

Identification and Expression Profiling of Two Saudi Arabia Catalase Genes from Wheat and Barley in Response to Abiotic and Hormonal Stresses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Isolation and Molecular Cloning of TtCAT1 and HvCAT1 cDNA
2.3. Bioinformatic Analysis of the Sequence of Catalase 1 Genes
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. CAT Activity Assay
- AS0 means the absorbance of the blank control;
- and AS2 stand for the absorbance of the samples;
- VT means the total volume of the crude enzyme solution (mL);
- represents the volume of the detected crude enzyme solution (mL);
- FW means the fresh weight of the sample (g), and t means the time from adding H2O2 to the last time (min).
- The activity of the catalase was calculated by the activity level of inoculation minus the level of the control at each corresponding time point.
3. Results
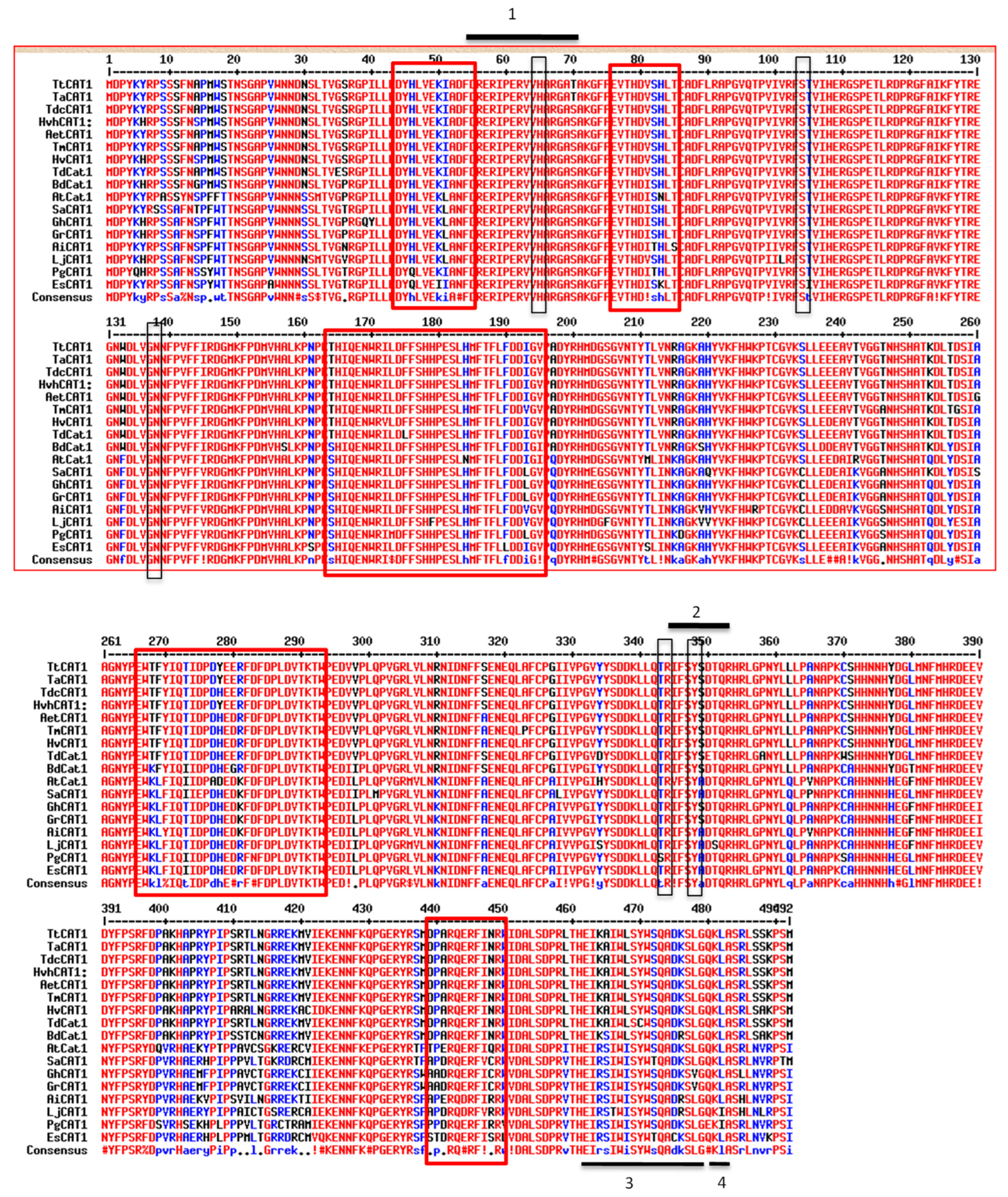
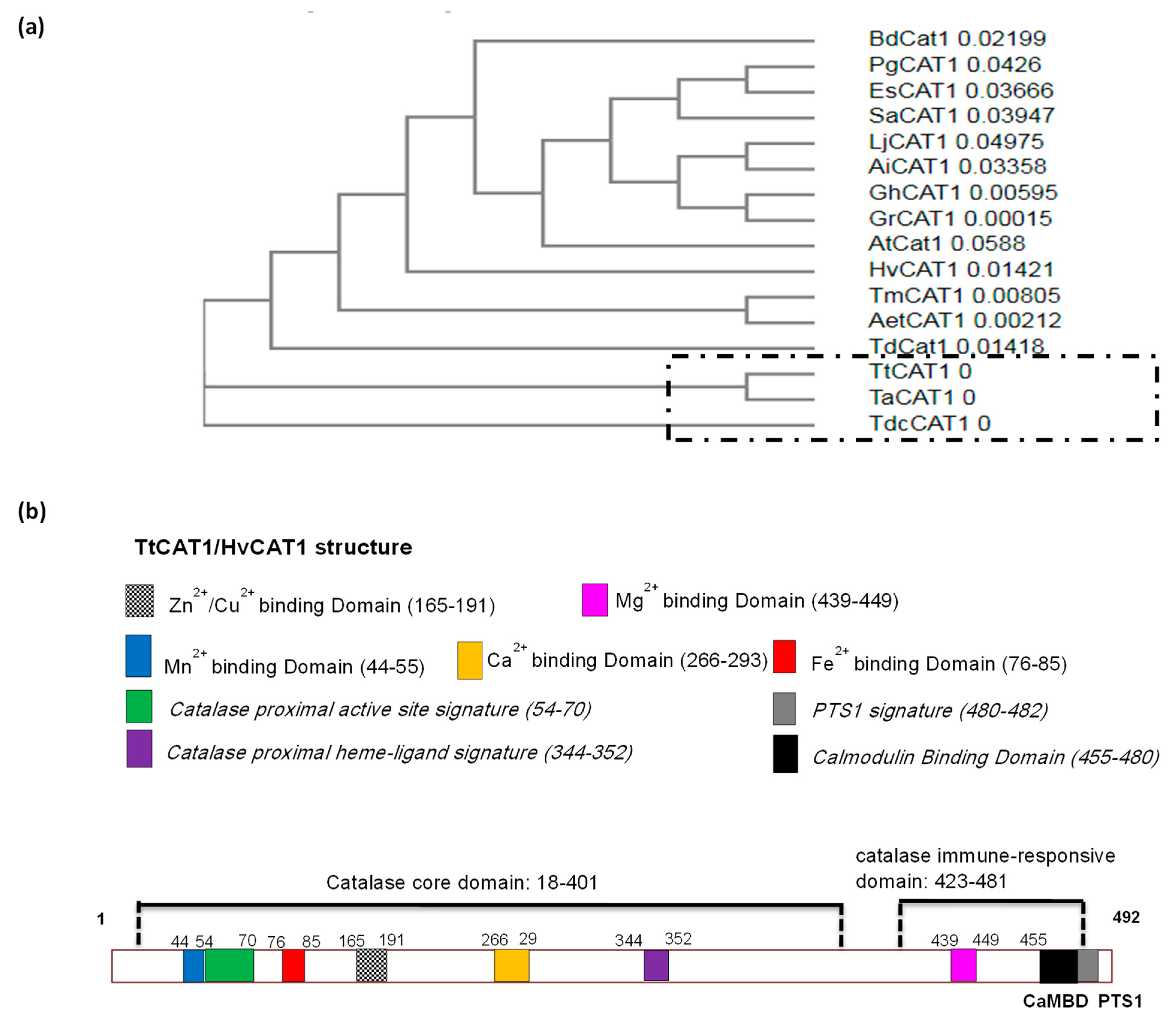
3.1. Isolation and Sequence Analysis of the TtCAT1 and HvCAT1 Genes
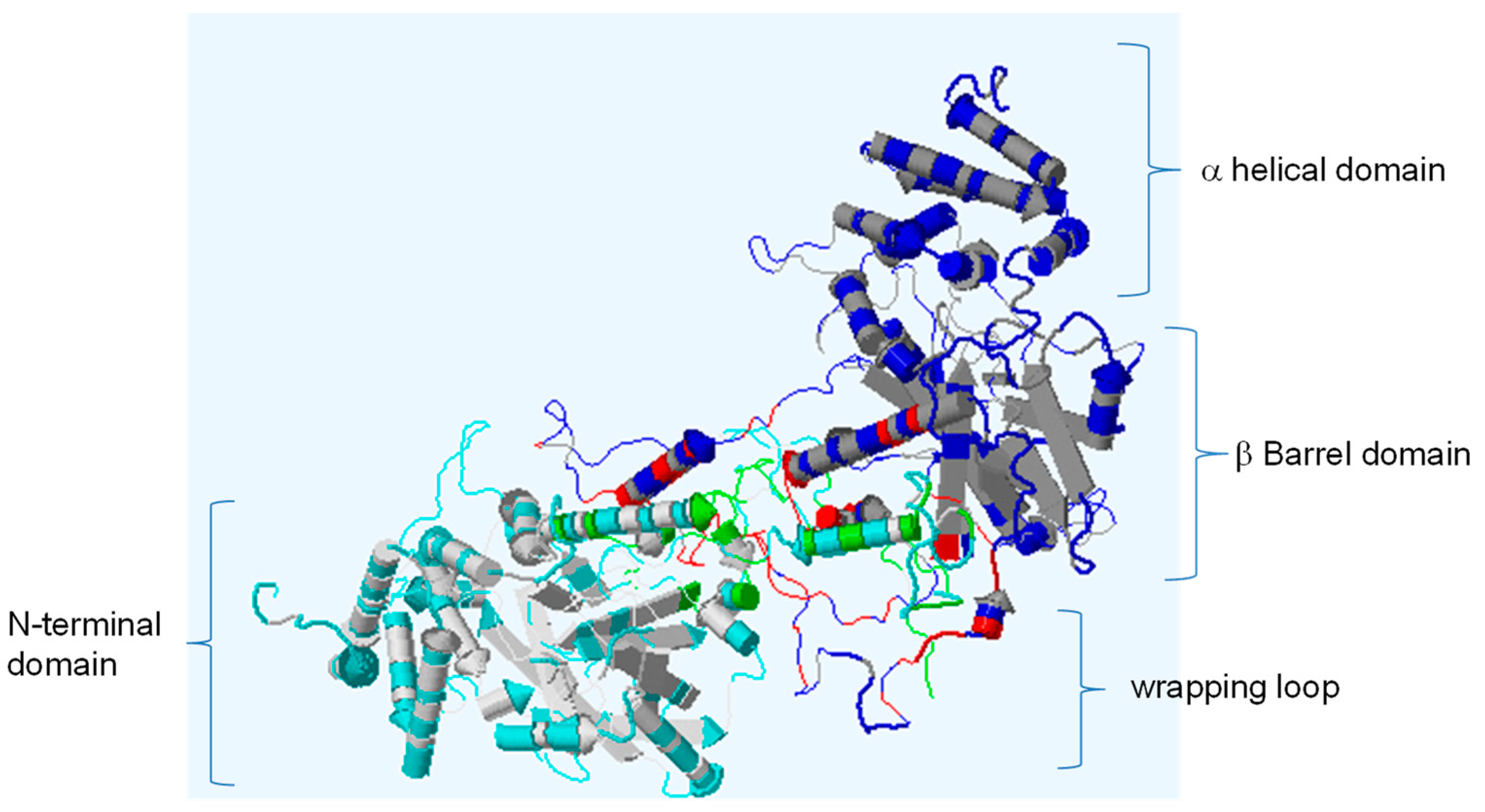
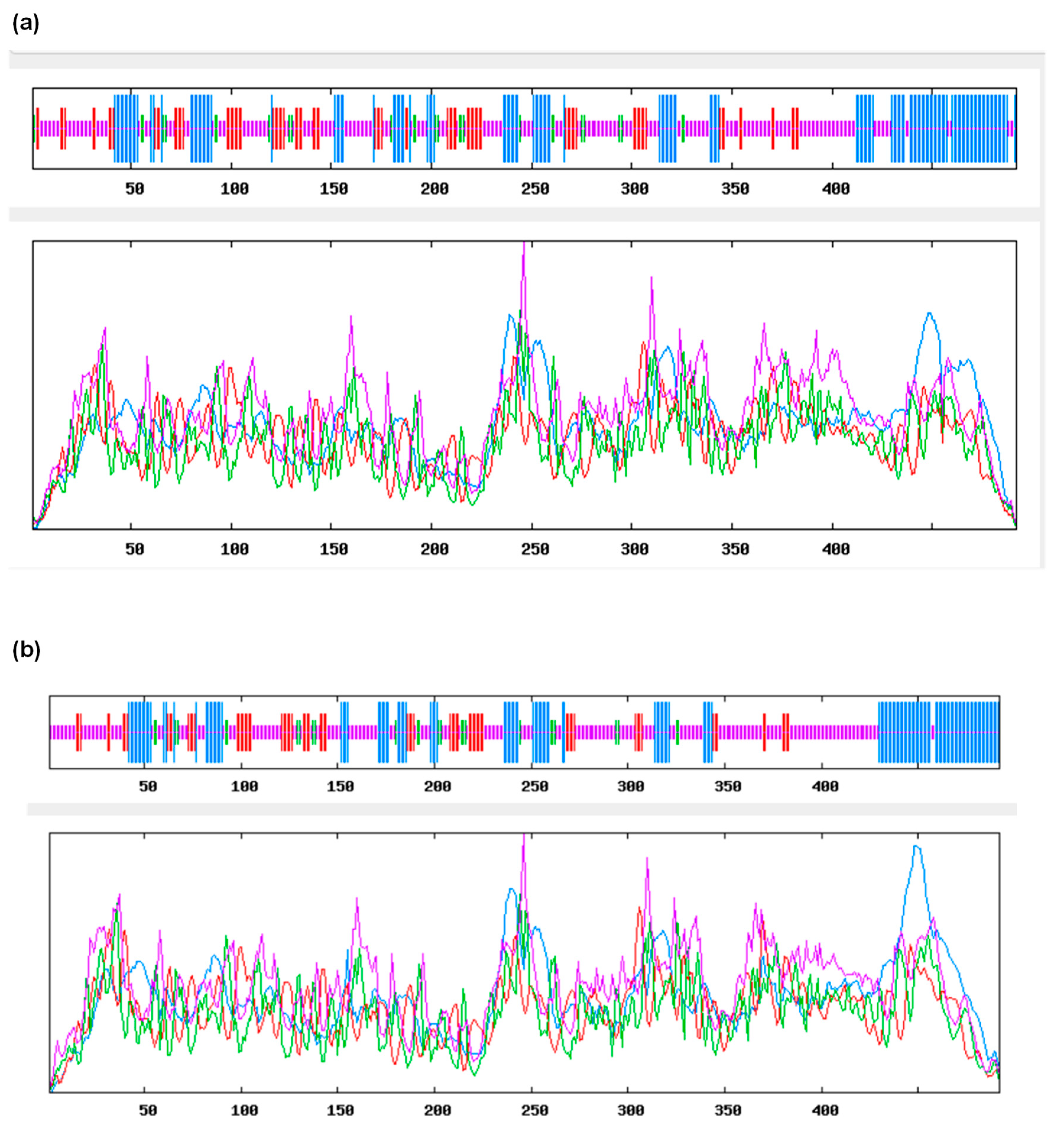
3.2. Structural Homology of TtCAT1 and HvCAT1 Proteins
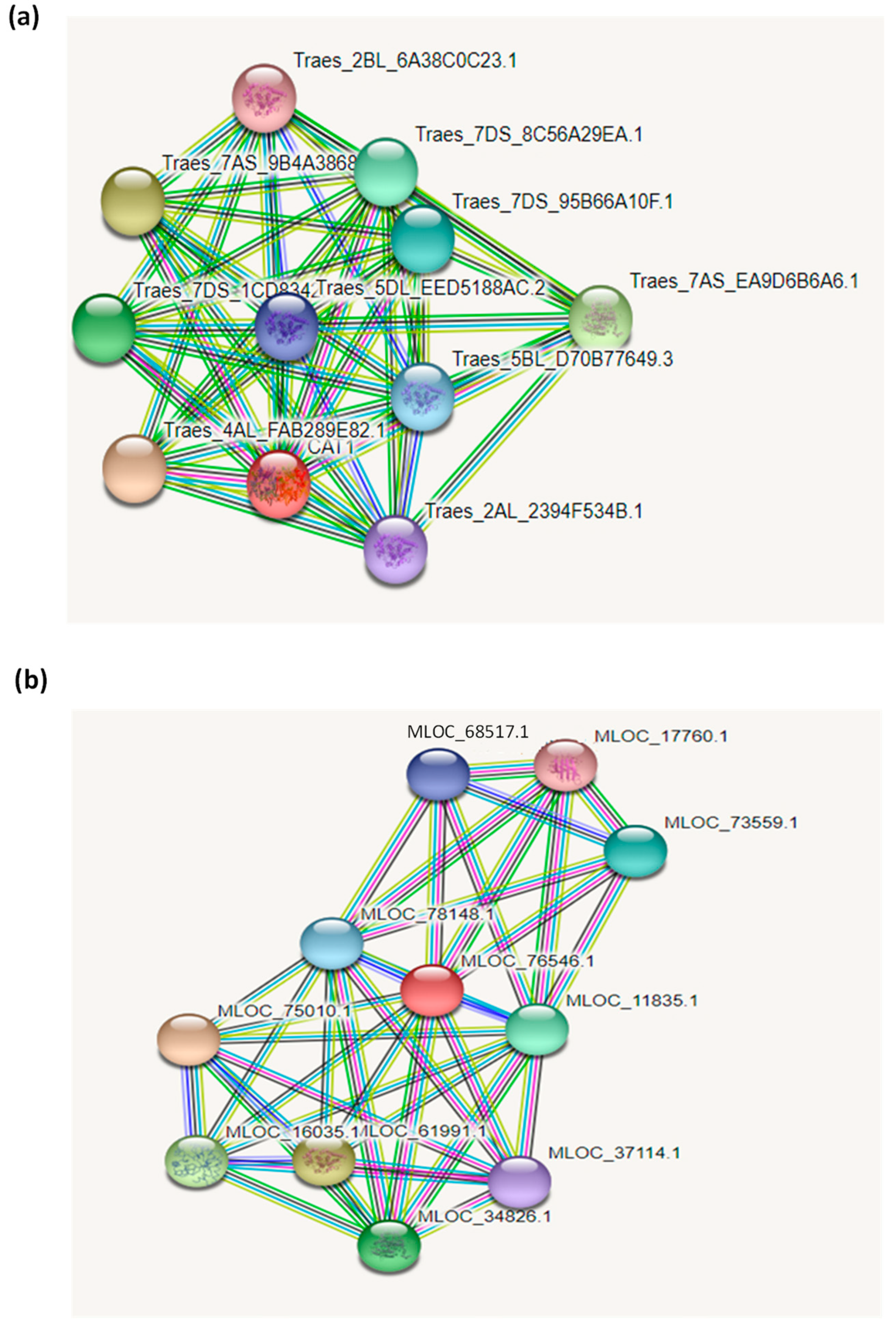
3.3. Interaction Network of Catalase Proteins
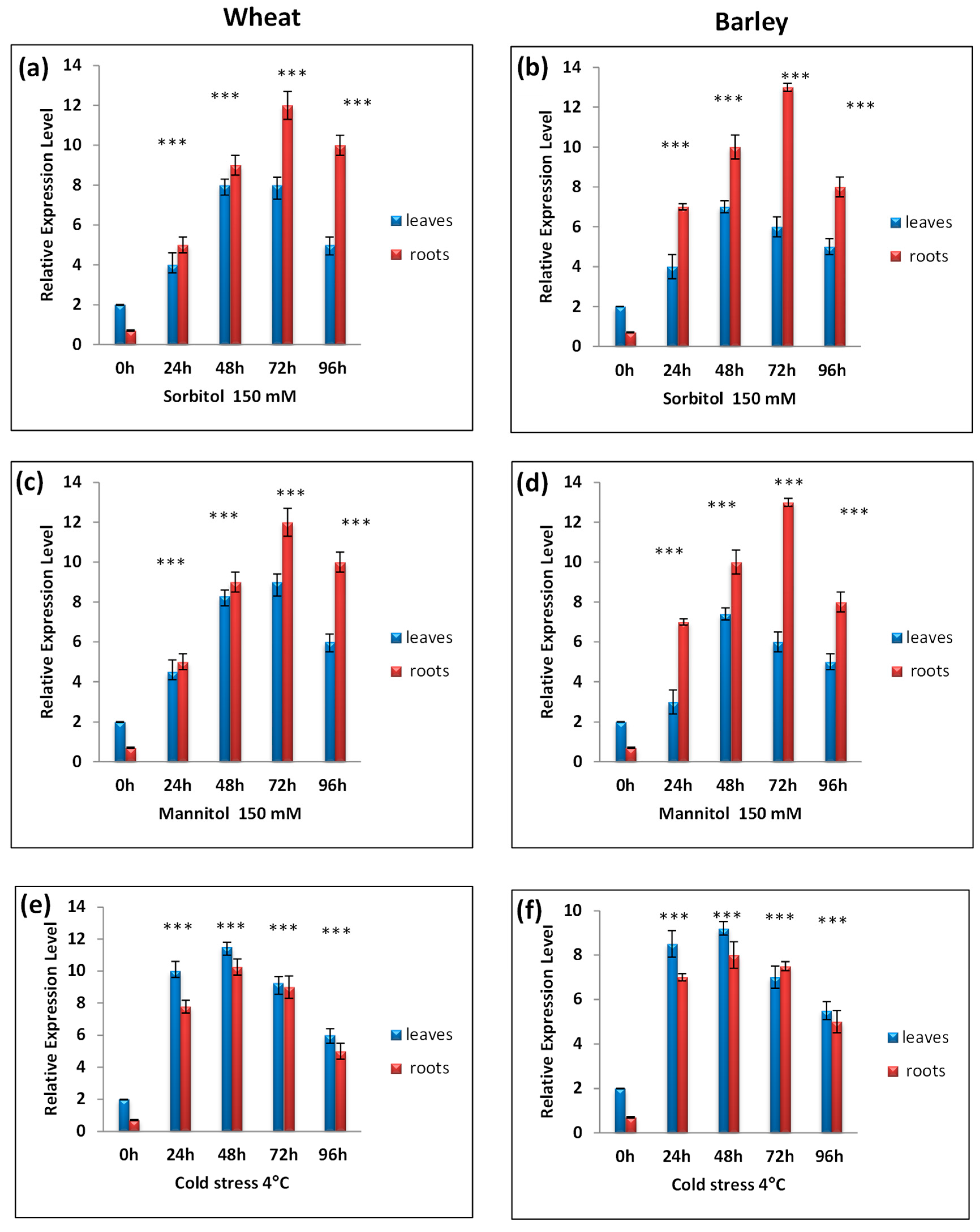
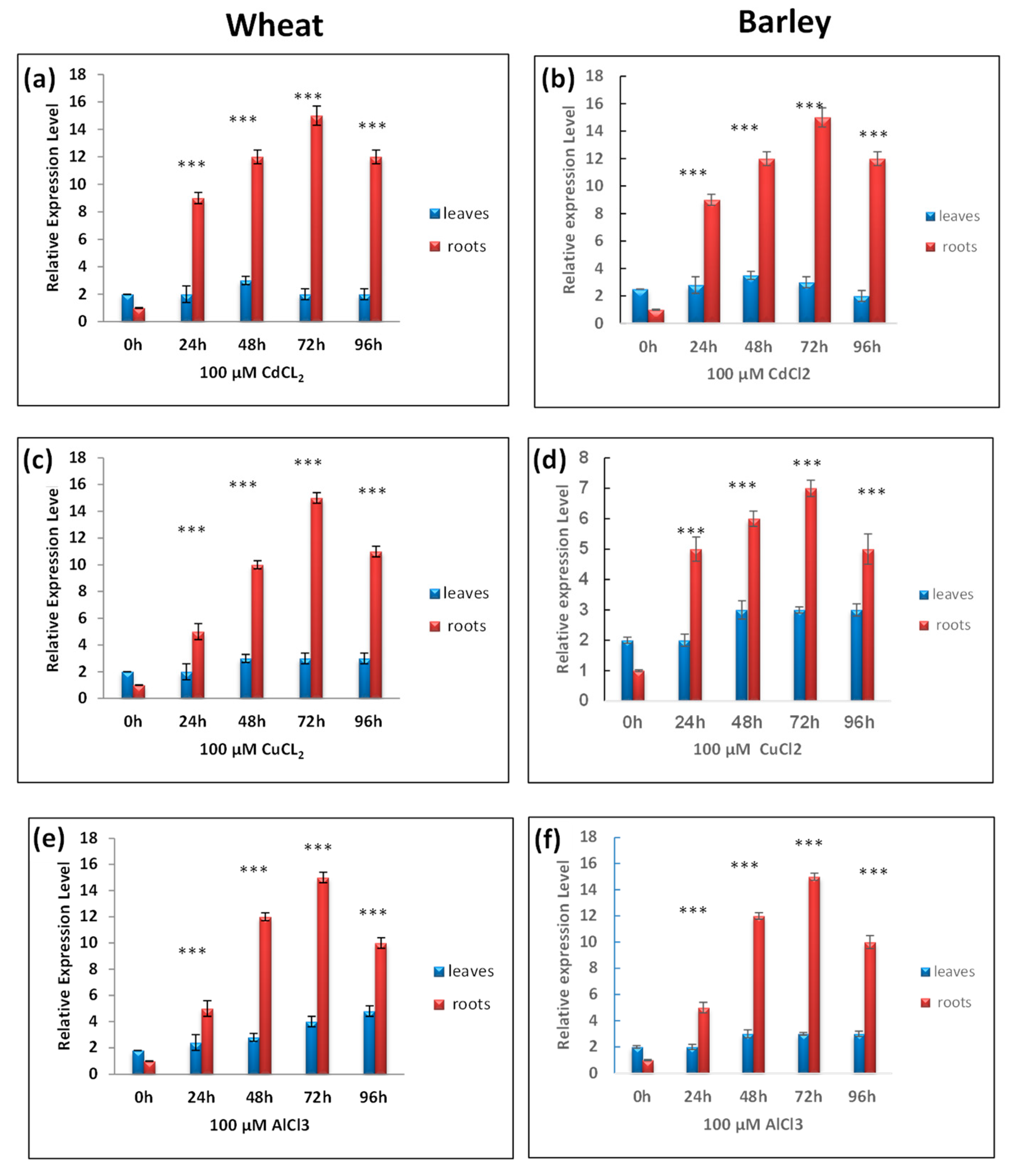
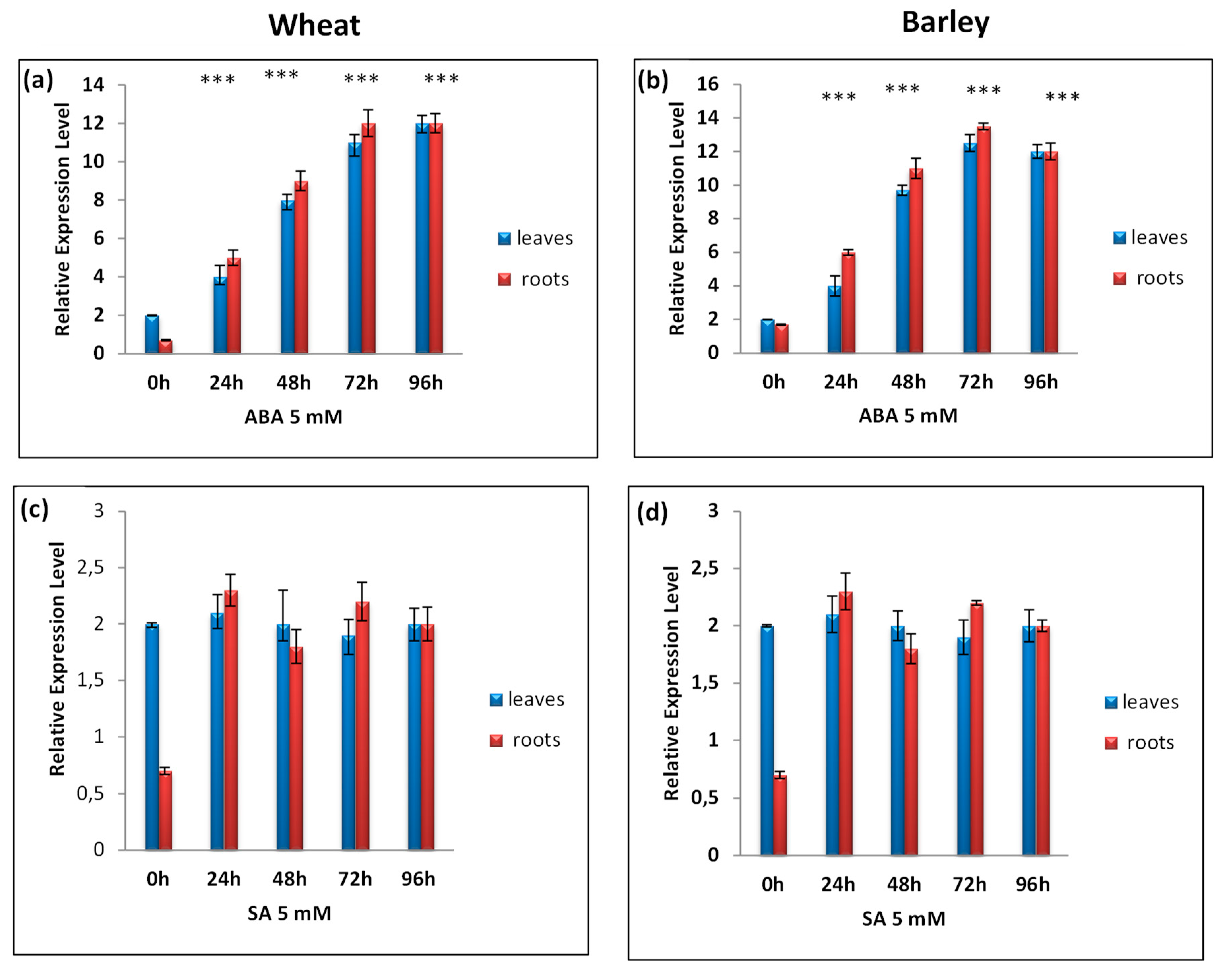
3.4. RT-qPCR Analysis of TtCAT1 and HvCAT1 Gene Expressions
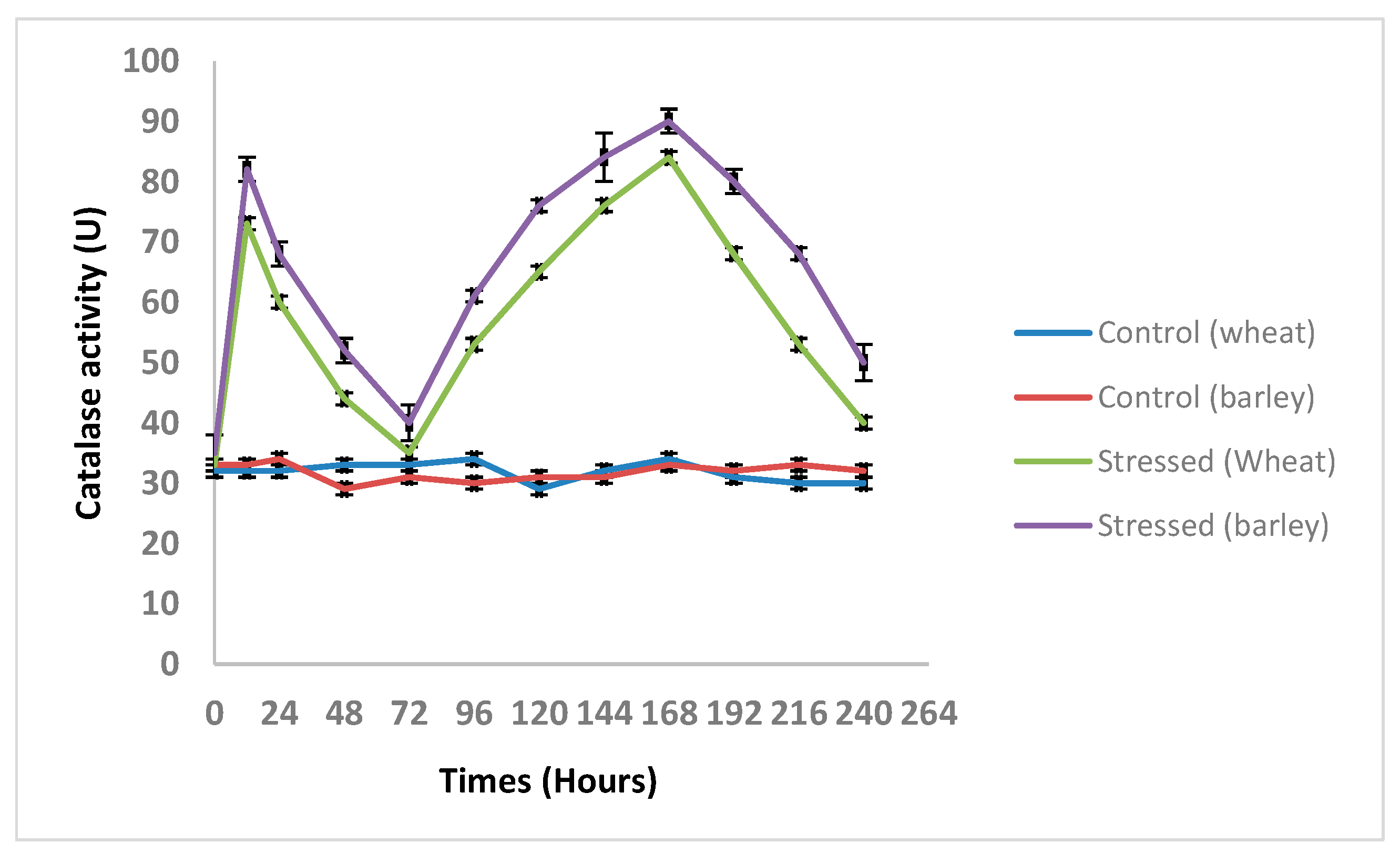
3.5. Changes in the TtCAT and HvCAT Activities under Different Abiotic Stresses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gechev, T.; Petrov, V. Reactive Oxygen Species and Abiotic Stress in Plants. Int. J. Mol. Sci. 2020, 21, 7433. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- He, J.; Xu, L.; Yang, L.; Wang, X. Epigallocatechin gallate is the most effective catechin against antioxidant stress via hydrogen peroxide and radical scavenging activity. Med. Sci. Monit. 2018, 24, 8198. [Google Scholar] [CrossRef]
- Su, Y.; Guo, J.; Ling, H.; Chen, S.; Wang, S.; Xu, L.; Allan, A.C. Isolation of a Novel Peroxisomal Catalase Gene from Sugarcane, Which Is Responsive to Biotic and Abiotic Stresses. PLoS ONE 2014, 9, e84426. [Google Scholar] [CrossRef]
- Feki, K.; Kamoun, Y.; Mahmoud, R.B.; Farhat-Khemakhem, A.; Gargouri, A.; Brini, F. Multiple abiotic stress tolerance of the transformants yeast cells and the transgenic Arabidopsis plants expressing a novel durum wheat catalase. Plant Physiol. Biochem. 2015, 97, 420–431. [Google Scholar] [CrossRef]
- Tounsi, S.; Kamoun, Y.; Feki, K.; Jemli, S.; Saïdi, M.N.; Ziadi, H.; Alcon, C.; Brini, F. Localization and expression analysis of a novel catalase from Triticum monococcum TmCAT1 involved in response to different environmental stresses. Plant Physiol. Biochem. 2019, 139, 366–378. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Gao, C.; Jiang, L.; Guo, D. PPero, a computational model for plant PTS1 type peroxisomal protein prediction. PLoS ONE 2017, 12, e0168912. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, Y.; Chen, D.; Liu, D.; Hu, M.; Jie Dong, J.; Zhang, X.; Song, L.; Shen, F. The Catalase Gene Family in Cotton: Genome-Wide Characterization and Bioinformatics Analysis. Cells 2019, 8, 86. [Google Scholar] [CrossRef]
- Hu, B.; Allina, J.; Bai, J.; Kesar, V.; Odin, J.A. Catalase and estradiol inhibit mitochondrial protein S-glutathionylation. Mol. Cell. Biochem. 2012, 367, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Higo, H.; Higo, K. Differential diurnal expression of rice catalase genes: The 5′-flanking region of CatA is not sufficient for circadian control. Plant Sci. 2000, 151, 39–46. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Yun, L.; Ji, L.; Li, G.; Ji, M.; Shi, Y.; Zheng, X. Catalase (CAT) Gene Family in Wheat (Triticum aestivum L.): Evolution, Expression Pattern and Function Analysis. Int. J. Mol. Sci. 2022, 23, 542. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.; Hussain, M.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-Wide Analysis, Identification, and Expression Pattern in Response to Multiple Hormones and Abiotic Stress Conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef]
- Md, M.R.; Sadia, S.A.; Afsana, H.A.; Ferdousi, B.; Mohammad, A. Response of catalase to drought in barley (Hordeum vulgare L.) seedlings and its purification. Afr. J. Biotechnol. 2020, 19, 478–486. [Google Scholar] [CrossRef]
- Giri, T.K.; Pramanik, K.; Barman, T.K.; Maity, S. Nano-encapsulation of dietary phytoconstituent capsaicin on emulsome: Evaluation of anticancer activity through the measurement of liver oxidative stress in rats. Anti-Cancer Agents Med. Chem. 2017, 17, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Heinlein, C.; Orendi, G.; Zentgraf, U. Senescence-specific regulation of catalases in Arabidopsis thaliana (L.) Heynh. Plant Cell Environ. 2006, 29, 1049–1060. [Google Scholar] [CrossRef]
- Du, Y.Y.; Wang, P.C.; Chen, J.; Song, C.P. Comprehensive functional analysis of the catalase gene family in Arabidopsis thaliana. J. Integr. Plant Biol. 2008, 50, 1318–1326. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Geourjon, C.; Deléage, D. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignements. CABIOS 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Reynolds, C.R.; Islam, S.A.; Sternberg, M.J.E. EzMol: A web server wizard for the rapid visualisation and image production of protein and nucleic acid structures. J. Mol. Biol. 2018, 430, 2244–2248. [Google Scholar] [CrossRef] [PubMed]
- Dimmer, E.C.; Huntley, R.P.; Alam-Faruque, Y.; Sawford, T.; O’Donovan, C.; Martin, M.J.; Bely, B.; Browne, P.; Chan, W.M.; Eberhardt, R.; et al. The UniProt-GO Annotation database in 2011. Nucl. Acids Res. 2012, 40, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.; Pethica, R.; Zhou, Y.; Talbot, C.; Vogel, C.; Madera, M.; Chothia, C.; Gough, J. SUPERFAMILY—Sophisticated comparative genomics, data mining, visualization and phylogeny. Nucl. Acids Res. 2009, 37, D380–D386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiefer, F.; Arnold, K.; Künzli, M.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository and associated resources. Nucl. Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, K.L.; Kim, J.; Truong, K.; Sherman, M.; Yuan, T.; Ikura, M. Calmodulin target database. J. Struct. Funct. Genom. 2000, 1, 8–14. [Google Scholar] [CrossRef]
- Pandit, S.B.; Bhadra, R.; Gowri, V.; Balaji, S.; Anand, B.; Srinivasan, N. SUPFAM: A database of sequence superfamilies of protein domains. BMC Bioinform. 2004, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Kopp, J.; Schwede, T. The SWISS-MODEL Repository of annotated three-dimensional protein structure homology models. Nucl. Acids Res. 2004, 32, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.S.; Cheng, C.W.; Su, W.C.; Chang, K.C.; Huang, S.W.; Hwang, J.K.; Lu, C.H. CELLO2GO: A Web Server for Protein subCELlular LOcalization Prediction with Functional Gene Ontology Annotation. PLoS ONE 2014, 9, e99368. [Google Scholar] [CrossRef] [Green Version]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucl. Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.H.; Wu, L.C.; Lee, T.Y.; Chen, S.P.; Huang, H.D.; Horng, J.T. EuLoc: A web-server for accurately predict protein subcellular localization in eukaryotes by incorporating various features of sequence segmentsinto the general form of Chou’s PseAAC. J. Comput.-Aided Mol. Des. 2013, 27, 91–103. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Ashraf, M.; Ali, Q. Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ. Exp. Bot. 2008, 63, 266–273. [Google Scholar] [CrossRef]
- Hao, L.; Zhou, L.; Xu, X.; Cao, J.; Xi, T. The role of salicylic acid and carrot embryogenic callus extracts in somatic embryogenesis of naked oat (Avena nuda). Plant Cell Tissue Organ Cult. 2006, 85, 109–113. [Google Scholar] [CrossRef]
- Frugoli, J.A.; Zhong, H.H.; Nuccio, M.L.; McCourt, P.; McPeek, M.A.; Thomas, T.L.; McClung, C.R. Catalase is encoded by a multigene family in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1996, 112, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, O.K.; Shchennikova, A.V.; Kochieva, E.Z.; Filyushin, M.A. Pathogenesis-RelatedGenes of PR1, PR2, PR4 and PR5 Families Are Involved in the Response to Fusarium Infection in Garlic (Allium sativum L.). Int. J. Mol. Sci. 2021, 22, 6688. [Google Scholar] [CrossRef] [PubMed]
- AlHudaib, K.A.; Alanazi, N.A.; Ghorbel, M.; El-Ganainy, S.M.; Brini, F. Isolation and Characterization of a Novel Pathogenesis-Related Protein-1 Gene (AvPR-1) with Induced Expression in Oat (Avena sativa L.) during Abiotic and Hormonal Stresses. Plants 2022, 11, 2284. [Google Scholar] [CrossRef]
- Chen, H.J.; Wu, S.D.; Huang, G.J.; Shen, C.Y.; Afiyanti, M.; Li, W.J.; Lin, Y.H. Expression of a cloned sweet potato catalase SPCAT1 alleviates ethephon-mediated leaf senescence and H2O2 elevation. J. Plant Physiol. 2012, 169, 86–97. [Google Scholar] [CrossRef]
- Ghorbel, M.; Feki, K.; Tounsi, S.; Haddaji, N.; Hanin, M.; Brini, F. The Activity of the Durum Wheat (Triticum durum L.) Catalase 1 (TdCAT1) Is Modulated by Calmodulin. Antioxidants 2022, 11, 1483. [Google Scholar] [CrossRef]
- Purev, M.; Kim, Y.J.; Kim, M.K.; Rama Krishna Pulla, R.K.; Yang, D.C. Isolation of a novel catalase (Cat1) gene from Panax ginseng and analysis of the response of this gene to various stresses. Plant Physiol. Biochem. 2010, 48, 451–460. [Google Scholar] [CrossRef]
- Loewen, P.C.; Villanueva, J.; Switala, J.; Donald, L.J.; Ivancich, A. Unprecedented access of phenolic substrates to the heme active site of a catalase: Substrate binding and peroxidase-like reactivity of Bacillus pumilus catalase monitored by X-ray crystallography and EPR spectroscopy. Proteins Struct. Funct. Bioinform. 2015, 83, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsova, B.; Schulze, W.X. Current status of the plant phosphorylation site database PhosPhAt and its use as a resource for molecular plant physiology. Front. Plant Sci. 2012, 3, 132. [Google Scholar] [CrossRef] [Green Version]
- Ghorbel, M.; Feki, K.; Tounsi, S.; Bouali, N.; Besbes, M.; Brini, F. The Putative Auto-Inhibitory Domain of Durum Wheat Catalase (TdCAT1) Positively Regulates Bacteria Cells in Response to Different Stress Conditions. Antioxidants 2022, 11, 1820. [Google Scholar] [CrossRef]
- Willekens, H.; Langebartels, C.; Tire, C.; Van Montagu, M.; Inze, D.; Van Camp, W. Differential expression of catalase genes in Nicotiana plumbaginifolia (L.). Proc. Natl. Acad. Sci. USA 1994, 91, 10450–10454. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, J.; Lee, Y.H.; Song, S.I. Rice CatA, CatB, and CatC are involved in environmental stress response, root growth, and photorespiration, respectively. J. Plant Biol. 2014, 57, 375–382. [Google Scholar] [CrossRef]
- Bouthiba, A.; Debaeke, P.; Hamoudi, S.A. Varietal differences in the response of durum wheat (Triticum turgidum L. var. durum) to irrigation strategies in a semi-arid region of Algeria. Irrig. Sci. 2008, 26, 239–251. [Google Scholar]
- Tyagi, S.; Singh, K.; Upadhyay, S.K. Molecular characterization revealed the role of catalases under abiotic and arsenic stress in bread wheat (Triticum aestivum L.). J. Hazard. Mater. 2021, 403, 123585. [Google Scholar] [CrossRef]
- Sooch, B.S.; Kauldhar, B.S.; Puri, M. Recent insights into microbial catalases: Isolation, production and purification. Biotechnol. Adv. 2014, 32, 1429–1447. [Google Scholar] [CrossRef]
- Oshima, Y.; Kamigaki, A.; Nakamori, C.; Mano, S.; Hayashi, M.; Nishimura, M.; Esaka, M. Plant catalase is imported into peroxisomes by Pex5p but is distinct from typical PTS1 import. Plant. Cell Physiol. 2008, 49, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, Y.; Suekawa, M.; Endo, S.; Fukami, Y.; Mano, S.; Nishimura, M.; Esaka, M. Effect of mutation of C-terminal and heme binding region of Arabidopsis catalaseon the import to peroxisomes. Biosci. Biotechnol. Biochem. 2018, 83, 322–325. [Google Scholar] [CrossRef]
- Kaur, N.; Hu, J. Defining the plant peroxisomal proteome: From Arabidopsis to rice. Front. Plant Sci. 2011, 2, 103. [Google Scholar] [CrossRef] [Green Version]
- Seth, D.; Hess, D.T.; Hausladen, A.; Wang, L.; Wang, Y.J.; Stamler, J.S. A multiplex enzymatic machinery for cellular protein S-nitrosylation. Mol. Cell 2018, 69, 451–464.e6. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wu, R.; Feng, J.; Feng, T.; Wang, C.; Hu, J.; Zhan, N.; Li, Y.; Ma, X.; Ren, B.; et al. Transnitrosylation mediated by the non-canonical catalase ROG1 regulates nitric oxide signaling in plants. Dev. Cell 2020, 53, 444–457.e5. [Google Scholar] [CrossRef] [PubMed]
- Pande, A.; Mun, B.G.; Rahim, W.; Khan, M.; Lee, D.S.; Lee, G.M.; Al Azzawi, T.N.I.; Hussain, A.; Kim, C.K.; Yun, B.W. Phytohormonal Regulation Through Protein S-Nitrosylation Under Stress. Front. Plant Sci. 2022, 13, 865542. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ruiz, M.; González-Gordo, S.; Cañas, A.; Campos, M.J.; Paradela, A.; Corpas, F.J.; Palma, J.M. Sweet Pepper (Capsicum annuum L.) Fruits Contain an Atypical Peroxisomal Catalase That Is Modulated by Reactive Oxygen and Nitrogen Species. Antioxydants 2019, 8, 374. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Total Number of Negatively Charged Residues (Asp + Glu): | Total Number of Positively Charged Residues (Arg + Lys): | Grand Average of Hydropathicity (GRAVY): | MW | Number of aa |
|---|---|---|---|---|---|
| TtCAT1 | 63 | 58 | −0.595 | 56807.99 | 492 |
| HvCAT1 | 63 | 58 | −0.591 | 56567.65 | 492 |
| TdCat1 | 65 | 58 | −0.600 | 56768.91 | 492 |
| TaCat1 | 63 | 58 | −0.595 | 56807.99 | 492 |
| AetCAT1 | 63 | 58 | −0.598 | 56723.87 | 492 |
| TmCAT1 | 62 | 58 | −0.590 | 56661.85 | 492 |
| BdCat1 | 61 | 58 | −0.608 | 56746.93 | 492 |
| AtCat1 | 62 | 58 | −0.574 | 56917.27 | 492 |
| TdcCAT1 | 63 | 58 | −0.599 | 56767.93 | 492 |
| PgCAT1 | 61 | 57 | −0.575 | 56766.98 | 492 |
| SaCAT1 | 61 | 59 | −0.548 | 56921.49 | 492 |
| GhCAT1 | 61 | 56 | −0.537 | 56855.20 | 492 |
| LjCAT1 | 61 | 57 | −0.547 | 56893.19 | 492 |
| EsCAT1 | 59 | 58 | −0.522 | 56689.18 | 492 |
| GrCAT1 | 61 | 57 | −0.538 | 56817.20 | 492 |
| AiCAT1 | 61 | 58 | −0.562 | 56955.22 | 492 |
| Protein | α-Helices | β-Turns | Random Coils | Extended Strands |
|---|---|---|---|---|
| TtCAT1 | 140 | 27 | 242 | 83 |
| HvCAT1 | 136 | 26 | 258 | 72 |
| TdCat1 | 135 | 27 | 256 | 74 |
| TaCat1 | 140 | 27 | 242 | 83 |
| AetCAT1 | 127 | 30 | 258 | 77 |
| TmCAT1 | 137 | 30 | 249 | 76 |
| BdCat1 | 134 | 29 | 257 | 72 |
| AtCat1 | 126 | 31 | 260 | 75 |
| TdcCAT1 | 131 | 28 | 258 | 75 |
| PgCAT1 | 138 | 29 | 250 | 75 |
| SaCAT1 | 135 | 27 | 253 | 77 |
| GhCAT1 | 139 | 29 | 249 | 75 |
| LjCAT1 | 130 | 25 | 262 | 75 |
| EsCAT1 | 137 | 28 | 255 | 72 |
| GrCAT1 | 141 | 29 | 248 | 74 |
| AiCAT1 | 134 | 32 | 247 | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghorbel, M.; Besbes, M.; Haddaji, N.; Bouali, N.; Brini, F. Identification and Expression Profiling of Two Saudi Arabia Catalase Genes from Wheat and Barley in Response to Abiotic and Hormonal Stresses. Antioxidants 2022, 11, 2208. https://doi.org/10.3390/antiox11112208
Ghorbel M, Besbes M, Haddaji N, Bouali N, Brini F. Identification and Expression Profiling of Two Saudi Arabia Catalase Genes from Wheat and Barley in Response to Abiotic and Hormonal Stresses. Antioxidants. 2022; 11(11):2208. https://doi.org/10.3390/antiox11112208
Chicago/Turabian StyleGhorbel, Mouna, Malek Besbes, Najla Haddaji, Nouha Bouali, and Faiçal Brini. 2022. "Identification and Expression Profiling of Two Saudi Arabia Catalase Genes from Wheat and Barley in Response to Abiotic and Hormonal Stresses" Antioxidants 11, no. 11: 2208. https://doi.org/10.3390/antiox11112208