
Environmental Enrichment Engages Vesicular Zinc Signaling to Enhance Hippocampal Neurogenesis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Housing Manipulation
2.3. BrdU Procedure
2.4. Tissue Preparation and Labeling
2.5. Microscopy and Cell Quantification
2.6. Behavioural Analysis
2.6.1. Spatial Object Recognition
2.6.2. Morris Water Task
2.7. BDNF Analysis and Zinc Incubated Slices
2.8. Statistical Analysis
3. Results
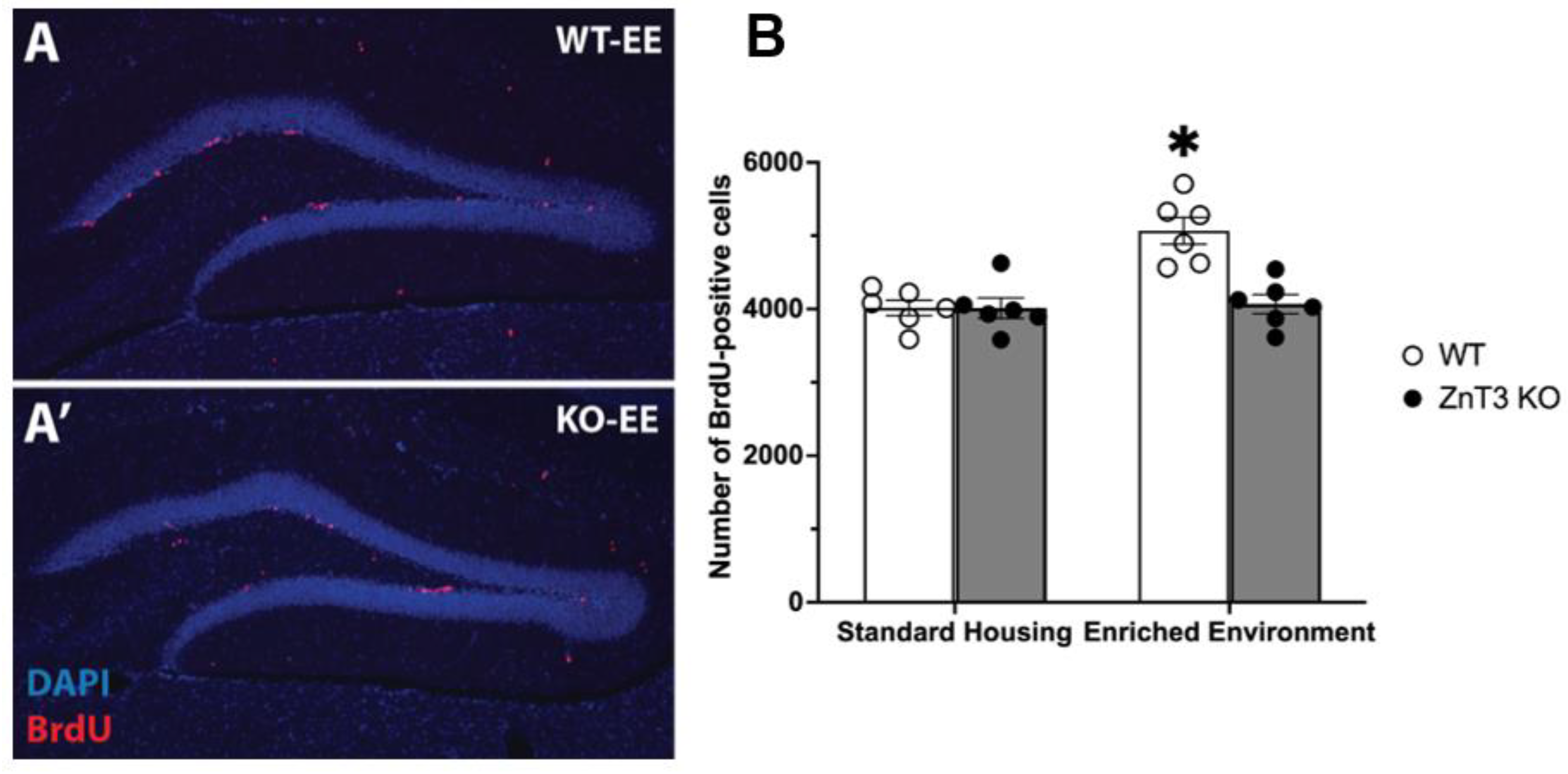
3.1. Knockout of ZnT3 Prevents Enriched Environment-Induced Proliferation without Affecting Baseline Proliferation
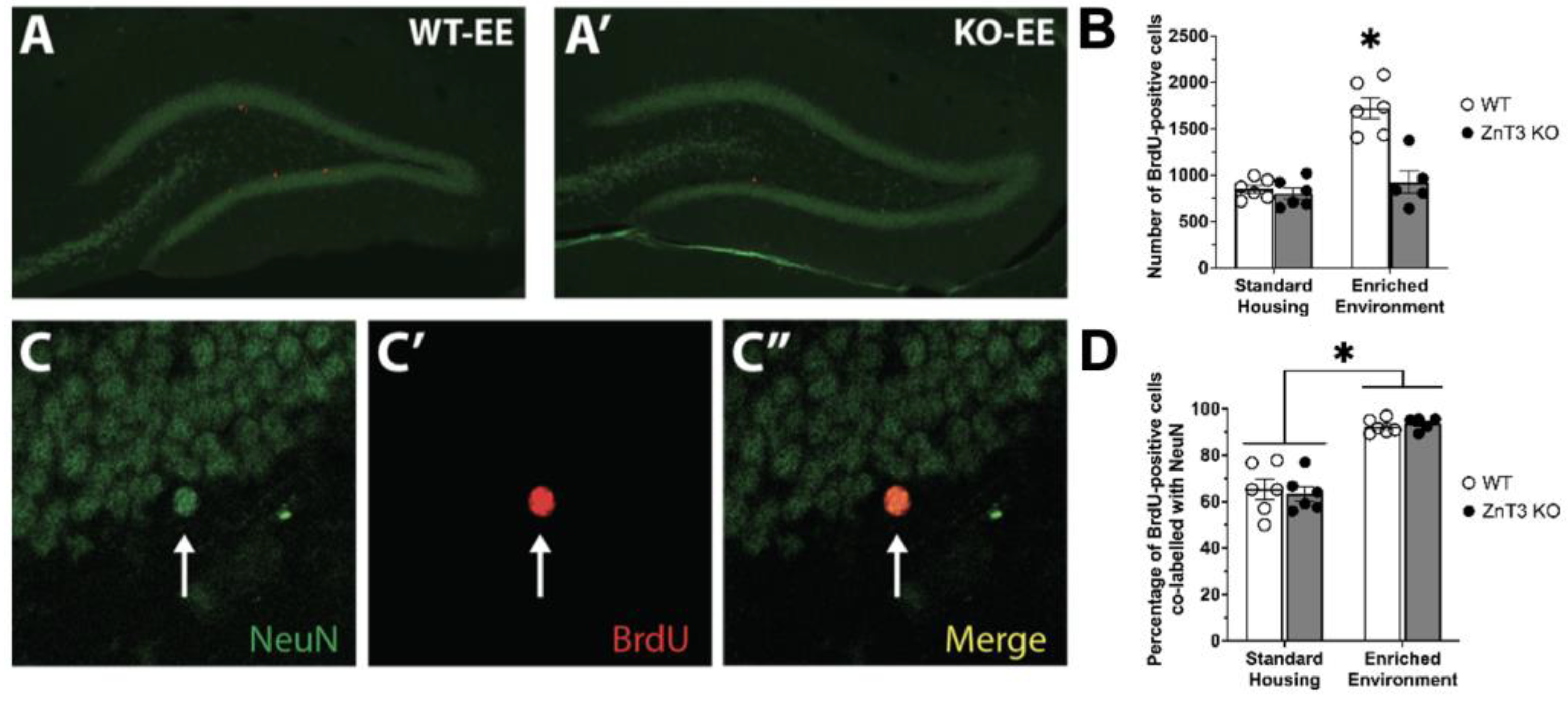
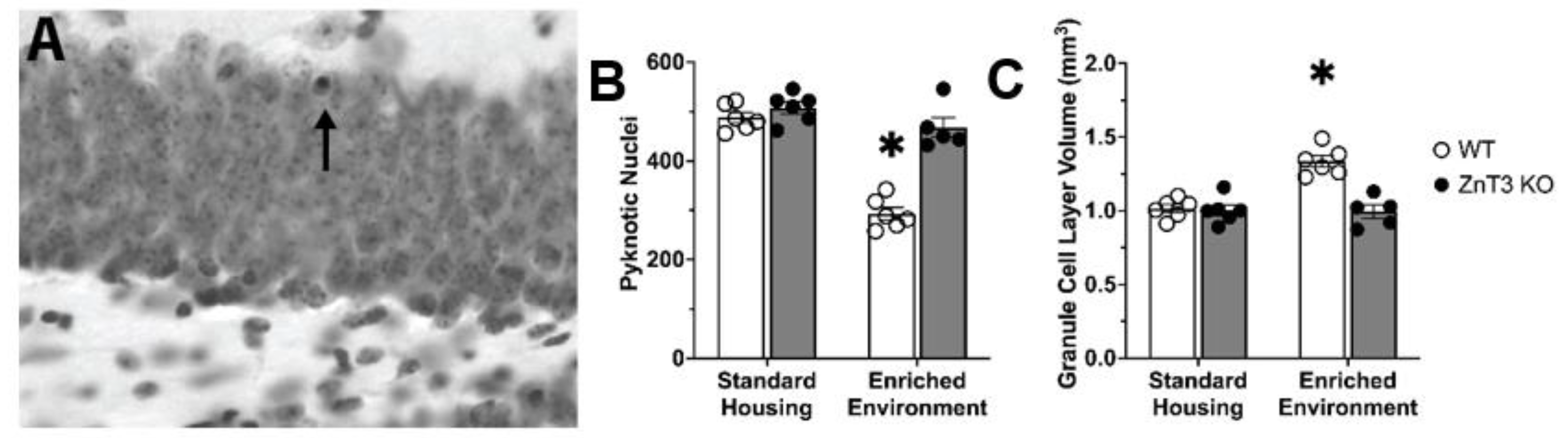
3.2. Vesicular Zinc Is Necessary for Enriched-Environment-Induced Survival of New Neurons
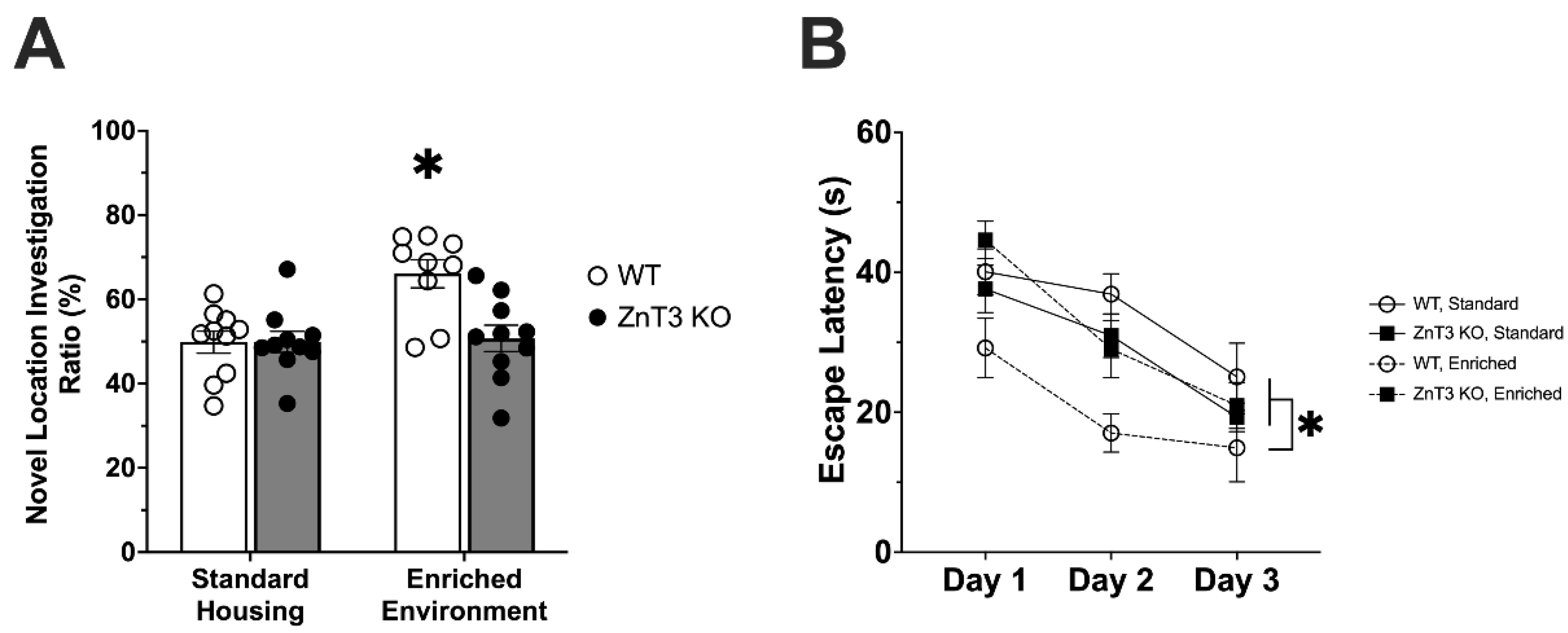
3.3. Mice Lacking Vesicular Zinc Show No Improvement in Spatial Memory following Environmental Enrichment
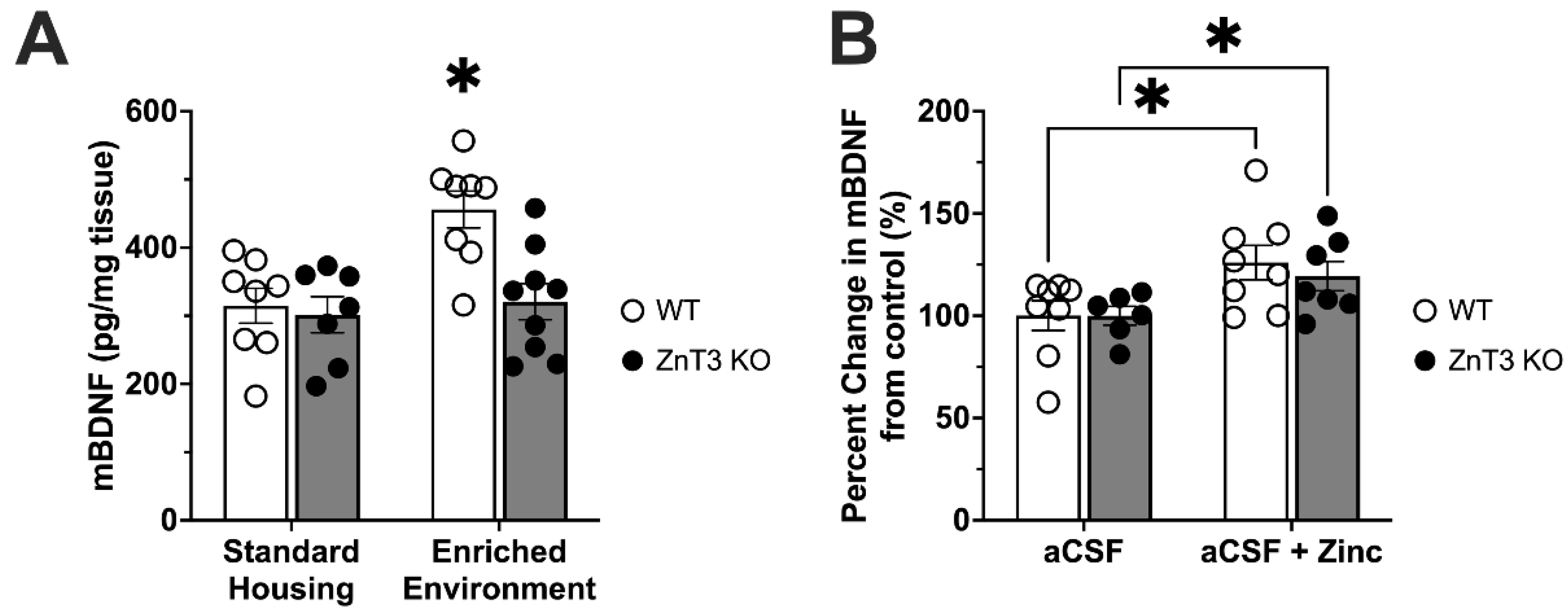
3.4. Zinc, Environmental Enrichment, and BDNF
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Beaulieu, C.; Dyck, R.; Cynader, M. Enrichment of Glutamate in Zinc-Containing Terminals of the Cat Visual Cortex. Neuroreport 1992, 3, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.S.; Dyck, R.H. Zinc and Cortical Plasticity. Brain Res. Rev. 2009, 59, 347–373. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Cole, T.B.; Quaife, C.J.; Findley, S.D. ZnT-3, a Putative Transporter of Zinc into Synaptic Vesicles. Proc. Natl. Acad. Sci. USA 1996, 93, 14934–14939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Noebels, J.L. Visualization of Transmitter Release with Zinc Fluorescence Detection at the Mouse Hippocampal Mossy Fibre Synapse. J. Physiol. 2005, 566, 747–758. [Google Scholar] [CrossRef] [PubMed]
- McAllister, B.B.; Dyck, R.H. Zinc Transporter 3 (ZnT3) and Vesicular Zinc in Central Nervous System Function. Neurosci. Biobehav. Rev. 2017, 80, 329–350. [Google Scholar] [CrossRef] [PubMed]
- Christine, C.W.; Choi, D.W. Effect of Zinc on NMDA Receptor-Mediated Channel Currents in Cortical Neurons. J. Neurosci. 1990, 10, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.Z.; Pan, E.; Xiong, Z.-Q.; McNamara, J.O. Zinc-Mediated Transactivation of TrkB Potentiates the Hippocampal Mossy Fiber-CA3 Pyramid Synapse. Neuron 2008, 57, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Slomianka, L. Neurons of Origin of Zinc-Containing Pathways and the Distribution of Zinc-Containing Boutons in the Hippocampal Region of the Rat. Neuroscience 1992, 48, 325–352. [Google Scholar] [CrossRef]
- Cole, T.B.; Wenzel, H.J.; Kafer, K.E.; Schwartzkroin, P.A.; Palmiter, R.D. Elimination of Zinc from Synaptic Vesicles in the Intact Mouse Brain by Disruption of the ZnT3 Gene. Proc. Natl. Acad. Sci. USA 1999, 96, 1716–1721. [Google Scholar] [CrossRef] [Green Version]
- Lopantsev, V.; Wenzel, H.J.; Cole, T.B.; Palmiter, R.D.; Schwartzkroin, P.A. Lack of Vesicular Zinc in Mossy Fibers Does Not Affect Synaptic Excitability of CA3 Pyramidal Cells in Zinc Transporter 3 Knockout Mice. Neuroscience 2003, 116, 237–248. [Google Scholar] [CrossRef]
- Cole, T.B.; Martyanova, A.; Palmiter, R.D. Removing Zinc from Synaptic Vesicles Does Not Impair Spatial Learning, Memory, or Sensorimotor Functions in the Mouse. Brain Res. 2001, 891, 253–265. [Google Scholar] [CrossRef]
- Adlard, P.A.; Parncutt, J.M.; Finkelstein, D.I.; Bush, A.I. Cognitive Loss in Zinc Transporter-3 Knock-out Mice: A Phenocopy for the Synaptic and Memory Deficits of Alzheimer’s Disease? J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 1631–1636. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.J.; Park, M.-H.; Choi, S.-Y.; Koh, J.-Y. Activation of the Trk Signaling Pathway by Extracellular Zinc. Role of Metalloproteinases. J. Biol. Chem. 2005, 280, 11995–12001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, E.G.; An, J.J.; Orefice, L.L.; Baydyuk, M.; Liao, G.-Y.; Zheng, K.; Lu, B.; Xu, B. BDNF Promotes Differentiation and Maturation of Adult-Born Neurons through GABAergic Transmission. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 14318–14330. [Google Scholar] [CrossRef] [Green Version]
- Barnabé-Heider, F.; Miller, F.D. Endogenously Produced Neurotrophins Regulate Survival and Differentiation of Cortical Progenitors via Distinct Signaling Pathways. J. Neurosci. 2003, 23, 5149–5160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levenson, C.W.; Morris, D. Zinc and Neurogenesis: Making New Neurons from Development to Adulthood. Adv. Nutr. 2011, 2, 96–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Aimone, J.B.; Gage, F.H. New Neurons and New Memories: How Does Adult Hippocampal Neurogenesis Affect Learning and Memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Kuhn, H.G.; Gage, F.H. More Hippocampal Neurons in Adult Mice Living in an Enriched Environment. Nature 1997, 386, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched Environment Increases Neurogenesis in the Adult Rat Dentate Gyrus and Improves Spatial Memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- Van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running Enhances Neurogenesis, Learning, and Long-Term Potentiation in Mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, D.; Lawlor, P.A.; Leone, P.; Dragunow, M.; During, M.J. Environmental Enrichment Inhibits Spontaneous Apoptosis, Prevents Seizures and Is Neuroprotective. Nat. Med. 1999, 5, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-Derived Neurotrophic Factor (BDNF) Is Required for the Enhancement of Hippocampal Neurogenesis Following Environmental Enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef] [PubMed]
- Kee, N.; Teixeira, C.M.; Wang, A.H.; Frankland, P.W. Preferential Incorporation of Adult-Generated Granule Cells into Spatial Memory Networks in the Dentate Gyrus. Nat. Neurosci. 2007, 10, 355–362. [Google Scholar] [CrossRef]
- Creer, D.J.; Romberg, C.; Saksida, L.M.; van Praag, H.; Bussey, T.J. Running Enhances Spatial Pattern Separation in Mice. Proc. Natl. Acad. Sci. USA 2010, 107, 2367–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O’Carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing Adult Hippocampal Neurogenesis Is Sufficient to Improve Pattern Separation. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shors, T.J.; Townsend, D.A.; Zhao, M.; Kozorovitskiy, Y.; Gould, E. Neurogenesis May Relate to Some but Not All Types of Hippocampal-Dependent Learning. Hippocampus 2002, 12, 578–584. [Google Scholar] [CrossRef]
- Madsen, T.M.; Kristjansen, P.E.G.; Bolwig, T.G.; Wörtwein, G. Arrested Neuronal Proliferation and Impaired Hippocampal Function Following Fractionated Brain Irradiation in the Adult Rat. Neuroscience 2003, 119, 635–642. [Google Scholar] [CrossRef]
- Snyder, J.S.; Hong, N.S.; McDonald, R.J.; Wojtowicz, J.M. A Role for Adult Neurogenesis in Spatial Long-Term Memory. Neuroscience 2005, 130, 843–852. [Google Scholar] [CrossRef]
- Winocur, G.; Wojtowicz, J.M.; Sekeres, M.; Snyder, J.S.; Wang, S. Inhibition of Neurogenesis Interferes with Hippocampus-Dependent Memory Function. Hippocampus 2006, 16, 296–304. [Google Scholar] [CrossRef]
- Saxe, M.D.; Battaglia, F.; Wang, J.-W.; Malleret, G.; David, D.J.; Monckton, J.E.; Garcia, A.D.R.; Sofroniew, M.V.; Kandel, E.R.; Santarelli, L.; et al. Ablation of Hippocampal Neurogenesis Impairs Contextual Fear Conditioning and Synaptic Plasticity in the Dentate Gyrus. Proc. Natl. Acad. Sci. USA 2006, 103, 17501–17506. [Google Scholar] [CrossRef] [Green Version]
- Dupret, D.; Revest, J.-M.; Koehl, M.; Ichas, F.; De Giorgi, F.; Costet, P.; Abrous, D.N.; Piazza, P.V. Spatial Relational Memory Requires Hippocampal Adult Neurogenesis. PLoS ONE 2008, 3, e1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, A.K.; Eadie, B.D.; Ernst, C.; Christie, B.R. Environmental Enrichment and Voluntary Exercise Massively Increase Neurogenesis in the Adult Hippocampus via Dissociable Pathways. Hippocampus 2006, 16, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, C.D.; Harburg, G.C.; Eisch, A.J. Determination of Key Aspects of Precursor Cell Proliferation, Cell Cycle Length and Kinetics in the Adult Mouse Subgranular Zone. Neuroscience 2007, 146, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taupin, P. BrdU Immunohistochemistry for Studying Adult Neurogenesis: Paradigms, Pitfalls, Limitations, and Validation. Brain Res. Rev. 2007, 53, 198–214. [Google Scholar] [CrossRef]
- Galea, L.A.M.; Uban, K.A.; Epp, J.R.; Brummelte, S.; Barha, C.K.; Wilson, W.L.; Lieblich, S.E.; Pawluski, J.L. Endocrine Regulation of Cognition and Neuroplasticity: Our Pursuit to Unveil the Complex Interaction between Hormones, the Brain, and Behaviour. Can. J. Exp. Psychol. Rev. Can. Psychol. Exp. 2008, 62, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Sliwowska, J.H.; Barker, J.M.; Barha, C.K.; Lan, N.; Weinberg, J.; Galea, L.A.M. Stress-Induced Suppression of Hippocampal Neurogenesis in Adult Male Rats Is Altered by Prenatal Ethanol Exposure. Stress 2010, 13, 302–314. [Google Scholar] [CrossRef] [Green Version]
- Kronenberg, G.; Bick-Sander, A.; Bunk, E.; Wolf, C.; Ehninger, D.; Kempermann, G. Physical Exercise Prevents Age-Related Decline in Precursor Cell Activity in the Mouse Dentate Gyrus. Neurobiol. Aging 2006, 27, 1505–1513. [Google Scholar] [CrossRef]
- Rakai, B.D.; Chrusch, M.J.; Spanswick, S.C.; Dyck, R.H.; Antle, M.C. Survival of Adult Generated Hippocampal Neurons Is Altered in Circadian Arrhythmic Mice. PLoS ONE 2014, 9, e99527. [Google Scholar] [CrossRef]
- Spanswick, S.C. Adrenalectomy-Induced Neuronal Degeneration: Development of a Novel Animal Model of Cognitive Dysfuntion and Neurogenic Treatment Strategies. Ph.D. Thesis, Department of Neuroscience, University of Lethbridge, Lethbridge, AB, Canada, 2010. [Google Scholar]
- Ferrer, I.; Soriano, E.; Del Rio, J.A.; Alcántara, S.; Auladell, C. Cell Death and Removal in the Cerebral Cortex during Development. Prog. Neurobiol. 1992, 39, 1–43. [Google Scholar] [CrossRef]
- Nuñez, J.L.; McCarthy, M.M. Estradiol Exacerbates Hippocampal Damage in a Model of Preterm Infant Brain Injury. Endocrinology 2003, 144, 2350–2359. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, H.J.G.; Bendtsen, T.F.; Korbo, L.; Marcussen, N.; Møller, A.; Nielsen, K.; Nyengaard, J.R.; Pakkenberg, B.; Sørensen, F.B.; Vesterby, A.; et al. Some New, Simple and Efficient Stereological Methods and Their Use in Pathological Research and Diagnosis. APMIS 1988, 96, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanswick, S.C.; Dyck, R.H. Object/Context Specific Memory Deficits Following Medial Frontal Cortex Damage in Mice. PLoS ONE 2012, 7, e43698. [Google Scholar] [CrossRef] [Green Version]
- Stone, S.S.D.; Teixeira, C.M.; Devito, L.M.; Zaslavsky, K.; Josselyn, S.A.; Lozano, A.M.; Frankland, P.W. Stimulation of Entorhinal Cortex Promotes Adult Neurogenesis and Facilitates Spatial Memory. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 13469–13484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polacchini, A.; Metelli, G.; Francavilla, R.; Baj, G.; Florean, M.; Mascaretti, L.G.; Tongiorgi, E. A Method for Reproducible Measurements of Serum BDNF: Comparison of the Performance of Six Commercial Assays. Sci. Rep. 2015, 5, 17989. [Google Scholar] [CrossRef] [Green Version]
- Valero, J.; España, J.; Parra-Damas, A.; Martín, E.; Rodríguez-Álvarez, J.; Saura, C.A. Short-Term Environmental Enrichment Rescues Adult Neurogenesis and Memory Deficits in APPSw,Ind Transgenic Mice. PLoS ONE 2011, 6, e16832. [Google Scholar] [CrossRef] [Green Version]
- Corniola, R.S.; Tassabehji, N.M.; Hare, J.; Sharma, G.; Levenson, C.W. Zinc Deficiency Impairs Neuronal Precursor Cell Proliferation and Induces Apoptosis via P53-Mediated Mechanisms. Brain Res. 2008, 1237, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.W.; Won, S.J.; Hamby, A.M.; Yoo, B.H.; Fan, Y.; Sheline, C.T.; Tamano, H.; Takeda, A.; Liu, J. Decreased Brain Zinc Availability Reduces Hippocampal Neurogenesis in Mice and Rats. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2009, 29, 1579–1588. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.E.; Dyck, R.H. Rapid, Experience-Dependent Changes in Levels of Synaptic Zinc in Primary Somatosensory Cortex of the Adult Mouse. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 2617–2625. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.E.; Dyck, R.H. Modulation of Synaptic Zinc in Barrel Cortex by Whisker Stimulation. Neuroscience 2005, 134, 355–359. [Google Scholar] [CrossRef]
- Czupryn, A.; Skangiel-Kramska, J. Differential Response of Synaptic Zinc Levels to Sensory Deprivation in the Barrel Cortex of Young and Adult Mice. Exp. Brain Res. 2001, 141, 567–572. [Google Scholar] [CrossRef]
- Nakashima, A.S.; Dyck, R.H. Enhanced Plasticity in Zincergic, Cortical Circuits after Exposure to Enriched Environments. J. Neurosci. 2008, 28, 13995–13999. [Google Scholar] [CrossRef] [Green Version]
- Rink, L. Zinc in Human Health; IOS Press: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Choi, S.H.; Li, Y.; Parada, L.F.; Sisodia, S.S. Regulation of Hippocampal Progenitor Cell Survival, Proliferation and Dendritic Development by BDNF. Mol. Neurodegener. 2009, 4, 52. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.-Y.; Sun, E.-S.; An, J.H.; Im, H.; Lee, S.-H.; Lee, J.-Y.; Han, P.-L.; Koh, J.-Y.; Kim, Y.-H. Zinc-Triggered Induction of Tissue Plasminogen Activator by Brain-Derived Neurotrophic Factor and Metalloproteinases. J. Neurochem. 2011, 118, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Z.; McNamara, J.O. Mutual Regulation of Src Family Kinases and the Neurotrophin Receptor TrkB. J. Biol. Chem. 2010, 285, 8207–8217. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Siao, C.-J.; Nagappan, G.; Marinic, T.; Jing, D.; McGrath, K.; Chen, Z.-Y.; Mark, W.; Tessarollo, L.; Lee, F.S.; et al. Neuronal Release of ProBDNF. Nat. Neurosci. 2009, 12, 113–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Duan, J.; Wang, X.; Zhong, X.; Hu, Z.; Huang, F.; Wang, H.; Zhang, J.; Li, F.; Zhang, J.; et al. Early Enriched Environment Induces an Increased Conversion of ProBDNF to BDNF in the Adult Rat’s Hippocampus. Behav. Brain Res. 2014, 265, 76–83. [Google Scholar] [CrossRef]
- Obiang, P.; Maubert, E.; Bardou, I.; Nicole, O.; Launay, S.; Bezin, L.; Vivien, D.; Agin, V. Enriched Housing Reverses Age-Associated Impairment of Cognitive Functions and TPA-Dependent Maturation of BDNF. Neurobiol. Learn. Mem. 2011, 96, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Travaglia, A.; La Mendola, D.; Magrì, A.; Pietropaolo, A.; Nicoletti, V.G.; Grasso, G.; Malgieri, G.; Fattorusso, R.; Isernia, C.; Rizzarelli, E. Zinc(II) Interactions with Brain-Derived Neurotrophic Factor N-Terminal Peptide Fragments: Inorganic Features and Biological Perspectives. Inorg. Chem. 2013, 52, 11075–11083. [Google Scholar] [CrossRef]
- Tao, X.; Finkbeiner, S.; Arnold, D.B.; Shaywitz, A.J.; Greenberg, M.E. Ca2+ Influx Regulates BDNF Transcription by a CREB Family Transcription Factor-Dependent Mechanism. Neuron 1998, 20, 709–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.Y.; Hong, D.K.; Jeong, J.H.; Lee, B.E.; Koh, J.-Y.; Suh, S.W. Zinc Transporter 3 Modulates Cell Proliferation and Neuronal Differentiation in the Adult Hippocampus. Stem Cells Dayt. Ohio 2020, 38, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Sindreu, C.; Palmiter, R.D.; Storm, D.R. Zinc Transporter ZnT-3 Regulates Presynaptic Erk1/2 Signaling and Hippocampus-Dependent Memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3366–3370. [Google Scholar] [CrossRef] [Green Version]
- Merritt, J.; Rhodes, J.S. Mouse Genetic Differences in Voluntary Wheel Running, Adult Hippocampal Neurogenesis and Learning on the Multi-Strain-Adapted plus Water Maze. Behav. Brain Res. 2015, 280, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Kobilo, T.; Liu, Q.-R.; Gandhi, K.; Mughal, M.; Shaham, Y.; van Praag, H. Running Is the Neurogenic and Neurotrophic Stimulus in Environmental Enrichment. Learn. Mem. 2011, 18, 605–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanselow, M.S.; Dong, H.-W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Valente, T.; Auladell, C. Developmental Expression of ZnT3 in Mouse Brain: Correlation between the Vesicular Zinc Transporter Protein and Chelatable Vesicular Zinc (CVZ) Cells. Glial and Neuronal CVZ Cells Interact. Mol. Cell. Neurosci. 2002, 21, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Frederickson, C.J.; Giblin, L.J.; Balaji, R.V.; Masalha, R.; Frederickson, C.J.; Zeng, Y.; Lopez, E.V.; Koh, J.-Y.; Chorin, U.; Besser, L.; et al. Synaptic Release of Zinc from Brain Slices: Factors Governing Release, Imaging, and Accurate Calculation of Concentration. J. Neurosci. Methods 2006, 154, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Frazzini, V.; Granzotto, A.; Bomba, M.; Noemi, M.; Vanessa, C.; d’Aurora, M.; Punzi, M.; Iorio, M.; Mosca, A.; Delli Pizzi, S.; et al. The Pharmacological Perturbation of Brain Zinc Impairs BDNF-Related Signaling and the Cognitive Performances of Young Mice. Sci. Rep. 2018, 8, 9768. [Google Scholar] [CrossRef] [Green Version]
- Frederickson, R.E.; Frederickson, C.J.; Danscher, G. In Situ Binding of Bouton Zinc Reversibly Disrupts Performance on a Spatial Memory Task. Behav. Brain Res. 1990, 38, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Daumas, S.; Halley, H.; Lassalle, J.-M. Disruption of Hippocampal CA3 Network: Effects on Episodic-like Memory Processing in C57BL/6J Mice. Eur. J. Neurosci. 2004, 20, 597–600. [Google Scholar] [CrossRef]
- Lassalle, J.M.; Bataille, T.; Halley, H. Reversible Inactivation of the Hippocampal Mossy Fiber Synapses in Mice Impairs Spatial Learning, but Neither Consolidation nor Memory Retrieval, in the Morris Navigation Task. Neurobiol. Learn. Mem. 2000, 73, 243–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, A.R. Evidence for Chelatable Zinc in the Extracellular Space of the Hippocampus, but Little Evidence for Synaptic Release of Zn. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 6847–6855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoletti, P.; Vergnano, A.M.; Barbour, B.; Casado, M. Zinc at Glutamatergic Synapses. Neuroscience 2009, 158, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Pan, E.; Zhang, X.; Huang, Z.; Krezel, A.; Zhao, M.; Tinberg, C.E.; Lippard, S.J.; McNamara, J.O. Vesicular Zinc Promotes Presynaptic and Inhibits Postsynaptic Long-Term Potentiation of Mossy Fiber-CA3 Synapse. Neuron 2011, 71, 1116–1126. [Google Scholar] [CrossRef] [Green Version]
- Radford, R.J.; Lippard, S.J. Chelators for Investigating Zinc Metalloneurochemistry. Curr. Opin. Chem. Biol. 2013, 17, 129–136. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrusch, M.J.; Fu, S.; Spanswick, S.C.; Vecchiarelli, H.A.; Patel, P.P.; Hill, M.N.; Dyck, R.H. Environmental Enrichment Engages Vesicular Zinc Signaling to Enhance Hippocampal Neurogenesis. Cells 2023, 12, 883. https://doi.org/10.3390/cells12060883
Chrusch MJ, Fu S, Spanswick SC, Vecchiarelli HA, Patel PP, Hill MN, Dyck RH. Environmental Enrichment Engages Vesicular Zinc Signaling to Enhance Hippocampal Neurogenesis. Cells. 2023; 12(6):883. https://doi.org/10.3390/cells12060883
Chicago/Turabian StyleChrusch, Michael J., Selena Fu, Simon C. Spanswick, Haley A. Vecchiarelli, Payal P. Patel, Matthew N. Hill, and Richard H. Dyck. 2023. "Environmental Enrichment Engages Vesicular Zinc Signaling to Enhance Hippocampal Neurogenesis" Cells 12, no. 6: 883. https://doi.org/10.3390/cells12060883