracoccus spp. (Alphaproteobacteria). PLoS ONE 2012, 7, e32277. [Google Scholar] [CrossRef] [Green Version]
- Valdés, J.; Pedroso, I.; Quatrini, R.; Dodson, R.J.; Tettelin, H.; Blake, R.; Eisen, J.A.; Holmes, D.S. Acidithiobacillus ferrooxidans metabolism: From genome sequence to industrial applications. BMC Genomics 2008, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Takai, K. Thiomicrospira thermophila sp. nov., a novel microaerobic, thermotolerant, sulfur-oxidizing chemolithomixotroph isolated from a deep-sea hydrothermal fumarole in the TOTO caldera, Mariana Arc, Western Pacific. Int. J. Syst. Evol. Microbiol. 2004, 54, 2325. [Google Scholar]
- Nelson, D.C.; Jrgensen, B.B.; Revsbech, N.P. Growth pattern and yield of a chemoautotrophic Beggiatoa sp. in oxygen-sulfide microgradients. Appl. Environ. Microbiol. 1986, 52, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, M.; Zhang, T.; Fang, H.H. Sulfur-driven autotrophic denitrification: Diversity, biochemistry, and engineering applications. Appl. Microbiol. Biotechnol. 2010, 88, 1027–1042. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lu, C.; Hou, N.; Xin, Y.; Liu, J.; Liu, H.; Xun, L. Sulfide production and oxidation by heterotrophic bacteria under aerobic conditions. ISME J. 2017, 11, 2754–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, Y.; Liu, H.; Cui, F.; Liu, H.; Xun, L. Recombinant Escherichia coli with sulfide: Quinone oxidoreductase and persulfide dioxygenase rapidly oxidises sulfide to sulfite and thiosulfate via a new pathway. Environ. Microbiol. 2016, 18, 5123–5136. [Google Scholar] [CrossRef]
- Lu, C.; Xia, Y.; Liu, D.; Zhao, R.; Gao, R.; Liu, H.; Xun, L. Cupriavidus necator H16 uses flavocytochrome c sulfide dehydrogenase to oxidize self-produced and added sulfide. Appl. Environ. Microbiol. 2017, 83, e01610-17. [Google Scholar] [CrossRef] [Green Version]
- Xin, Y.; Gao, R.; Cui, F.; Lu, C.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. The heterotrophic bacterium Cupriavidus pinatubonensis JMP134 oxidizes sulfide to sulfate with thiosulfate as a key intermediate. Appl. Environ. Microbiol. 2020, 86, e01835-20. [Google Scholar] [CrossRef]
- Fan, K.; Xu, X.; Xu, F.; Shi, J.; Sun, K.; Fedorova, I.; Ren, N.; Lee, D.; Chen, C. A novel intra- and extracellular distribution pattern of elemental sulfur in Pseudomonas sp. C27-driven denitrifying sulfide removal process. Environ. Res. 2022, 213, 113674. [Google Scholar] [CrossRef]
- Jorgensen, B.B. Ecology of the bacteria of the sulphur cycle with special reference to anoxic-oxic interface environments. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1982, 298, 543–561. [Google Scholar] [CrossRef]
- Luther, G.W.I.; Findlay, A.J.; Macdonald, D.J.; Owings, S.M.; Hanson, T.E.; Beinart, R.A.; Girguis, P.R. Thermodynamics and kinetics of sulfide oxidation by oxygen: A look at inorganically controlled reactions and biologically mediated processes in the environment. Front. Microbiol. 2011, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Marazioti, C.; Kornaros, M.; Lyberatos, G. Kinetic modeling of a mixed culture of Pseudomonas denitrificans and Bacillus subtilis under aerobic and anoxic operating conditions. Water Res. 2003, 37, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, R.; Xu, X.; Fang, N.; Wang, A.; Ren, N.; Lee, D. Enhanced performance of denitrifying sulfide removal process at high carbon to nitrogen ratios under micro-aerobic condition. Bioresour. Technol. 2017, 232, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Lohwacharin, J.; Annachhatre, A.P. Biological sulfide oxidation in an airlift bioreactor. Bioresour. Technol. 2010, 101, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, C.; Shao, B.; Wang, W.; Xu, X.; Zhou, X.; Xiang, Y.; Zhao, L.; Lee, D.; Ren, N. Heterotrophic sulfide-oxidizing nitrate-reducing bacteria enables the high performance of integrated autotrophic-heterotrophic denitrification (IAHD) process under high sulfide loading. Water Res. 2020, 178, 115848. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Zhang, L.; Jiang, F. Indirect sulfur reduction via polysulfide contributes to serious odor problem in a sewer receiving nitrate dosage. Water Res. 2016, 100, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Biswal, B.K.; Guo, G.; Deng, Y.; Huang, H.; Chen, G.; Wu, D. Biological nitrogen removal from wastewater using sulphur-driven autotrophic denitrification. Appl. Microbiol. Biotechnol. 2019, 103, 6023–6039. [Google Scholar] [CrossRef]
- Sahinkaya, E.; Yurtsever, A.; Aktas, O.; Ucar, D.; Wang, Z. Sulfur-based autotrophic denitrification of drinking water using a membrane bioreactor. Chem. Eng. J. 2015, 268, 180–186. [Google Scholar] [CrossRef]
- Sahinkaya, E.; Kilic, A.; Duygulu, B. Pilot and full scale applications of sulfur-based autotrophic denitrification process for nitrate removal from activated sludge process effluent. Water Res. 2014, 60, 210–217. [Google Scholar] [CrossRef]
- Jamieson-Hanes, J.H.; Gibson, B.D.; Lindsay, M.B.J.; Kim, Y.; Ptacek, C.J.; Blowes, D.W. Chromium isotope fractionation during reduction of CR(VI) under saturated flow conditions. Environ. Sci. Technol. 2012, 46, 6783–6789. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, B.; Qiu, R.; Lai, C.; Jiang, Y.; He, C.; Guo, J. Microbial chromate reduction coupled to anaerobic oxidation of elemental sulfur or zerovalent iron. Environ. Sci. Technol. 2019, 53, 3198–3207. [Google Scholar] [CrossRef]
- Loy, A.; Duller, S.; Wagner, M. Evolution and ecology of microbes dissimilating sulfur compounds Insights from siroheme sulfite reductases. In Microbial Sulfur Metabolism; Springer: Berlin/Heidelberg, Germany, 2008; pp. 46–59. [Google Scholar] [CrossRef]
- Dahl, C. Cytoplasmic sulfur trafficking in sulfur-oxidizing prokaryotes. IUBMB Life 2015, 67, 268–274. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Chemical Formula | Valence of Sulfur |
|---|---|---|
| Inorganic sulfur species: | ||
| Sulfide | H2S/HS−/S2− | −2 |
| Pyretic sulfur | FeS/FeS2 | −2 and −1 |
| Inorganic polysulfides | H-Sn-H/Sn2− (n ≥ 2) | −1 and 0 |
| Elemental sulfur | S/S8/S0 | 0 |
| Thiosulfate | S2O32− | +2 |
| Sulfur dioxide | SO2 | +4 |
| Sulfite | SO32− | +4 |
| Sulfate | SO42− | +6 |
| Organic sulfur species: | ||
| Reduced organic sulfur compounds | Cysteine, methionine | −2 |
| Organic polysulfides | R-Sn-R/R-SnH (n ≥ 2) | 0 |
| Taxonomic Category | Electron Donor | Reference |
|---|---|---|
| Archaea | ||
| Crenarchaeota: | ||
| Acidianus | H2 | [82] |
| Thermoproteus | H2, peptides, maltose, formate, fumarate, ethanol, malate, methanol, glycogen, starch, amylopectin, formamide | [83] |
| Euryarchaeota: | ||
| Pyrococcus | Complex substrates, amino acids, starch, maltose, pyruvate | [84] |
| Methanococcus | H2, formate | [85] |
| Bacteria | ||
| Aquifex | H2, sulfur, thiosulfate | [86] |
| Desulfurobacterium | H2 | [87] |
| Desulfuromonas | Acetate, pyruvate, ethanol | [88] |
| Desulfuromusa | Acetate, propionate | [89] |
| Fervidobacterium | Sugars, pyruvate, yeast extract | [90] |
| Geobacter | Acetate | [91] |
| Sulfospirillum | H2, formate | [92] |
| Thermotoga | Sugars, peptone, yeast extract, bacterial and archaeal cell homogenates | [93] |
| Thermosipho | Yeast extract, brain heart infusion, peptone, tryptone | [94] |
| Wolinella | H2, formate | [95] |
| Taxonomic Category | Representative Species | Metabolic Features | Sulfur Oxidation Genes | Distributed Environment | Reference |
|---|---|---|---|---|---|
| GSB Chlorobi | Chlorobaculum tepidum, Chlorobaculum thiosulfatiphilum | Obligate phototrophy; S2–, S0, or S2O32− as e− donors for reduction of CO2; extracellular S0 globules; potential mixotrophy | SoxXAYZB, APS reductase, Qmo complex, and Fcc | Anaerobic waters, oceans, soils, the Yellowstone hot springs and sediments | [115,116] |
| PSB Chromatiaceae | Allochromatium warmingi Isochromatium buderi | Photoautotrophy except for Rheinheimera spp.; S2− and S0 as e− donors of photosynthesis; intracellular S0 globules | - | Oceans, stagnant aquifers, eutrophic lakes with water bodies, and extreme environments rich in sulfides | [117,118] |
| Ectothiorhodospiraceae | Allochromatium vinosum Ectothiorhodospira vacuolata | Oxidation of S2− for all the members; extracellular S0 globules; polysulfides under alkaline conditions; some can oxidize S2O32− to SO42− | SoxXAYZB, Sqr, DsrABEFHCMKLJOPNRS, APS reductase, and Fcc | [119,120] | |
| PNSB Alphaproteobacteria | Rhodopseudomonas palustris | The preferred photoheterotrophy under anaerobic conditions; photolithoautotrophy with S2−/S2O32− | SoxXAYZBCD, SoxEF, and Sqr | Waste ponds, coastal lagoons and other aquatic-habitat stagnant areas, sediments, wet soils, and rice paddies | [121] |
| Betaproteobacteria | Rhodocyclus purpureus | Chemoorganotrophy/chemolithoautotrophy under aerobic or microaerobic conditions | - | [122] | |
| CSB Alphaproteobacteria | Paracoccus spp. | Facultative chemolithoautotrophy; oxidation of S2−, S0, S2O32−, or SO32− to SO42− | SoxXAYZBCD and SoxEF | Activated sludge, wastewater treatment systems, farmland, and natural ecological environment such as orchards | [123] |
| Acidithiobacillia | Acidithiobacillus ferrooxidans | Obligate chemolithoautotrophy; oxidation of S0, S2O32−, or S4O62− by the incomplete Sox system; S0 globules as intermediates | SoxXAYZB and Sqr | [124] | |
| Gammaproteobacteria | Thiomicrospira crunogena | Obligate chemolithoautotrophy; extracellular S0 globules under low oxygen/pH; transient accumulation of SO32− or polythionate during S0 globules or S2O32− oxidation | SoxXAYZBCD and Sqr | [125] | |
| Gammaproteobacteria | Beggiatoa spp. | Chemolithoheterotrophy/mixotrophy; intracellular S0 globules | Dsr, Sqr, and APS reductase | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, K.; Wang, W.; Xu, X.; Yuan, Y.; Ren, N.; Lee, D.-J.; Chen, C. Recent Advances in Biotechnologies for the Treatment of Environmental Pollutants Based on Reactive Sulfur Species. Antioxidants 2023, 12, 767. https://doi.org/10.3390/antiox12030767
Fan K, Wang W, Xu X, Yuan Y, Ren N, Lee D-J, Chen C. Recent Advances in Biotechnologies for the Treatment of Environmental Pollutants Based on Reactive Sulfur Species. Antioxidants. 2023; 12(3):767. https://doi.org/10.3390/antiox12030767
Chicago/Turabian StyleFan, Kaili, Wei Wang, Xijun Xu, Yuan Yuan, Nanqi Ren, Duu-Jong Lee, and Chuan Chen. 2023. "Recent Advances in Biotechnologies for the Treatment of Environmental Pollutants Based on Reactive Sulfur Species" Antioxidants 12, no. 3: 767. https://doi.org/10.3390/antiox12030767