

Impact of Elevated Brain IL-6 in Transgenic Mice on the Behavioral and Neurochemical Consequences of Chronic Alcohol Exposure
Abstract
:1. Introduction
2. Materials and Methods
2.1. Transgenic Mice
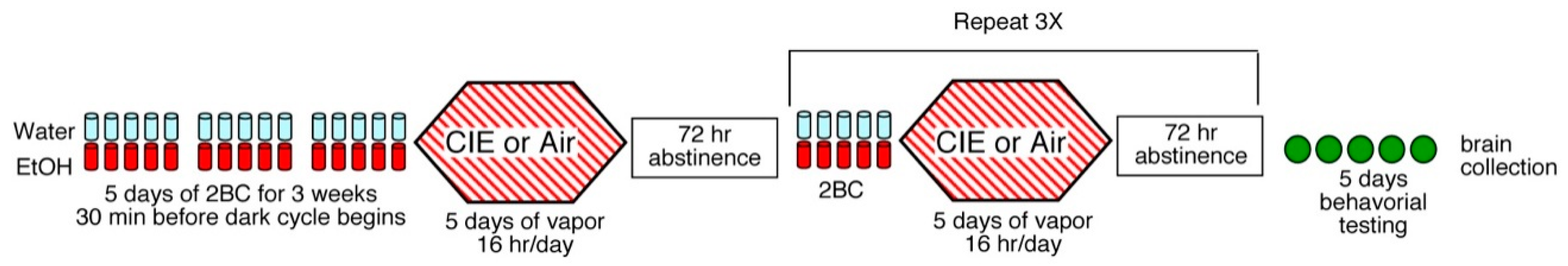
2.2. Two-Bottle Choice Drinking-Chronic Intermittent Alcohol Exposure Protocol (2BC-CIE)
2.3. Behavior Tests for Negative Affect/Emotionality
2.3.1. Light/Dark Transfer Test
2.3.2. Digging Test
2.3.3. Open Field Test
2.3.4. Forced Swim Test
2.3.5. Tail Flick Test
2.4. Protein Assay
2.4.1. Preparation of Protein Samples
2.4.2. Western Blot
2.4.3. Antibodies
2.5. IL-6 and TNF-Alpha Levels
2.6. Statistics
3. Results
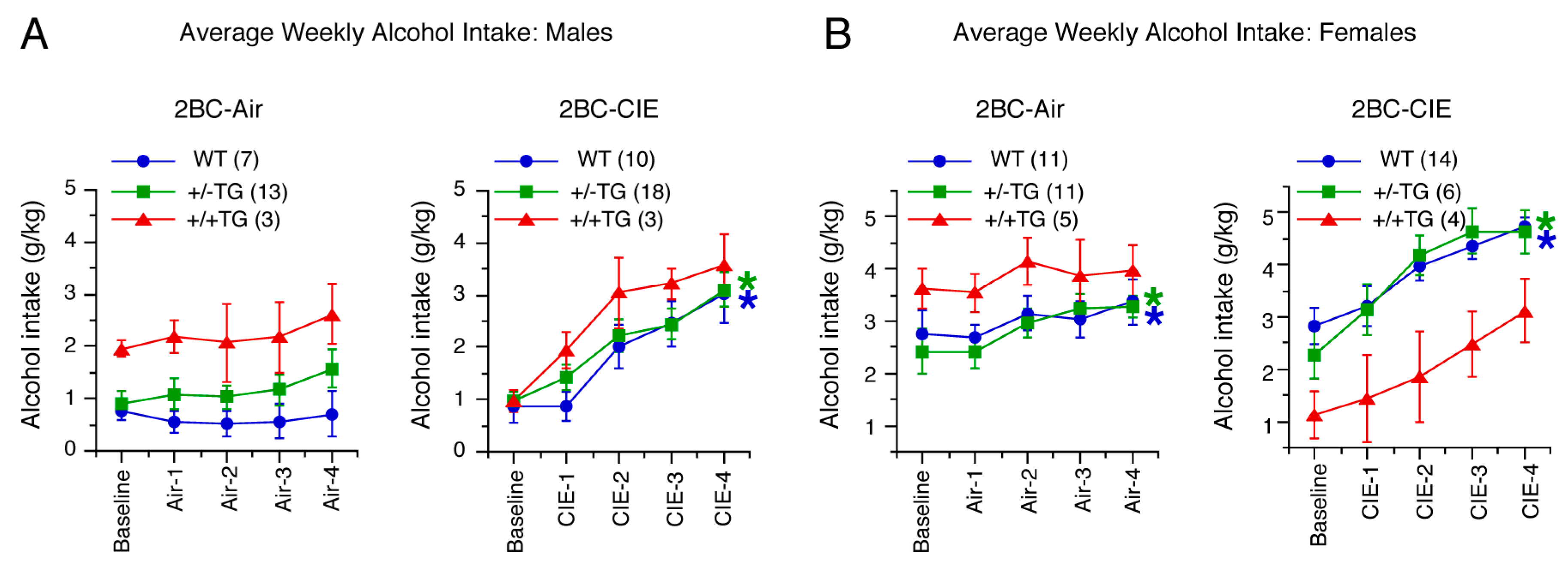
3.1. CIE Treatment Escalates 2BC Alcohol Drinking in All Genotypes
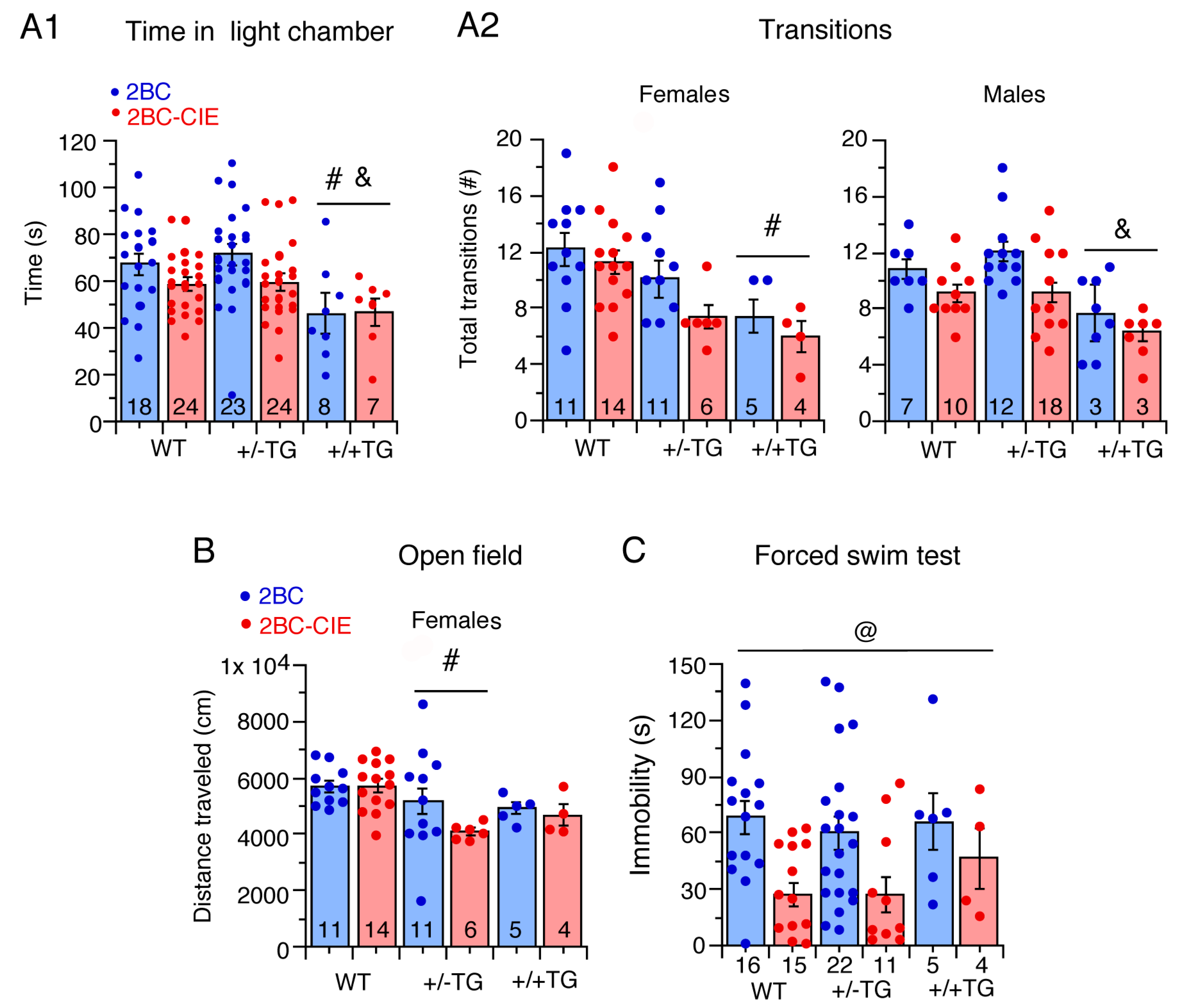
3.2. CIE/Abstinence and Negative Affect/Emotionality
3.2.1. Light/Dark Transfer Test
3.2.2. Digging Test
3.2.3. Open Field Test
3.2.4. Forced Swim Test
3.2.5. Tail Flick Test
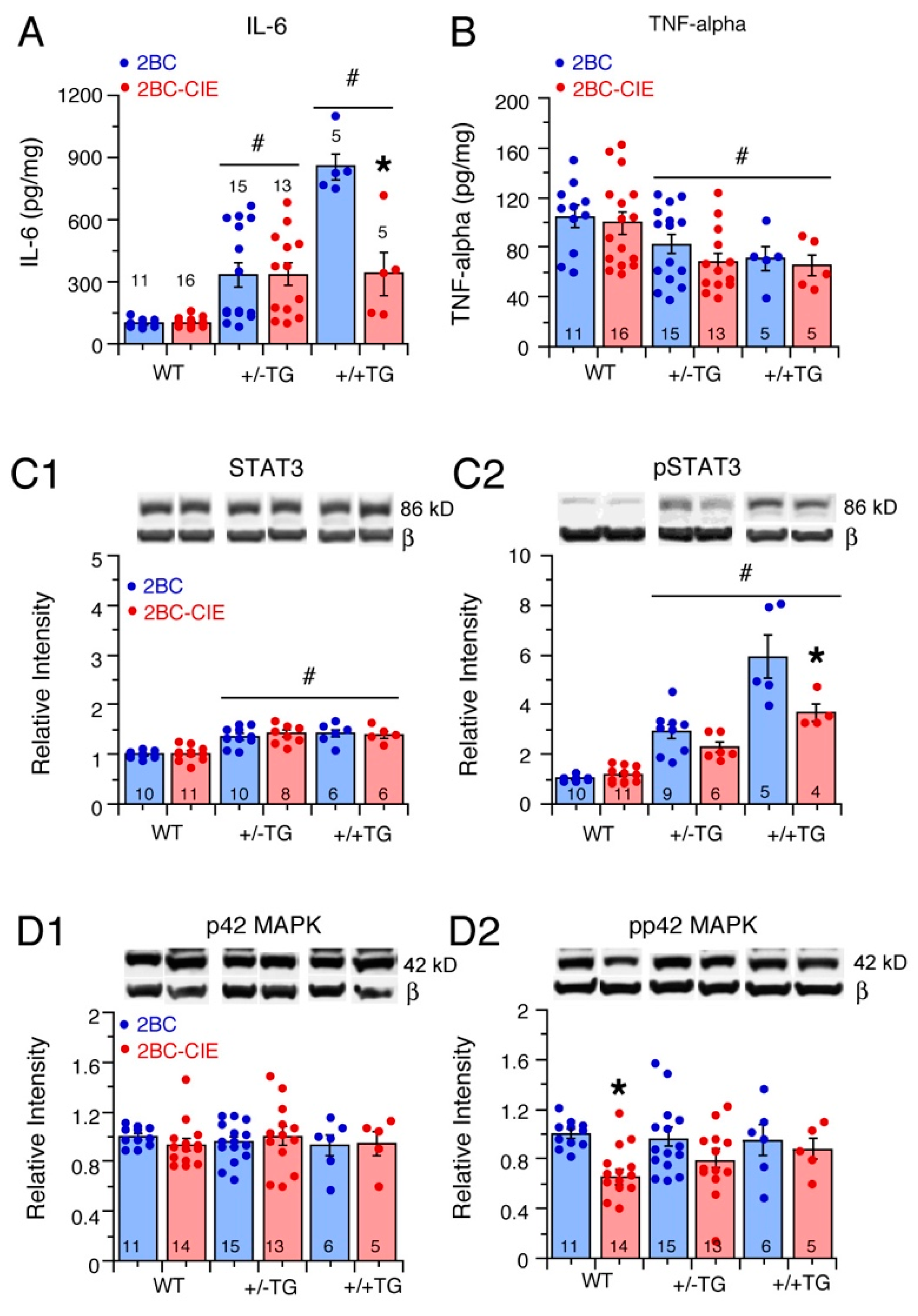
3.3. Effects of 2BC and 2BC-CIE on Neuroimmune Factors in the Hippocampus
3.3.1. IL-6 Levels
3.3.2. TNF-Alpha Levels
3.4. Effects of 2BC and 2BC-CIE on IL-6 Signal Transduction Partners STAT3 and p42/44MAPK in the Hippocampus
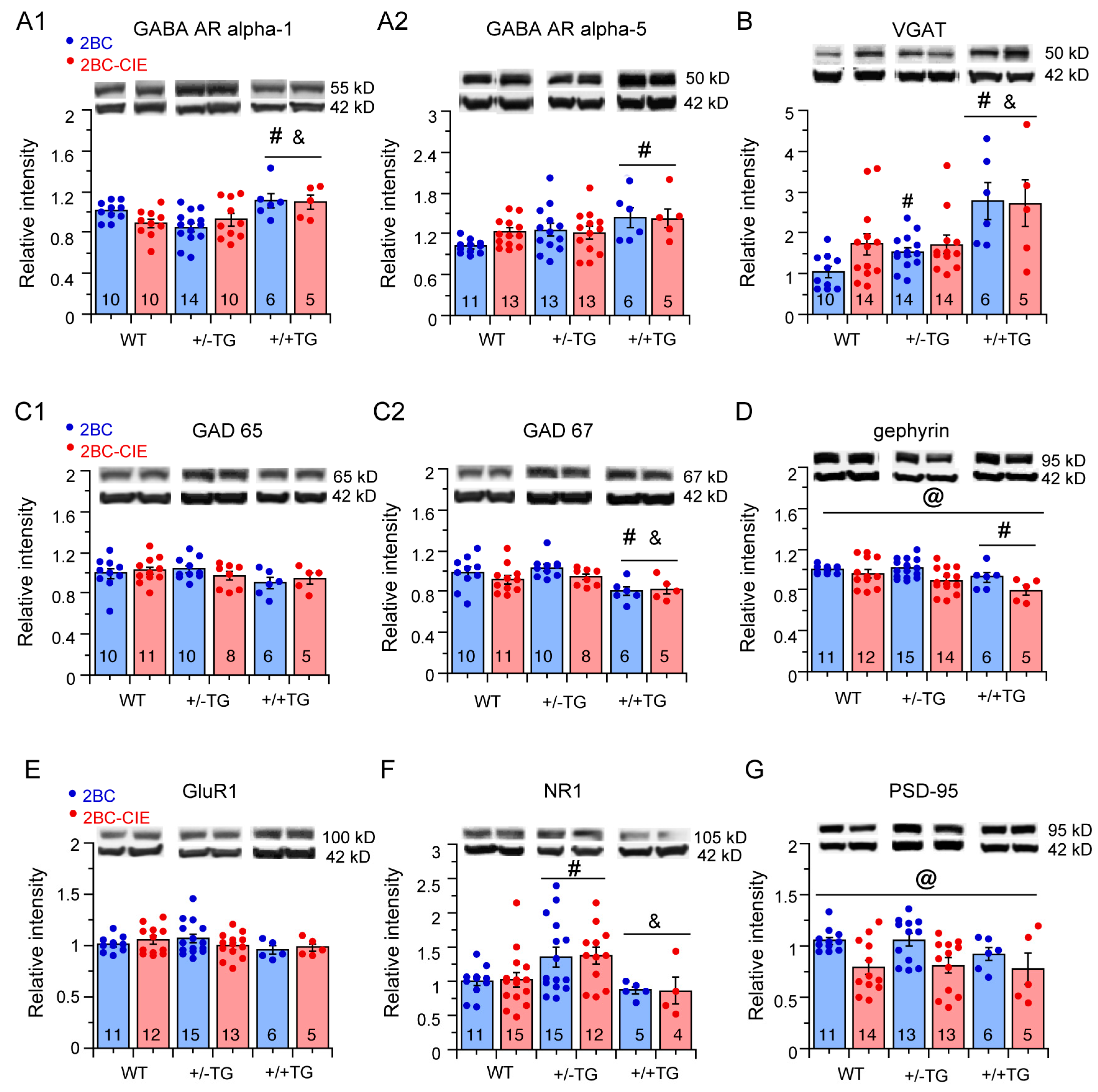
3.5. Effects of 2BC and 2BC-CIE on Inhibitory and Excitatory Synaptic Proteins in the Hippocampus
3.5.1. Inhibitory Synaptic Proteins
3.5.2. Excitatory Synaptic Proteins
4. Discussion
4.1. Alcohol Drinking
4.2. Tests for Negative Affect/Emotionality
4.3. Neuroimmune Factors
4.4. Synaptic Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crews, F.T.; Lawrimore, C.J.; Walter, T.J.; Coleman, L.G., Jr. The role of neuroimmune signaling in alcoholism. Neuropharmacology 2017, 122, 56–73. [Google Scholar] [CrossRef]
- Erickson, E.K.; Grantham, E.K.; Warden, A.S.; Harris, R.A. Neuroimmune signaling in alcohol use disorder. Pharmacol. Biochem. Behav. 2019, 177, 34–60. [Google Scholar] [CrossRef]
- Perkins, A.E.; Varlinskaya, E.I.; Deak, T. From adolescence to late aging: A comprehensive review of social behavior, alcohol, and neuroinflammation across the lifespan. Int. Rev. Neurobiol. 2019, 148, 231–303. [Google Scholar] [CrossRef]
- Roberto, M.; Patel, R.R.; Bajo, M. Ethanol and Cytokines in the Central Nervous System. Handb. Exp. Pharmacol. 2018, 248, 397–431. [Google Scholar] [CrossRef]
- Donegan, J.J.; Girotti, M.; Weinberg, M.S.; Morilak, D.A. A novel role for brain interleukin-6: Facilitation of cognitive flexibility in rat orbitofrontal cortex. J. Neurosci. 2014, 34, 953–962. [Google Scholar] [CrossRef]
- Jankord, R.; Zhang, R.; Flak, J.N.; Solomon, M.B.; Albertz, J.; Herman, J.P. Stress activation of IL-6 neurons in the hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R343–R351. [Google Scholar] [CrossRef] [PubMed]
- Aniszewska, A.; Chlodzinska, N.; Bartkowska, K.; Winnicka, M.M.; Turlejski, K.; Djavadian, R.L. The expression of interleukin-6 and its receptor in various brain regions and their roles in exploratory behavior and stress responses. J. Neuroimmunol. 2015, 284, 1–9. [Google Scholar] [CrossRef]
- Liberman, A.C.; Trias, E.; da Silva Chagas, L.; Trindade, P.; Dos Santos Pereira, M.; Refojo, D.; Hedin-Pereira, C.; Serfaty, C.A. Neuroimmune and Inflammatory Signals in Complex Disorders of the Central Nervous System. Neuroimmunomodulation 2018, 25, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, J.; Alfonso-Loeches, S.; Guerri, C. Impact of the Innate Immune Response in the Actions of Ethanol on the Central Nervous System. Alcohol. Clin. Exp. Res. 2016, 40, 2260–2270. [Google Scholar] [CrossRef] [PubMed]
- Atzori, M.; Garcia-Oscos, F.; Mendez, J.A. Role of IL-6 in the etiology of hyperexcitable neuropsychiatric conditions: Experimental evidence and therapeutic implications. Future Med. Chem. 2012, 4, 2177–2192. [Google Scholar] [CrossRef]
- Dugan, L.L.; Ali, S.S.; Shekhtman, G.; Roberts, A.J.; Lucero, J.; Quick, K.L.; Behrens, M.M. IL-6 mediated degeneration of forebrain GABAergic interneurons and cognitive impairment in aged mice through activation of neuronal NADPH oxidase. PLoS ONE 2009, 4, e5518. [Google Scholar] [CrossRef]
- Wei, H.; Alberts, I.; Li, X. Brain IL-6 and autism. Neuroscience 2013, 252, 320–325. [Google Scholar] [CrossRef]
- Willis, E.F.; MacDonald, K.P.A.; Nguyen, Q.H.; Garrido, A.L.; Gillespie, E.R.; Harley, S.B.R.; Bartlett, P.F.; Schroder, W.A.; Yates, A.G.; Anthony, D.C.; et al. Repopulating Microglia Promote Brain Repair in an IL-6-Dependent Manner. Cell 2020, 180, 833–846.e816. [Google Scholar] [CrossRef] [PubMed]
- Kummer, M.P.; Ising, C.; Kummer, C.; Sarlus, H.; Griep, A.; Vieira-Saecker, A.; Schwartz, S.; Halle, A.; Brückner, M.; Händler, K.; et al. Microglial PD-1 stimulation by astrocytic PD-L1 suppresses neuroinflammation and Alzheimer’s disease pathology. EMBO J. 2021, 40, e108662. [Google Scholar] [CrossRef]
- Kölliker-Frers, R.; Udovin, L.; Otero-Losada, M.; Kobiec, T.; Herrera, M.I.; Palacios, J.; Razzitte, G.; Capani, F. Neuroinflammation: An Integrating Overview of Reactive-Neuroimmune Cell Interactions in Health and Disease. Mediat. Inflamm. 2021, 2021, 9999146. [Google Scholar] [CrossRef] [PubMed]
- Spooren, A.; Kolmus, K.; Laureys, G.; Clinckers, R.; De Keyser, J.; Haegeman, G.; Gerlo, S. Interleukin-6, a mental cytokine. Brain Res. Rev. 2011, 67, 157–183. [Google Scholar] [CrossRef]
- Ye, S.M.; Johnson, R.W. Regulation of interleukin-6 gene expression in brain of aged mice by nuclear factor kappaB. J. Neuroimmunol. 2001, 117, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.R.; Guerin, S.P.; Nixon, K.; Fonken, L.K. The neuroimmune system—Where aging and excess alcohol intersect. Alcohol 2022, 107, 153–167. [Google Scholar] [CrossRef]
- Nunes, P.T.; Kipp, B.T.; Reitz, N.L.; Savage, L.M. Aging with alcohol-related brain damage: Critical brain circuits associated with cognitive dysfunction. Int. Rev. Neurobiol. 2019, 148, 101–168. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Buck, H.M.; Bordner, K.; Richey, L.; Jones, M.E.; Deak, T. Intoxication- and withdrawal-dependent expression of central and peripheral cytokines following initial ethanol exposure. Alcohol. Clin. Exp. Res. 2014, 38, 2186–2198. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Gano, A.; Paniccia, J.E.; Deak, T. Male adolescent rats display blunted cytokine responses in the CNS after acute ethanol or lipopolysaccharide exposure. Physiol. Behav. 2015, 148, 131–144. [Google Scholar] [CrossRef]
- Gano, A.; Pautassi, R.M.; Doremus-Fitzwater, T.L.; Deak, T. Conditioned effects of ethanol on the immune system. Exp. Biol. Med. (Maywood) 2017, 242, 718–730. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Paniccia, J.E.; Gano, A.; Vore, A.S.; Deak, T. Differential effects of acute versus chronic stress on ethanol sensitivity: Evidence for interactions on both behavioral and neuroimmune outcomes. Brain. Behav. Immun. 2018, 70, 141–156. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, Y.; Tang, X.; Jing, C.; Qiu, S.; Li, B.; Li, Y. IL-6 and IL-1β upregulation and tau protein phosphorylation in response to chronic alcohol exposure in the mouse hippocampus. Neuroreport 2021, 32, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Baxter-Potter, L.N.; Henricks, A.M.; Berger, A.L.; Bieniasz, K.V.; Lugo, J.M.; McLaughlin, R.J. Alcohol vapor exposure differentially impacts mesocorticolimbic cytokine expression in a sex-, region-, and duration-specific manner. Neuroscience 2017, 346, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R., Jr.; Bandiera, S.; Souza, D.G.; Bellaver, B.; Caletti, G.; Quincozes-Santos, A.; Elisabetsky, E.; Gomez, R. N-acetylcysteine Prevents Alcohol Related Neuroinflammation in Rats. Neurochem. Res. 2017, 42, 2135–2141. [Google Scholar] [CrossRef]
- Gruol, D.L.; Melkonian, C.; Huitron-Resendiz, S.; Roberts, A.J. Alcohol alters IL-6 Signal Transduction in the CNS of Transgenic Mice with Increased Astrocyte Expression of IL-6. Cell. Mol. Neurobiol. 2020, 41, 733–750. [Google Scholar] [CrossRef]
- Gruol, D.L.; Calderon, D.; French, K.; Melkonian, C.; Huitron-Resendiz, S.; Cates-Gatto, C.; Roberts, A.J. Neuroimmune interactions with binge alcohol drinking in the cerebellum of IL-6 transgenic mice. Neuropharmacology 2023, 228, 109455. [Google Scholar] [CrossRef]
- Chen, W.Y.; Chen, H.; Hamada, K.; Gatta, E.; Chen, Y.; Zhang, H.; Drnevich, J.; Krishnan, H.R.; Maienschein-Cline, M.; Grayson, D.R.; et al. Transcriptomics identifies STAT3 as a key regulator of hippocampal gene expression and anhedonia during withdrawal from chronic alcohol exposure. Transl. Psychiatry 2021, 11, 298. [Google Scholar] [CrossRef] [PubMed]
- Gruol, D.L.; Huitron-Resendiz, S.; Roberts, A.J. Altered brain activity during withdrawal from chronic alcohol is associated with changes in IL-6 signal transduction and GABAergic mechanisms in transgenic mice with increased astrocyte expression of IL-6. Neuropharmacology 2018, 138, 32–46. [Google Scholar] [CrossRef]
- Sukoff Rizzo, S.J.; Neal, S.J.; Hughes, Z.A.; Beyna, M.; Rosenzweig-Lipson, S.; Moss, S.J.; Brandon, N.J. Evidence for sustained elevation of IL-6 in the CNS as a key contributor of depressive-like phenotypes. Transl. Psychiatry 2012, 2, e199. [Google Scholar] [CrossRef]
- Ting, E.Y.; Yang, A.C.; Tsai, S.J. Role of Interleukin-6 in Depressive Disorder. Int. J. Mol. Sci. 2020, 21, 2194. [Google Scholar] [CrossRef]
- Hao, Y.; Jing, H.; Bi, Q.; Zhang, J.; Qin, L.; Yang, P. Intra-amygdala microinfusion of IL-6 impairs the auditory fear conditioning of rats via JAK/STAT activation. Behav. Brain Res. 2014, 275, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Armario, A.; Hernandez, J.; Bluethmann, H.; Hidalgo, J. IL-6 deficiency leads to increased emotionality in mice: Evidence in transgenic mice carrying a null mutation for IL-6. J. Neuroimmunol. 1998, 92, 160–169. [Google Scholar] [CrossRef]
- Baier, P.C.; May, U.; Scheller, J.; Rose-John, S.; Schiffelholz, T. Impaired hippocampus-dependent and -independent learning in IL-6 deficient mice. Behav. Brain Res. 2009, 200, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Heyser, C.J.; Masliah, E.; Samimi, A.; Campbell, I.L.; Gold, L.H. Progressive decline in avoidance learning paralleled by inflammatory neurodegeneration in transgenic mice expressing interleukin 6 in the brain. Proc. Natl. Acad. Sci. USA 1997, 94, 1500–1505. [Google Scholar] [CrossRef]
- Ma, T.C.; Zhu, X.Z. Intrahippocampal infusion of interleukin-6 impairs avoidance learning in rats. Zhongguo Yao Li Xue Bao 1997, 18, 121–123. [Google Scholar]
- Wu, T.H.; Lin, C.H. IL-6 mediated alterations on immobile behavior of rats in the forced swim test via ERK1/2 activation in specific brain regions. Behav. Brain Res. 2008, 193, 183–191. [Google Scholar] [CrossRef]
- Hryniewicz, A.; Bialuk, I.; Kaminski, K.A.; Winnicka, M.M. Impairment of recognition memory in interleukin-6 knock-out mice. Eur. J. Pharmacol. 2007, 577, 219–220. [Google Scholar] [CrossRef]
- Bialuk, I.; Taranta, A.; Winnicka, M.M. IL-6 deficiency alters spatial memory in 4- and 24-month-old mice. Neurobiol. Learn. Mem. 2018, 155, 21–29. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Ponomarev, I.; Geil, C.; Bergeson, S.; Koob, G.F.; Harris, R.A. Neuroimmune regulation of alcohol consumption: Behavioral validation of genes obtained from genomic studies. Addict. Biol. 2012, 17, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Gruol, D.L. IL-6 regulation of synaptic function in the CNS. Neuropharmacology 2015, 96, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Gruol, D.L. Neuroimmune regulation of neurophysiology in the cerebellum. Cerebellum 2013, 12, 307–309. [Google Scholar] [CrossRef]
- Tancredi, V.; D’Antuono, M.; Cafe, C.; Giovedi, S.; Bue, M.C.; D’Arcangelo, G.; Onofri, F.; Benfenati, F. The inhibitory effects of interleukin-6 on synaptic plasticity in the rat hippocampus are associated with an inhibition of mitogen-activated protein kinase ERK. J. Neurochem. 2000, 75, 634–643. [Google Scholar] [CrossRef]
- D’Arcangelo, G.; Tancredi, V.; Onofri, F.; D’Antuono, M.; Giovedì, S.; Benfenati, F. Interleukin-6 inhibits neurotransmitter release and the spread of excitation in the rat cerebral cortex. Eur. J. Neurosci. 2000, 12, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Li, A.J.; Katafuchi, T.; Oda, S.; Hori, T.; Oomura, Y. Interleukin-6 inhibits long-term potentiation in rat hippocampal slices. Brain Res. 1997, 748, 30–38. [Google Scholar] [CrossRef]
- Garcia-Oscos, F.; Salgado, H.; Hall, S.; Thomas, F.; Farmer, G.E.; Bermeo, J.; Galindo, L.C.; Ramirez, R.D.; D’Mello, S.; Rose-John, S.; et al. The stress-induced cytokine interleukin-6 decreases the inhibition/excitation ratio in the rat temporal cortex via trans-signaling. Biol. Psychiatry 2012, 71, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Balschun, D.; Wetzel, W.; Del Rey, A.; Pitossi, F.; Schneider, H.; Zuschratter, W.; Besedovsky, H.O. Interleukin-6: A cytokine to forget. FASEB J. 2004, 18, 1788–1790. [Google Scholar] [CrossRef]
- Bellinger, F.P.; Madamba, S.G.; Campbell, I.L.; Siggins, G.R. Reduced long-term potentiation in the dentate gyrus of transgenic mice with cerebral overexpression of interleukin-6. Neurosci. Lett. 1995, 198, 95–98. [Google Scholar] [CrossRef]
- Roberts, A.J.; Khom, S.; Bajo, M.; Vlkolinsky, R.; Polis, I.; Cates-Gatto, C.; Roberto, M.; Gruol, D.L. Increased IL-6 expression in astrocytes is associated with emotionality, alterations in central amygdala GABAergic transmission, and excitability during alcohol withdrawal. Brain. Behav. Immun. 2019, 82, 188–202. [Google Scholar] [CrossRef]
- Bloch, S.; Holleran, K.M.; Kash, T.L.; Vazey, E.M.; Rinker, J.A.; Lebonville, C.L.; O’Hara, K.; Lopez, M.F.; Jones, S.R.; Grant, K.A.; et al. Assessing negative affect in mice during abstinence from alcohol drinking: Limitations and future challenges. Alcohol 2022, 100, 41–56. [Google Scholar] [CrossRef]
- Bartley, P.C.; Rezvani, A.H. Alcohol and cognition—Consideration of age of initiation, usage patterns and gender: A brief review. Curr. Drug Abus. Rev. 2012, 5, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Lees, B.; Meredith, L.R.; Kirkland, A.E.; Bryant, B.E.; Squeglia, L.M. Effect of alcohol use on the adolescent brain and behavior. Pharmacol. Biochem. Behav. 2020, 192, 172906. [Google Scholar] [CrossRef]
- Cheaha, D.; Keawpradub, N.; Sawangjaroen, K.; Phukpattaranont, P.; Kumarnsit, E. Effects of an alkaloid-rich extract from Mitragyna speciosa leaves and fluoxetine on sleep profiles, EEG spectral frequency and ethanol withdrawal symptoms in rats. Phytomedicine 2015, 22, 1000–1008. [Google Scholar] [CrossRef]
- Sidhu, H.; Kreifeldt, M.; Contet, C. Affective Disturbances During Withdrawal from Chronic Intermittent Ethanol Inhalation in C57BL/6J and DBA/2J Male Mice. Alcohol. Clin. Exp. Res. 2018. [Google Scholar] [CrossRef]
- Farr, S.A.; Scherrer, J.F.; Banks, W.A.; Flood, J.F.; Morley, J.E. Chronic ethanol consumption impairs learning and memory after cessation of ethanol. Alcohol. Clin. Exp. Res. 2005, 29, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.V.; Puro, A.C.; Manos, J.C.; Huitron-Resendiz, S.; Reyes, K.C.; Liu, K.; Vo, K.; Roberts, A.J.; Gruol, D.L. Transgenic mice with increased astrocyte expression of IL-6 show altered effects of acute ethanol on synaptic function. Neuropharmacology 2016, 103, 27–43. [Google Scholar] [CrossRef]
- Gruol, D.L.; Hernandez, R.V.; Roberts, A. Alcohol Enhances Responses to High Frequency Stimulation in Hippocampus from Transgenic Mice with Increased Astrocyte Expression of IL-6. Cell. Mol. Neurobiol. 2020, 41, 1299–1310. [Google Scholar] [CrossRef]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The Rodent Forced Swim Test Measures Stress-Coping Strategy, Not Depression-like Behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef]
- Holleran, K.M.; Winder, D.G. Preclinical voluntary drinking models for alcohol abstinence-induced affective disturbances in mice. Genes Brain Behav. 2017, 16, 8–14. [Google Scholar] [CrossRef]
- März, P.; Gadient, R.A.; Otten, U. Expression of interleukin-6 receptor (IL-6R) and gp130 mRNA in PC12 cells and sympathetic neurons: Modulation by tumor necrosis factor alpha (TNF-alpha). Brain Res. 1996, 706, 71–79. [Google Scholar] [CrossRef]
- Matsumoto, J.; Dohgu, S.; Takata, F.; Machida, T.; Bölükbaşi Hatip, F.F.; Hatip-Al-Khatib, I.; Yamauchi, A.; Kataoka, Y. TNF-α-sensitive brain pericytes activate microglia by releasing IL-6 through cooperation between IκB-NFκB and JAK-STAT3 pathways. Brain Res. 2018, 1692, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Sawada, M.; Suzumura, A.; Marunouchi, T. TNF alpha induces IL-6 production by astrocytes but not by microglia. Brain Res. 1992, 583, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, N.J.; Oh, J.W.; Repovic, P.; Benveniste, E.N. Interleukin-6 (IL-6) production by astrocytes: Autocrine regulation by IL-6 and the soluble IL-6 receptor. J. Neurosci. 1999, 19, 5236–5244. [Google Scholar] [CrossRef]
- Campbell, I.L.; Abraham, C.R.; Masliah, E.; Kemper, P.; Inglis, J.D.; Oldstone, M.B.; Mucke, L. Neurologic disease induced in transgenic mice by cerebral overexpression of interleukin 6. Proc. Natl. Acad. Sci. USA 1993, 90, 10061–10065. [Google Scholar] [CrossRef]
- Gruol, D.L.; Vo, K.; Bray, J.G. Increased astrocyte expression of IL-6 or CCL2 in transgenic mice alters levels of hippocampal and cerebellar proteins. Front. Cell. Neurosci. 2014, 8, 234. [Google Scholar] [CrossRef]
- Becker, H.C.; Lopez, M.F. Increased ethanol drinking after repeated chronic ethanol exposure and withdrawal experience in C57BL/6 mice. Alcohol. Clin. Exp. Res. 2004, 28, 1829–1838. [Google Scholar] [CrossRef]
- Chu, K.; Koob, G.F.; Cole, M.; Zorrilla, E.P.; Roberts, A.J. Dependence-induced increases in ethanol self-administration in mice are blocked by the CRF1 receptor antagonist antalarmin and by CRF1 receptor knockout. Pharmacol. Biochem. Behav. 2007, 86, 813–821. [Google Scholar] [CrossRef]
- Schweitzer, P.; Cates-Gatto, C.; Varodayan, F.P.; Nadav, T.; Roberto, M.; Lasek, A.W.; Roberts, A.J. Dependence-induced ethanol drinking and GABA neurotransmission are altered in Alk deficient mice. Neuropharmacology 2016, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.; Wrenn, C.C.; Harris, A.P.; Thayer, K.E.; Crawley, J.N. Behavioral profiles of inbred strains on novel olfactory, spatial and emotional tests for reference memory in mice. Genes Brain Behav. 2002, 1, 55–69. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Cao, B.J.; Dalvi, A.; Holmes, A. Animal models of anxiety: An ethological perspective. Braz. J. Med. Biol. Res. 1997, 30, 289–304. [Google Scholar] [CrossRef]
- Deacon, R.M. Digging and marble burying in mice: Simple methods for in vivo identification of biological impacts. Nat. Protoc. 2006, 1, 122–124. [Google Scholar] [CrossRef]
- Raible, D.J.; Frey, L.C.; Brooks-Kayal, A.R. Effects of JAK2-STAT3 signaling after cerebral insults. Jak-Stat 2014, 3, e29510. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N. Behavioral phenotyping of transgenic and knockout mice: Experimental design and evaluation of general health, sensory functions, motor abilities, and specific behavioral tests. Brain Res. 1999, 835, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Markou, A.; Lucki, I. Assessing antidepressant activity in rodents: Recent developments and future needs. Trends Pharmacol. Sci. 2002, 23, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D. Animal models of depression: Utility for transgenic research. Rev. Neurosci. 2000, 11, 53–58. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral despair in mice: A primary screening test for antidepressants. Arch. Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar]
- Lucki, I. The forced swimming test as a model for core and component behavioral effects of antidepressant drugs. Behav. Pharmacol. 1997, 8, 523–532. [Google Scholar] [CrossRef]
- Athanason, A.C.; Nadav, T.; Cates-Gatto, C.; Roberts, A.J.; Roberto, M.; Varodayan, F.P. Chronic ethanol alters adrenergic receptor gene expression and produces cognitive deficits in male mice. Neurobiol. Stress 2023, 24, 100542. [Google Scholar] [CrossRef]
- Okhuarobo, A.; Angelo, M.; Bolton, J.L.; Lopez, C.; Igbe, I.; Baram, T.Z.; Contet, C. Influence of early-life adversity on responses to acute and chronic ethanol in female mice. Alcohol. Clin. Exp. Res. 2023, 47, 336–347. [Google Scholar] [CrossRef]
- Gruol, D.L.; Melkonian, C.; Ly, K.; Sisouvanthong, J.; Tan, Y.; Roberts, A.J. Alcohol and IL-6 alter expression of synaptic proteins in cerebellum of transgenic mice with increased astrocyte expression of IL-6. Neuroscience 2020, 442, 124–137. [Google Scholar] [CrossRef]
- Borgonetti, V.; Roberts, A.J.; Bajo, M.; Galeotti, N.; Roberto, M. Chronic alcohol induced mechanical allodynia by promoting neuroinflammation: A mouse model of alcohol-evoked neuropathic pain. Br. J. Pharmacol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Hilderbrand, E.R.; Lasek, A.W. Studying Sex Differences in Animal Models of Addiction: An Emphasis on Alcohol-Related Behaviors. ACS Chem. Neurosci. 2018, 9, 1907–1916. [Google Scholar] [CrossRef]
- Yoneyama, N.; Crabbe, J.C.; Ford, M.M.; Murillo, A.; Finn, D.A. Voluntary ethanol consumption in 22 inbred mouse strains. Alcohol 2008, 42, 149–160. [Google Scholar] [CrossRef] [PubMed]
- McCool, B.A.; Chappell, A.M. Chronic intermittent ethanol inhalation increases ethanol self-administration in both C57BL/6J and DBA/2J mice. Alcohol 2015, 49, 111–120. [Google Scholar] [CrossRef]
- Roberto, M.; Nelson, T.E.; Ur, C.L.; Brunelli, M.; Sanna, P.P.; Gruol, D.L. The transient depression of hippocampal CA1 LTP induced by chronic intermittent ethanol exposure is associated with an inhibition of the MAP kinase pathway. Eur. J. Neurosci. 2003, 17, 1646–1654. [Google Scholar] [CrossRef]
- Brantley, E.C.; Benveniste, E.N. Signal transducer and activator of transcription-3: A molecular hub for signaling pathways in gliomas. Mol. Cancer Res. 2008, 6, 675–684. [Google Scholar] [CrossRef]
- Dziennis, S.; Alkayed, N.J. Role of signal transducer and activator of transcription 3 in neuronal survival and regeneration. Rev. Neurosci. 2008, 19, 341–361. [Google Scholar] [PubMed]
- Nicolas, C.S.; Amici, M.; Bortolotto, Z.A.; Doherty, A.; Csaba, Z.; Fafouri, A.; Dournaud, P.; Gressens, P.; Collingridge, G.L.; Peineau, S. The role of JAK-STAT signaling within the CNS. Jak-Stat 2013, 2, e22925. [Google Scholar] [CrossRef] [PubMed]
- Fraga-Junior, E.B.; Fernandes, I.L.; Rohden, C.A.H.; Doneda, D.L.; Ynoue, H.N.; Rios-Santos, F.; Lima, E.; Buss, Z.S.; Vandresen-Filho, S. Attenuation of the levels of pro-inflammatory cytokines prevents depressive-like behavior during ethanol withdrawal in mice. Brain Res. Bull. 2022, 191, 9–19. [Google Scholar] [CrossRef]
- Fasick, V.; Spengler, R.N.; Samankan, S.; Nader, N.D.; Ignatowski, T.A. The hippocampus and TNF: Common links between chronic pain and depression. Neurosci. Biobehav. Rev. 2015, 53, 139–159. [Google Scholar] [CrossRef]
- Prenosil, G.A.; Schneider Gasser, E.M.; Rudolph, U.; Keist, R.; Fritschy, J.M.; Vogt, K.E. Specific subtypes of GABAA receptors mediate phasic and tonic forms of inhibition in hippocampal pyramidal neurons. J. Neurophysiol. 2006, 96, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Van Skike, C.E.; Diaz-Granados, J.L.; Matthews, D.B. Chronic intermittent ethanol exposure produces persistent anxiety in adolescent and adult rats. Alcohol. Clin. Exp. Res. 2015, 39, 262–271. [Google Scholar] [CrossRef]
- Charlton, M.E.; Sweetnam, P.M.; Fitzgerald, L.W.; Terwilliger, R.Z.; Nestler, E.J.; Duman, R.S. Chronic ethanol administration regulates the expression of GABAA receptor alpha 1 and alpha 5 subunits in the ventral tegmental area and hippocampus. J. Neurochem. 1997, 68, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Grobin, A.C.; Papadeas, S.T.; Morrow, A.L. Regional variations in the effects of chronic ethanol administration on GABA(A) receptor expression: Potential mechanisms. Neurochem. Int. 2000, 37, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Mhatre, M.C.; Pena, G.; Sieghart, W.; Ticku, M.K. Antibodies specific for GABAA receptor alpha subunits reveal that chronic alcohol treatment down-regulates alpha-subunit expression in rat brain regions. J. Neurochem. 1993, 61, 1620–1625. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Test | Post Alcohol Day of Testing | Method of Measuring Behavior | Used to Assess |
|---|---|---|---|
| light/dark transfer test | 3 days post alcohol | scored by experimenter blind to genotype and treatment | anxiety-like behavior, exploratory drive |
| digging test | 4 days post alcohol | live scored by experimenter blind to genotype and treatment | response to novelty and possibly irritability |
| open field test | 5 days post alcohol | Noldus Ethovision XT v14 software | emotionality, activity |
| forced swim test | 6 days post alcohol | scored by experimenter blind to genotype and treatment | helpless-like behavior |
| tail flick test | 7 days post alcohol | automated scoring by device (Ugo Basile) | pain sensitivity |
| Behavioral Test (Mice 3–8 Days Withdrawn) | TG+/− vs. WT | TG +/+ vs. WT | TG +/+ vs. TG+/− | 2BC-CIE vs. 2BC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Naïve 1 | 2BC | 2BC- CIE | Naïve 1 | 2BC | 2BC- CIE | Naïve 1 | 2BC | 2BC- CIE | WT | +/− TG | +/+ TG | |
| 1. Light/dark transfer | ||||||||||||
| -time spent in light | ns | overall ns | ns | overall ↓ | ns | overall ↓ | overall ns | |||||
| -#of transitions | ns | overall ns | ↓ | M & F ↓ | ns | overall M ↓ | overall M ↓ | |||||
| 2. Open field test | ||||||||||||
| -distance | ns | overall F ↓ | ↓ | overall ns | ns | overall ns | overall ns | |||||
| -velocity | ns | overall ns | ↓ | overall ns | ns | overall ns | overall ns | |||||
| -center time | ns | overall ns | ns | overall ns | ns | overall ns | overall ns | |||||
| 3. Digging bouts | ns | overall ns | ↓ | overall ns | ns | overall ns | overall ns | |||||
| 4. Forced swim test | ||||||||||||
| -Immobility | ns | overall ns | ↑ | overall ns | ns | overall ns | overall ↓ | |||||
| 5. Tail flick test | ns | overall ns | ns | overall ns | ns | overall ns | overall ↓ | |||||
| Protein | Alcohol-Naïve 1 Hippocampus | Alcohol-Exposed, 3–8 Days Abstinent Hippocampus | CIE (24 Withdrawn) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TG vs. WT | 2BC TG vs. WT | 2BC-CIE TG vs. WT | 2BC-CIE vs. 2BC | Hippocampus 2 | Cerebellum 3 | ||||||||
| +/− TG | +/+ TG | +/− TG | +/+ TG | +/− TG | +/+ TG | WT | +/− TG | +/+ TG | WT vs. Naïve | +/−TG vs. Naïve | WT vs. Naïve | +/−TG vs. Naïve | |
| 1. Neuroimmune | |||||||||||||
| IL-6 | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ns | ns | ↓ | - | - | ns | ↓ |
| TNF-alpha | - | - | +/−TG and +/+TG ↓ | overall ns | - | - | - | - | |||||
| 2. Signal transduction | |||||||||||||
| STAT3 | ↑ | ↑ | +/−TG and +/+TG ↑ | overall ns | ns | ns | ↓ | ns | |||||
| pSTAT3 | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ns | ns | ↓ | ↑ | ↓ | ↑ | ↓ |
| p42MAPK | ns | - | overall ns | - | - | ns | ns | ||||||
| pp42MAPK | ns | - | ns | ns | ns | ns | ↓ | ns | ns | - | - | ns | ↓ |
| p44MAPK | ns | - | overall ns | - | - | ns | ns | ||||||
| pp44MAPK | ns | - | overall ns | - | - | ns | ↓ | ||||||
| Protein | Alcohol-Naïve 1 | Alcohol-Exposed, Abstinent 3–8 Days | CIE, Abstinent 24 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hippocampus | Hippocampus | Hippocampus 2BC-CIE vs. 2BC | Hippocampus 2 | Cerebellum 3 | ||||||||
| +/−TG vs. WT | +/+TG vs. WT | +/−TG vs. WT | +/+TG vs. WT | +/+TG vs. +/− TG | WT | +/− TG | +/+ TG | WT vs. Naïve | +/− vs. Naïve | WT vs. Naïve | +/−TG vs. Naive | |
| 1. Inhibitory synaptic transmission | ||||||||||||
| GABAAR a-1 | ns | ns | ns | ↑ | ↑ | overall ns | ns | ns | ns | ns | ||
| GABAAR a-5 | ↑ | ↑ | ns | ↑ | ns | overall ns | ↓ | ↓ | ↓ | ↓ | ||
| GAD 65 | ns | ↓ | overall ns | overall ns | ns | ns | ns | ↓ | ||||
| GAD 67 | ns | ↓ | ns | ↓ | ↓ | overall ns | ns | ns | ns | ↓ | ||
| VGAT | ↑ | - | ↑ M | ↑ F,M | ↑ F | overall ns | ↓ | ↓ | ↓ | ↓ | ||
| Gephyrin | ↓ | ↓ | ns | ↓ | ns | overall ↓ | ns | ns | ↓ | ns | ||
| 2. Excitatory synaptic transmission | ||||||||||||
| GluR1 | ns | ns | ns overall ns | overall ns | - | - | ↓ | ns | ||||
| NR1 | ns | ns | ↑ | ns | ↓ | overall ns | - | - | - | - | ||
| PSD95 | ns | ns | overall ns | overall ↓ | - | - | - | - | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruol, D.L.; Calderon, D.; Huitron-Resendiz, S.; Cates-Gatto, C.; Roberts, A.J. Impact of Elevated Brain IL-6 in Transgenic Mice on the Behavioral and Neurochemical Consequences of Chronic Alcohol Exposure. Cells 2023, 12, 2306. https://doi.org/10.3390/cells12182306
Gruol DL, Calderon D, Huitron-Resendiz S, Cates-Gatto C, Roberts AJ. Impact of Elevated Brain IL-6 in Transgenic Mice on the Behavioral and Neurochemical Consequences of Chronic Alcohol Exposure. Cells. 2023; 12(18):2306. https://doi.org/10.3390/cells12182306
Chicago/Turabian StyleGruol, Donna L., Delilah Calderon, Salvador Huitron-Resendiz, Chelsea Cates-Gatto, and Amanda J. Roberts. 2023. "Impact of Elevated Brain IL-6 in Transgenic Mice on the Behavioral and Neurochemical Consequences of Chronic Alcohol Exposure" Cells 12, no. 18: 2306. https://doi.org/10.3390/cells12182306