
Riboflavin and Its Derivates as Potential Photosensitizers in the Photodynamic Treatment of Skin Cancers

Abstract
:1. Introduction
2. Blue Light in PDT Therapy for Skin Cancer and Diseases
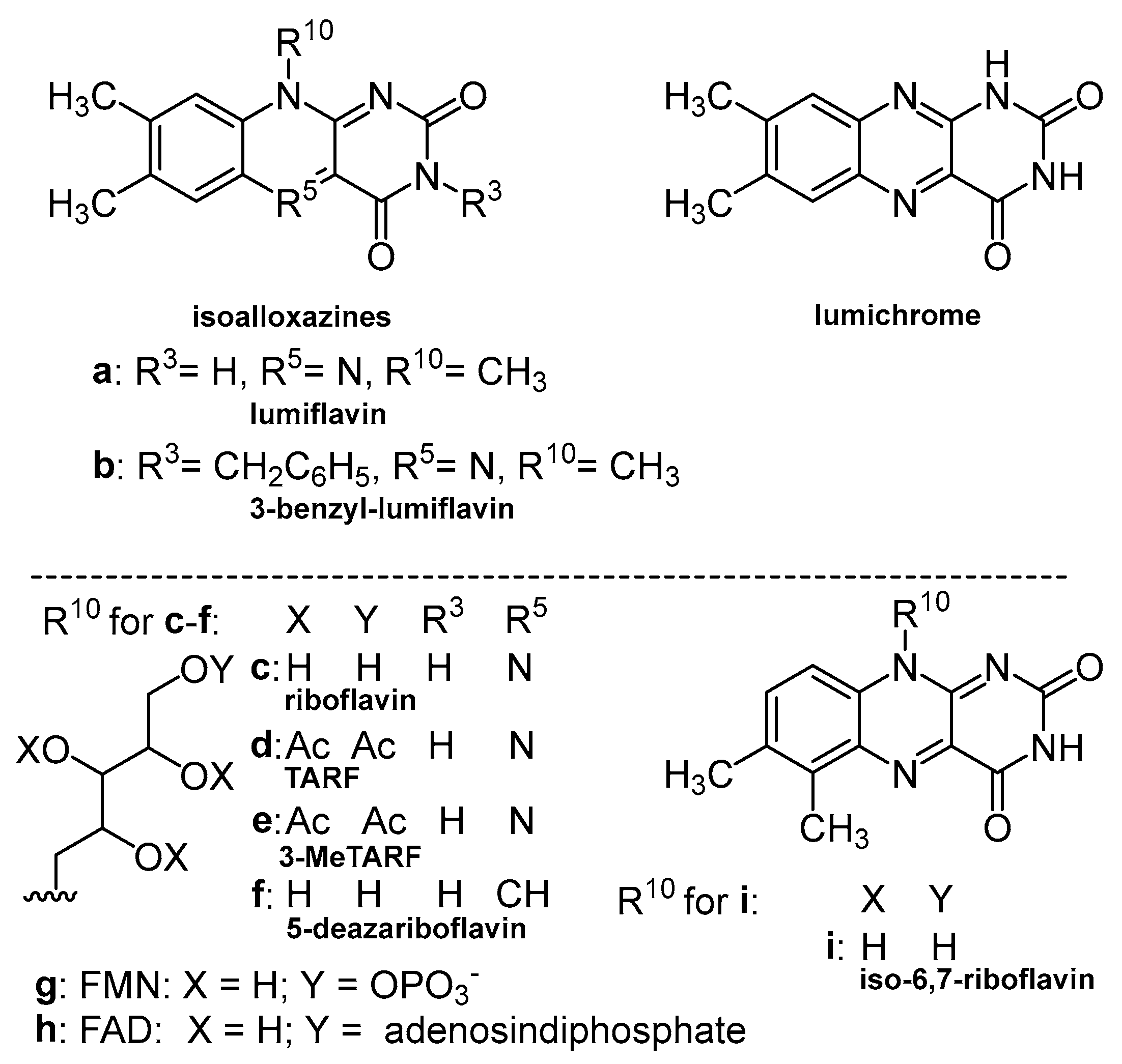
3. Photoreactivity of Flavin Derivatives: Photochemical Aspects
4. Photosensitization with Flavin Sensitizers: Biophysical Approach
5. Riboflavin and Its Derivates in PDT: Biological Issues
6. Apoptosis as a Mechanism of 3MeTARF-Induced Melanoma and Non-Melanoma Skin Cancer Cell Death
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agostinis, P.; Breyssens, H.; Buytaert, E.; Hendrickx, N. Regulatory pathways in photodynamic therapy induced apoptosis. Photochem. Photobiol. Sci. 2004, 3, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Castano, A.P.; Demidova, T.N.; Hamblin, M.R. Mechanisms in photodynamic therapy: Part one—Photosensitizers, photo-chemistry and cellular localization. Photodiagn. Photodyn. 2004, 1, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Fotodynamiczna Metoda Rozpoznawania i Leczenia Nowotworów; Scientific editoship Alfreda Graczyk; Publishing House Bellona: Warsaw, Poland, 1999.
- Songca, S.P.; Adjei, Y. Applications of antimicrobial photodynamic therapy against bacterial biofilms. Int. J. Mol. Sci. 2022, 23, 3209. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.J.; Jemec, G.B.; Arendrup, M.C.; Saunte, D.M.L. Photodynamic therapy treatment of superficial fungal infections: A systematic review. Photodiagnosis Photodyn. Ther. 2020, 31, 101774. [Google Scholar] [CrossRef]
- Piksa, M.; Lian, C.; Samuel, I.C.; Pawlik, K.J.; Samuel, I.D.W.; Matczyszyn, K. The role of the light source in antimicrobial photodynamic therapy. Chem. Soc. Rev. 2023, 52, 1697–1722. [Google Scholar] [CrossRef]
- Gilaberte, Y.; Rezusta, A.; Juarranz, A.; Hamblin, M.R. Editorial: Antimicrobial photodynamic therapy: A new paradigm in the fight against infections. Front. Med. 2021, 8, 788888. [Google Scholar] [CrossRef]
- Juzeniene, A.; Peng, Q.; Moan, J. Milestones in the development of photodynamic therapy and fluorescence diagnosis. Photochem. Photobiol. Sci. 2007, 6, 1234–1245. [Google Scholar] [CrossRef]
- Mroz, P.; Tegos, G.P.; Gali, H.; Wharton, T.; Sarna, T.; Hamblin, M.R. Photodynamic therapy with fullerenes. Photochem. Photobiol. Sci. 2007, 6, 1139–1149. [Google Scholar] [CrossRef]
- Vatansever, F.; de Melo, W.C.M.A.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered, around reactive oxygen species—Bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef]
- Bastos, E.L.; Quina, F.H.; Baptista, M.S. Endogenous photosensitizers in human skin. Chem. Rev. 2023, 123, 9720–9785. [Google Scholar] [CrossRef]
- Iqbal, J.; Husain, A.; Gupta, A. Photooxidation of acyclovir with thermally generated triplet excited ketones. A comparison with Type I and II photosensitizers. Chem. Pharm. Bull. 2006, 54, 519–521. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Cadet, J.; Greer, A.; Thomas, A.H. Photosensitization reactions of biomolecules: Definition, targets and mechanisms. Photochem. Photobiol. 2021, 97, 1456–1483. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Cadet, J.; Di Mascio, P.; Ghogare, A.A.; Greer, A.; Hamblin, M.R.; Lorente, C.; Nunez, S.C.; Ribeiro, M.S.; Thomas, A.H.; et al. Type I and Type II photosensitized oxidation reactions: Guidelines and mechanistic pathways. Photochem. Photobiol. 2017, 93, 912–919. [Google Scholar] [CrossRef]
- Min, D.B.; Boff, J.M. Chemistry and reaction of singlet oxygen in foods. Compr. Rev. Food Sci. Food Saf. 2002, 1, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Mang, T.S. Lasers and light sources for PDT: Past, present and future. Photodiagnosis Photodyn. Ther. 2004, 1, 43–48. [Google Scholar] [CrossRef]
- Brancaleon, L.; Moseley, H. Laser and non-laser light sources for photodynamic therapy. Lasers Med. Sci. 2002, 17, 173–186. [Google Scholar] [CrossRef]
- Moan, J. The Photochemical yield of singlet oxygen from porphyrins in different states of aggregation. Photochem. Photobiol. 1984, 39, 445–449. [Google Scholar] [CrossRef]
- Mang, T.S.; Dougherty, T.J.; Potter, W.R.; Boyle, D.G.; Somer, S.; Moan, J. Photobleaching of porphyrins used in photodynamic therapy and implications for therapy. Photochem. Photobiol. 1987, 45, 501–506. [Google Scholar] [CrossRef]
- Allison, R.R.; Downie, G.H.; Cuenca, R.; Hu, X.H.; Childs, C.J.; Sibata, C.H. Photosensitizers in clinical PDT. Photodiagnosis Photodyn. Ther. 2004, 1, 27–42. [Google Scholar] [CrossRef]
- Donnelly, R.F.; McCarron, P.A.; Tunney, M.M. Antifungal photodynamic therapy. Microbiol. Res. 2008, 163, 1–12. [Google Scholar] [CrossRef]
- Nakamura, S.; Mukai, T.; Senoh, M. Candela-class high-brightness InGaN/AlGaN double-heterostructure blue-light-emitting diodes. Appl. Phys. Lett. 1994, 64, 1687–1689. [Google Scholar] [CrossRef]
- Dai, T.; Gupta, A.; Murray, C.K.; Vrahas, M.S.; Tegos, G.P.; Hamblin, M.R. Blue light for infectious diseases: Propionibacterium acnes, Helicobacter pylori, and beyond? Drug Resist. Updat. 2012, 15, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Lipovsky, A.; Nitzan, Y.; Gedanken, A.; Lubart, R. Visible light-induced killing of bacteria as a function of wavelength: Im-plication for wound healing. Laser Surg. Med. 2010, 42, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Woolsey, G. Inactivation of bacterial pathogens following exposure to light from a 405-nanometer light-emitting diode array. Appl. Environ. Microbiol. 2009, 75, 1932–1937. [Google Scholar] [CrossRef]
- Liebmann, J.; Born, M.; Kolb-Bachofen, V. Blue-light irradiation regulates proliferation and differentiation in human skin cells. J. Investig. Dermatol. 2010, 130, 259–269. [Google Scholar] [CrossRef]
- Barolet, D. Light-emitting diodes (LEDs) in dermatology. Semin. Cutan. Med. Surg. 2008, 27, 227–238. [Google Scholar] [CrossRef]
- Ash, C.; Dubec, M.; Donne, K.; Bashford, T. Effect of wavelength and beam width on penetration in light-tissue interaction using computational methods. Lasers Med. Sci. 2017, 32, 1909–1918. [Google Scholar] [CrossRef]
- Jagdeo, J.; Austin, E.; Mamalis, A.; Wong, C.; Ho, D.; Siegel, D.M. Light-emitting diodes in dermatology: A systematic review of randomized controlled trials MS. Lasers Surg. Med. 2018, 50, 613–628. [Google Scholar] [CrossRef]
- Heelis, P.F. The photophysical and photochemical properties of flavins (isoalloxazines). Chem. Soc. Rev. 1982, 11, 15–39. [Google Scholar] [CrossRef]
- Ahmad, I.; Ahmed, S.; Sheraz, M.A.; Vaid, F.H.; Ansari, I.A. Effect of divalent anions on photodegradation kinetics and pathways of riboflavin in aqueous solution. Int. J. Pharm. 2010, 390, 174–182. [Google Scholar] [CrossRef]
- Insińska-Rak, M.; Golczak, A.; Sikorski, M. Photochemistry of riboflavin derivatives in methanolic solutions. J. Phys. Chem. A 2012, 116, 1199–1207. [Google Scholar] [CrossRef]
- Insińska-Rak, M.; Prukała, D.; Golczak, A.; Fornal, E.; Sikorski, M. Riboflavin degradation products; combined photochemical and mass spectrometry approach. J. Photochem. Photobiol. A Chem. 2020, 403, 112837. [Google Scholar] [CrossRef]
- Wolnicka-Glubisz, A.; Pawlak, A.; Insinska-Rak, M.; Zadlo, A. Analysis of photoreactivity and phototoxicity of riboflavin’s analogue 3MeTARF. J. Photochem. Photobiol. B Biol. 2020, 205, 111820. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Ahmed, S.; Anwar, Z.; Sheraz, M.A.; Sikorski, M. Photostability and photostabilization of drugs and drug products. Int. J. Photoenergy 2016, 2016, 1–19. [Google Scholar] [CrossRef]
- Ribes, J.; Cossard, P.; Al Yaman, K.; Bestel, I.; Badarau, E. Investigating the photosensitization activities of flavins irradiated by blue LEDs. RSC Adv. 2023, 13, 2355–2364. [Google Scholar] [CrossRef]
- Moore, W.M.; Baylor, J. The photochemistry of riboflavin IV. The photobleaching of some nitrogen-9 substituted isoalloxazines and flavins. J. Am. Chem. Soc. 1969, 91, 7170–7179. [Google Scholar] [CrossRef]
- Jorns, M.S.; Schollnhammer, G.; Hemmerich, P. Intramolecular addition of the riboflavin side chain. Anion-catalyzed neutral photochemistry. JBIC J. Biol. Inorg. Chem. 1975, 57, 35–48. [Google Scholar] [CrossRef]
- Larson, R.A.; Stackhouse, P.L.; Crowley, T.O. Riboflavin tetraacetate: A potentially useful photosensitizing agent for the treatment of contaminated waters. Environ. Sci. Technol. 1992, 26, 1792–1798. [Google Scholar] [CrossRef]
- Remucal, C.K.; McNeill, K. Photosensitized amino acid degradation in the presence of riboflavin and its derivatives. Environ. Sci. Technol. 2011, 45, 5230–5237. [Google Scholar] [CrossRef]
- Banekovich, C.; Matuszczak, B. 2′,3′,4′,5′-Tetraacetyl-N(3)-carboxymethylriboflavin derivatives: Synthesis and fluorescence studies. Tetrahedron Lett. 2005, 46, 5053–5056. [Google Scholar] [CrossRef]
- Sikorska, E.; Khmelinskii, I.; Komasa, A.; Koput, J.; Ferreira, L.F.; Herance, J.R.; Bourdelande, J.L.; Williams, S.L.; Worrall, D.R.; Insińska-Rak, M.; et al. Spectroscopy and photophysics of flavin related compounds: Riboflavin and iso-(6,7)-riboflavin. Chem. Phys. 2005, 314, 239–247. [Google Scholar] [CrossRef]
- Silva, A.V.; López-Sánchez, A.; Junqueira, H.C.; Rivas, L.; Baptista, M.S.; Orellana, G. Riboflavin derivatives for enhanced photodynamic activity against Leishmania parasites. Tetrahedron 2015, 71, 457–462. [Google Scholar] [CrossRef]
- Insińska-Rak, M.; Sikorska, E.; Bourdelande, J.L.; Khmelinskii, I.V.; Prukała, W.; Dobek, K.; Karolczak, J.; Machado, I.F.; Fer-reira, L.F.V.; Dulewicz, E. New photochemically stable riboflavin analogue-3-methyl-riboflavin tetraacetate. J. Photochem. Photobiol. A 2007, 186, 14–23. [Google Scholar] [CrossRef]
- Lafaye, C.; Aumonier, S.; Torra, J.; Signor, L.; von Stetten, D.; Noirclerc-Savoye, M.; Shu, X.; Ruiz González, R.; Gotthard, G.; Royant, A.; et al. Ribofavin binding proteins for singlet oxygen production. Photochem. Photobiol. Sci. 2022, 21, 1545–1555. [Google Scholar] [CrossRef]
- Mogensen, D.J.; Etzerodt, M.; Ogilby, P.R. Photoinduced bleaching in an efficient singlet oxygen photosensitizing protein: Identifying a culprit in the flavin-binding LOV-based protein SOPP3. J. Photochem. Photobiol. A Chem. 2022, 429, 113894. [Google Scholar] [CrossRef]
- Westberg, M.; Brenghoj, M.; Etzerodt, M.; Ogilby, P.R. No photon wasted: An efficient and selective singlet oxygen photosen-sitizing protein. J. Phys. Chem. B 2017, 121, 9366–9371. [Google Scholar] [CrossRef]
- Insińska-Rak, M.; Sikorska, E.; Bourdelande, J.L.; Khmelinskii, I.V.; Prukała, W.; Dobek, K.; Karolczak, J.; Machado, I.F.; Ferreira, L.F.; Komasa, A.; et al. Spectroscopy and photophysics of flavin-related compounds: 5-deaza-riboflavin. J. Mol. Struct. 2005, 783, 184–190. [Google Scholar] [CrossRef]
- Sikorska, E.; Khmelinskii, I.V.; Prukała, W.; Williams, S.L.; Patel, M.; Worrall, D.R.; Bourdelande, J.L.; Koput, J.; Sikorski, M. Spectroscopy and photophysics of lumiflavins and lumichromes. J. Phys. Chem. A 2004, 108, 1501–1508. [Google Scholar] [CrossRef]
- Zirak, P.; Penzkofer, A.; Mathes, T.; Hegemann, P. Absorption and emission spectroscopic characterization of BLUF protein Slr1694 from Synechocystis sp. PCC6803 with roseoflavin cofactor. J. Photochem. Photobiol. B Biol. 2009, 97, 61–70. [Google Scholar] [CrossRef]
- Chacon, J.N.; McLearie, J.; Sinclair, R.S. Singlet oxygen yields and radical contributions in the dye- sensitized photo-oxidation in methanol of esters of poly- unsaturated fatty-acids (oleic, linoleic, linolenic and archidonic). Photochem. Photobiol. 1988, 47, 647–656. [Google Scholar] [CrossRef]
- Moore, W.M.; McDaniels, J.C.; Hen, J.A. The photochemistry of riboflavin—VI. The photophysical properties of isoalloxazines. Photochem. Photobiol. 1977, 25, 505–512. [Google Scholar] [CrossRef]
- Holzer, W.; Penzkofer, A.; Hegemann, P. Absorption and emission spectroscopic characterisation of the LOV2-His domain of phot from Chlamydomonas reinhardtii. Chem. Phys. 2005, 308, 79–91. [Google Scholar] [CrossRef]
- Bertolotti, S.G.; Previtali, C.M.; Rufs, A.M.; Encinas, M.V. Riboflavin triethanolamine as photoinitiator system of vinyl polymerization. A mechanistic study by laser flash photolysis. Macromolecules 1999, 32, 2920–2924. [Google Scholar] [CrossRef]
- Sikorski, M.; Sikorska, E.; Koziolowa, A.; Moreno, R.G.; Bourdelande, J.; Steer, R.; Wilkinson, F. Photophysical properties of lumichromes in water. J. Photochem. Photobiol. B Biol. 2001, 60, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Suwannasom, N.; Kao, I.; Pruß, A.; Georgieva, R.; Bäumler, H. Riboflavin: The health benefits of a forgotten natural vitamin. Int. J. Mol. Sci. 2020, 21, 950. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Singh, P.K.; Srivastava, A.; Singh, P.P. Synthetic applications of flavin photocatalysis: A review. RSC Adv. 2021, 11, 14251–14259. [Google Scholar] [CrossRef]
- Lindén, A.A.; Krüger, L.; Bäckvall, J.-E. Highly selective sulfoxidation of allylic and vinylic sulfides by hydrogen peroxide using a flavin as catalyst. J. Org. Chem. 2003, 68, 5890–5896. [Google Scholar] [CrossRef]
- Cui, H.; Hwang, H.-M.; Cook, S.; Zeng, K. Effect of photosensitizer riboflavin on the fate of 2,4,6-trinitrotoluene in a freshwater environment. Chemosphere 2001, 44, 621–625. [Google Scholar] [CrossRef]
- Lu, C.Y.; Han, Z.H.; Liu, G.S.; Cai, X.C.; Chen, Y.L.; Yao, S. Photophysical and photochemical processes of riboflavin (vitamin B2) by means of the transient absorption spectra in aqueous solution. Sci. China Ser. B Chem. 2001, 44, 39–48. [Google Scholar] [CrossRef]
- Lu, C.-Y.; Lin, W.-Z.; Wang, W.-F.; Han, Z.-H.; Zheng, Z.-D.; Yao, S.-D. Kinetic observation of rapid electron transfer between pyrimidine electron adducts and sensitizers of riboflavin, flavin adenine dinucleotide (FAD) and chloranil: A pulse radiolysis study. Radiat. Phys. Chem. 2000, 59, 61–66. [Google Scholar] [CrossRef]
- Lu, C.-Y.; Wang, W.-F.; Lin, W.-Z.; Han, Z.-H.; Yao, S.-D.; Lin, N.-Y. Generation and photosensitization properties of the oxidized radical of riboflavin: A laser flash photolysis study. J. Photochem. Photobiol. B Biol. 1999, 52, 111–116. [Google Scholar] [CrossRef]
- Cardoso, D.R.; Libardi, S.H.; Skibsted, L.H. Riboflavin as a photosensitizer. Effects on human health and food quality. Food Funct. 2012, 3, 487–502. [Google Scholar] [CrossRef] [PubMed]
- King, J.M.; Min, D.B. Riboflavin-photosensitized singlet oxygen oxidation product of vitamin D2. J. Am. Oil Chem. Soc. 2002, 79, 983–987. [Google Scholar] [CrossRef]
- King, J.M.; Min, D.B. Riboflavin photosensitized singlet oxygen oxidation of vitamin D. J. Food Sci. 1998, 63, 31–34. [Google Scholar] [CrossRef]
- Grininger, M.; Staudt, H.; Johansson, P.; Wachtveitl, J.; Oesterhelt, D. Dodecin is the key player in flavin homeostasis of archaea. Pediatrics 2009, 284, 13068–13076. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, M.; Khmelinskii, I.; Sikorska, E. Spectral Properties of flavins. In Flavin-Based Catalysis; Cibulka, R., Fraaije, M., Eds.; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2021; pp. 67–96. [Google Scholar]
- Climent, T.; Gonzalez-Luque, R.; Merchan, M.; Serrano-Andres, L. Theoretical insight into the spectroscopy and photochem-istry of isoalloxazine, the flavin core ring. J. Phys. Chem. A 2006, 110, 13584–13590. [Google Scholar] [CrossRef]
- Prukała, D.; Gierszewski, M.; Pędziński, T.; Sikorski, M. Influence of pH on spectral and photophysical properties of 9-methyl-5-deazaalloxazine and 10-ethyl-5-deaza-isoalloxazine. J. Photochem. Photobiol. A Chem. 2014, 275, 12–20. [Google Scholar] [CrossRef]
- Senda, T.; Senda, M.; Kimura, S.; Ishida, T. Redox control of protein conformation in flavoproteins. Antioxid. Redox Signal. 2009, 11, 1741–1766. [Google Scholar] [CrossRef]
- Kao, Y.-T.; Saxena, C.; He, T.-F.; Guo, L.; Wang, L.; Sancar, A.; Zhong, D. Ultrafast dynamics of flavins in five redox states. J. Am. Chem. Soc. 2008, 130, 13132–13139. [Google Scholar] [CrossRef]
- McBride, R.A.; Barnard, D.T.; Jacoby-Morris, K.; Harun-Or-Rashid, M.; Stanley, J. Reduced flavin in aqueous solution is non-fluorescent. Biochemistry 2023, 62, 759–769. [Google Scholar] [CrossRef]
- Visser, A.J.W.G.; Ghisla, S.; Massey, V.; Muller, F.; Veeger, C. Fluorescence properties of reduced flavins and flavoproteins. JBIC J. Biol. Inorg. Chem. 1979, 101, 13–21. [Google Scholar] [CrossRef]
- Zhuang, B.; Aleksandrov, A.; Seo, D.; Vos, M.H. Excited-state properties of fully reduced flavins in ferredoxin–NADP+ oxi-doreductase. J. Phys. Chem. Lett. 2023, 14, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-Y.; Chang, K.-C.; Chen, L.-Y.; Hu, A. Low-dose blue light irradiation enhances the antimicrobial activities of cur-cumin against Propionibacterium acnes. J. Photochem. Photobiol. B 2018, 189, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Wolnicka-Glubisz, A.; Wisniewska-Becker, A. Dual action of curcumin as an anti- and pro-oxidant from a biophysical perspective. Antioxidants 2023, 12, 1725. [Google Scholar] [CrossRef]
- Zheng, Y.; Yu, E.; Weng, Q.; Zhou, L.; Li, Q. Optimization of hydrogel containing toluidine blue O for photodynamic therapy in treating acne. Lasers Med. Sci. 2019, 34, 1535–1545. [Google Scholar] [CrossRef]
- Wangsuwan, S.; Meephansan, J. Comparative study of photodynamic therapy with riboflavin-tryptophan Gel and 13% 5-Aminolevulinic acid in the treatment of mild to moderate acne vulgaris. Clin. Cosmet. Investig. Dermatol. 2019, 12, 805–814. [Google Scholar] [CrossRef]
- Corbin, F. Pathogen inactivation of blood components: Current status and introduction of an approach using riboflavin as a photosensitizer. Int. J. Hematol. 2002, 76, 253–257. [Google Scholar] [CrossRef]
- Reddy, H.L.; Dayan, A.D.; Cavagnaro, J.; Gad, S.; Li, J.; Goodrich, R.P. Toxicity testing of a novel riboflavin-based technology for pathogen reduction and white blood cell inactivation. Transfus. Med. Rev. 2008, 22, 133–153. [Google Scholar] [CrossRef]
- Balint, B.; Jovicic-Gojkov, D.; Todorovic-Balint, M.; Subota, V.; Pavlovic, M.; Goodrich, R. Plasma constituent integrity in pre-storage vs. post-storage riboflavin and UV-light treatment—A comparative study. Transf. Apher. Sci. 2013, 49, 434–443. [Google Scholar] [CrossRef]
- Hornsey, V.S.; Drummond, O.; Morrison, A.; McMillan, L.; MacGregor, I.R.; Prowse, C.V. Pathogen reduction of fresh plasma using riboflavin and ultraviolet light: Effects on plasma coagulation proteins. Transfusion 2009, 49, 2167–2172. [Google Scholar] [CrossRef]
- Smith, J.; Rock, G. Protein quality in Mirasol pathogen reduction technology–treated, apheresis-derived fresh-frozen plasma. Transfusion 2010, 50, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Farah, N.; Chin, V.K.; Chong, P.P.; Lim, W.F.; Lim, C.W.; Basir, R.; Chang, S.K.; Lee, T.Y. Riboflavin as a promising antimicrobial agent? A multi-perspective review. Curr. Res. Microb. Sci. 2022, 3, 100111. [Google Scholar] [CrossRef] [PubMed]
- Buninowska, I.; Wroński, N.; Insińska-Rak, M.; Sikorski, M.; Wolnicka-Glubisz, A. Tetraacetyl riboflavin derivative mediates caspase 3/7 activation via MAPK in A431 cells upon blue light influence. Photochem. Photobiol. 2023. [Google Scholar] [CrossRef]
- Akasov, R.A.; Sholina, N.V.; Khochenkov, D.A.; Alova, A.; Gorelkin, P.V.; Erofeev, A.S.; Generalova, A.N.; Khaydukov, E.V. Photodynamic therapy of melanoma by blue-light photoactivation of flavin mononucleotide. Sci. Rep. 2019, 9, 9679. [Google Scholar] [CrossRef]
- Bergh, V.J.V.; Bruzell, E.; Hegge, A.B.; Tønnesen, H.H. Influence of formulation on photoinactivation of bacteria by lumichrome. Die Pharm. 2015, 70. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, L.; He, C.; Tai, S.; Zhu, L.; Ma, C.; Yang, T.; Cheng, F.; Sun, X.; Cui, R.; et al. An experimental study on riboflavin photosensitization treatment for inactivation of circulating HCT116 tumor cells. J. Photochem. Photobiol. B 2019, 196, 111496. [Google Scholar] [CrossRef]
- Salman, M.; Naseem, I. Riboflavin as adjuvant with cisplatin: Study in mouse skin cancer model. Front. Biosci. 2015, 7, 242–254. [Google Scholar] [CrossRef]
- Yang, M.-Y.; Chang, C.-J.; Chen, L.-Y. Blue light induced reactive oxygen species from flavin mononucleotide and flavin ad-enine dinucleotide on lethality of HeLa cells. J. Photochem. Photobiol. B 2017, 173, 325–332. [Google Scholar] [CrossRef]
- Davis, L.E.; Shalin, S.C.; Tackett, A.J. Current state of melanoma diagnosis and treatment. Cancer Biol. Ther. 2019, 20, 1366–1379. [Google Scholar] [CrossRef]
- Cives, M.; Mannavola, F.; Lospalluti, L.; Sergi, M.C.; Cazzato, G.; Filoni, E.; Cavallo, F.; Giudice, G.; Stucci, L.S.; Porta, C.; et al. Non-melanoma skin cancers: Biological and clinical features. Int. J. Mol. Sci. 2020, 21, 5394. [Google Scholar] [CrossRef]
- Juarez, A.V.; Sosa, L.d.V.; De Paul, A.L.; Costa, A.P.; Farina, M.; Leal, R.B.; Torres, A.I.; Pons, P. Riboflavin acetate induces apoptosis in squamous carcinoma cells after photodynamic therapy. J. Photochem. Photobiol. B Biol. 2015, 153, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Borenstein, B. CRC Handbook of Food Additives; Borenstein, B., Ed.; The Chemical Rubber Co.: Boca Raton, FL, USA; Washington, DC, USA, 1972; pp. 85–111. [Google Scholar]
- Bartmann, L.; Schumacher, D.; von Stillfried, S.; Sternkopf, M.; Alampour-Rajabi, S.; van Zandvoort, M.A.M.J.; Kiessling, F.; Wu, Z. Evaluation of riboflavin transporters as targets for drug delivery and theranostics. Front. Pharmacol. 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Peechakara, B.V.; Gupta, M. Vitamin B2 (Riboflavin). In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023, bookshelf ID: NBK525977. National Library of Medicine. Available online: https://www.ncbi.nlm.nih.gov/books/NBK525977/ (accessed on 28 August 2023).
- Muñoz, M.A.; Pacheco, A.; Becker, M.I.; Silva, E.; Ebensperger, R.; Garcia, A.M.; De Ioannes, A.E.; Edwards, A.M. Different cell death mechanisms are induced by a hydrophobic flavin in human tumor cells after visible light irradiation. J. Photochem. Photobiol. B Biol. 2011, 103, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Crocker, L.B.; Lee, J.H.; Mital, S.; Mills, G.C.; Schack, S.; Bistrović-Popov, A.; Franck, C.O.; Mela, I.; Kaminski, C.F.; Christie, G.; et al. Tuning riboflavin derivatives for photodynamic inactivation of pathogens. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Arbiser, J.L.; Bonner, M.Y.; Gilbert, L.C. Targeting the duality of cancer. Npj Precis. Oncol. 2017, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Lossi, L. The concept of intrinsic versus extrinsic apoptosis. Biochem. J. 2022, 479, 357–384. [Google Scholar] [CrossRef]
- Hu, H.; Xu, F.; Yang, W.; Ren, J.; Ge, W.; Yang, P. Apoptosis as an underlying mechanism in lymphocytes induced by riboflavin and ultraviolet light. Transfus. Apher. Sci. 2020, 59, 102899. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef]
- Kirkin, V.; Joos, S.; Zörnig, M. The role of Bcl-2 family members in tumorigenesis. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2004, 1644, 229–249. [Google Scholar] [CrossRef]
- Ahmad, I.; Fasiliullah, Q.; Vaid, F.H.M. A study of simultaneous photolysis and photoaddition reactions of riboflavin in aqueous solution. J. Photochem. Photobiol. B 2004, 75, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Niemz, A.; Imbriglio, J.; Rotello, V.M. Model systems for flavoenzyme activity: One- and two-electron reduction of flavins in aprotic hydrophobic environments. J. Am. Chem. Soc. 1997, 119, 887–892. [Google Scholar] [CrossRef]
- Grajek, H.; Żurkowska, G.; Kuśba, J. Influence of diffusion on nonradiative energy transfer between FMN molecules in aqueous solutions. J. Photochem. Photobiol. B Biol. 2005, 80, 145–155. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sensitizer | λA [nm] | λF [nm] | ΦF | τF [ns] | ΦISC | ΦΔ |
|---|---|---|---|---|---|---|
| RiboflavinMeOH [a] | 444 | 532 | 0.39 | 6.4/5.4 [k] | 0.6 [l] | 0.51 |
| Riboflavinwater [b] | 444 | 537 | 0.28 | 5.1 | 0.7 [m] | 0.48 [l] |
| TARFMeOH [c] | 446 | 525 | 0.46 | - | - | - |
| TARFwater [b] | 446 | - | - | - | 0.66 | |
| 3MeTARFMeOH [d] | 448 | 513 | 0.089/0.12 [c] | 5.4 | - | 0.61/0.46 [e] |
| 3MeTARFwater [d] | 451 | 520 | 0.11 | 4.4 | - | - |
| 3MeTARFPBS/DMSO [e] | 450 | - | - | - | - | 0.49 |
| FMNTRIS(d) [f] | - | - | - | - | - | 0.57 |
| FMNwater [g] | - | - | 0.23 [n] | 5.1 [n] | - | 0.59 |
| miniSOG [FMN]TRIS(d) [f] | - | - | 0.4 | 5.5 | 0.6 | 0.04 |
| miniSOG [Rfl]TRIS(d) [f] | - | - | - | - | - | 0.10 |
| SOPP3 [h] | - | - | - | - | - | 0.61D2O |
| Iso-6,7-riboflavinMeOH [a] | 447 | 552 | 0.20 | 4.2 | - | 0.70 |
| 5-deazariboflavinMeOH [i] | 400 | 455 | 0.11 | 3.98 | - | 0.33 |
| LumichromeMeOH [j] | 384 | 453 | 0.032 | 1.04 [o] | - | 0.85 |
| Lumichromewater [j] | 385 | 479 | 0.088 | 2.7 | 0.63 [b]/0.69 [p] | 0.36D2O [p] |
| [Ref.]: [a] [42]; [b] [40]; [c] [43]; [d] [44]; [e] [34]; [f] [45]; [g] [46]; [h] [47]; [i] [48]; [j] [49]; [k] [50]; [l] [51]; [m] [52]; [n] [53]; [o] [54]; [p] [55]. | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Insińska-Rak, M.; Sikorski, M.; Wolnicka-Glubisz, A. Riboflavin and Its Derivates as Potential Photosensitizers in the Photodynamic Treatment of Skin Cancers. Cells 2023, 12, 2304. https://doi.org/10.3390/cells12182304
Insińska-Rak M, Sikorski M, Wolnicka-Glubisz A. Riboflavin and Its Derivates as Potential Photosensitizers in the Photodynamic Treatment of Skin Cancers. Cells. 2023; 12(18):2304. https://doi.org/10.3390/cells12182304
Chicago/Turabian StyleInsińska-Rak, Małgorzata, Marek Sikorski, and Agnieszka Wolnicka-Glubisz. 2023. "Riboflavin and Its Derivates as Potential Photosensitizers in the Photodynamic Treatment of Skin Cancers" Cells 12, no. 18: 2304. https://doi.org/10.3390/cells12182304