
Role of Chemokine Cxcl12a in Mediating the Stimulatory Effects of Ethanol on Embryonic Development of Subpopulations of Hypocretin/Orexin Neurons and Their Projections

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Embryonic Ethanol Treatment
2.3. cxcl12a mRNA Synthesis and Injection
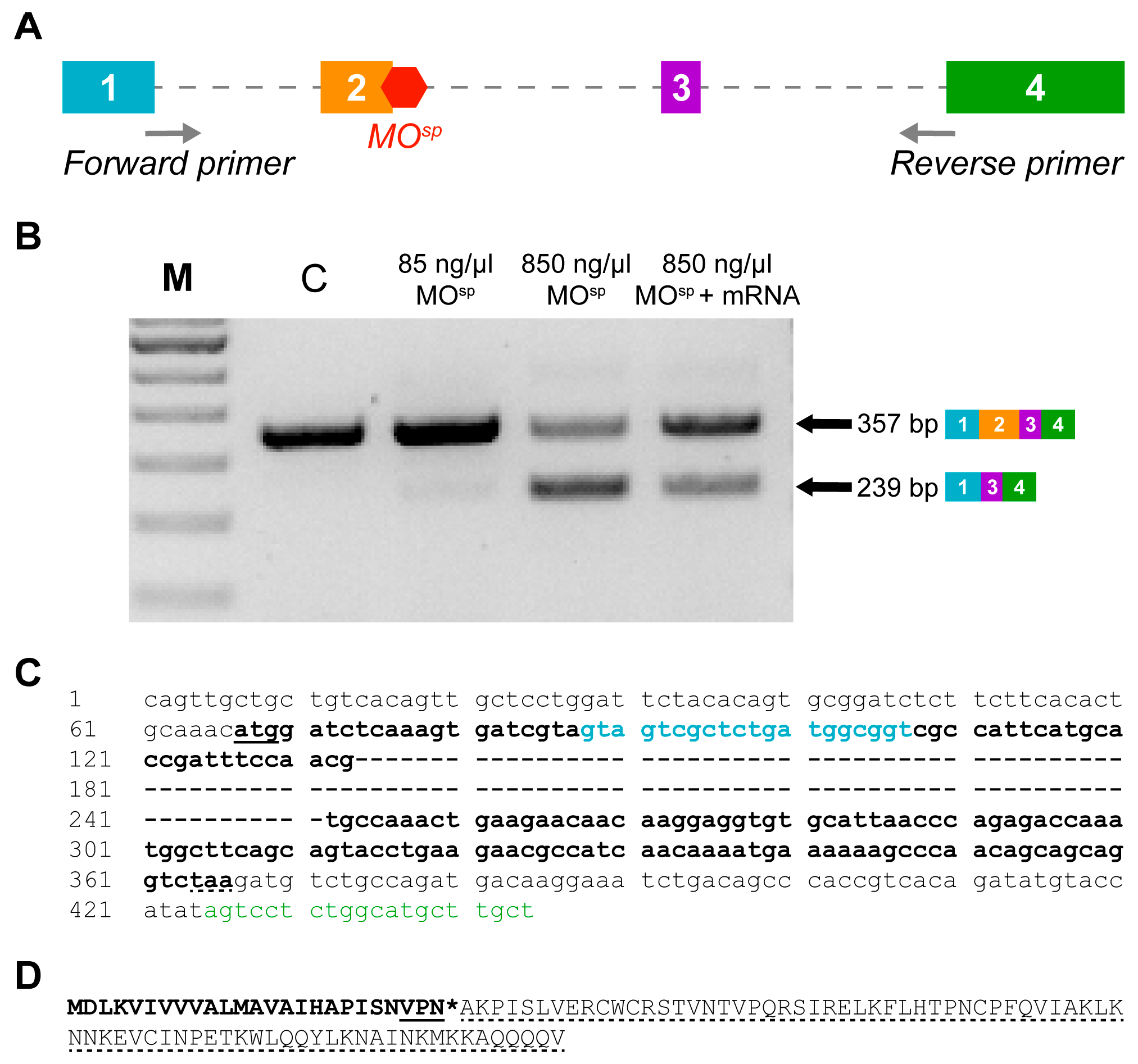
2.4. cxcl12a Morpholino Design and Injection
2.5. RNA Extraction, cDNA Synthesis and RT-PCR
2.6. Microscopy and Image Analysis
2.7. Statistical Analyses
3. Results
3.1. Confirmation of cxcl12a mRNA and MOsp
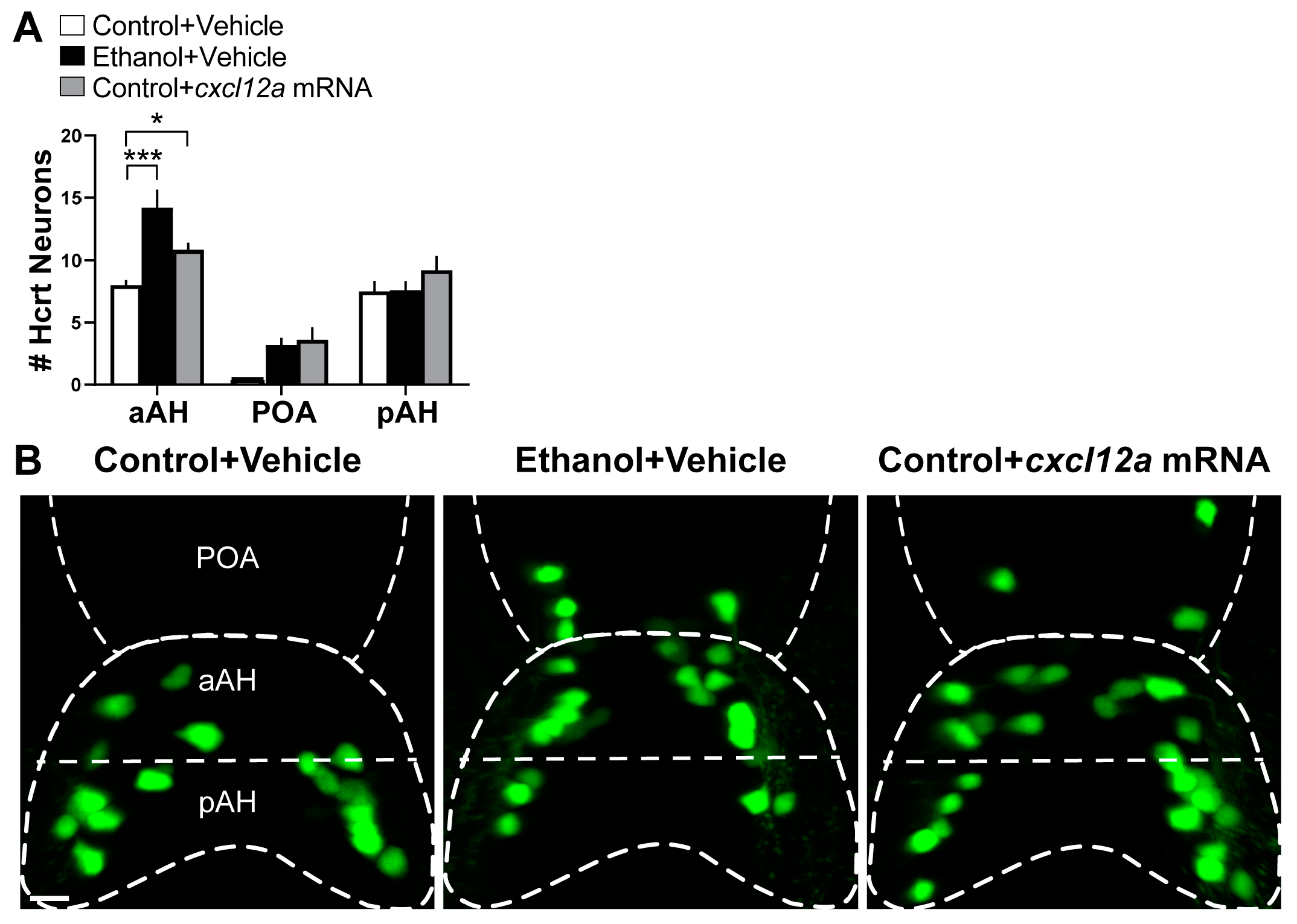
3.2. Cxcl12a Overexpression Mimics the Ethanol-Induced Increase in Number of Hcrt Neurons in the aAH and POA with No Effect in the pAH
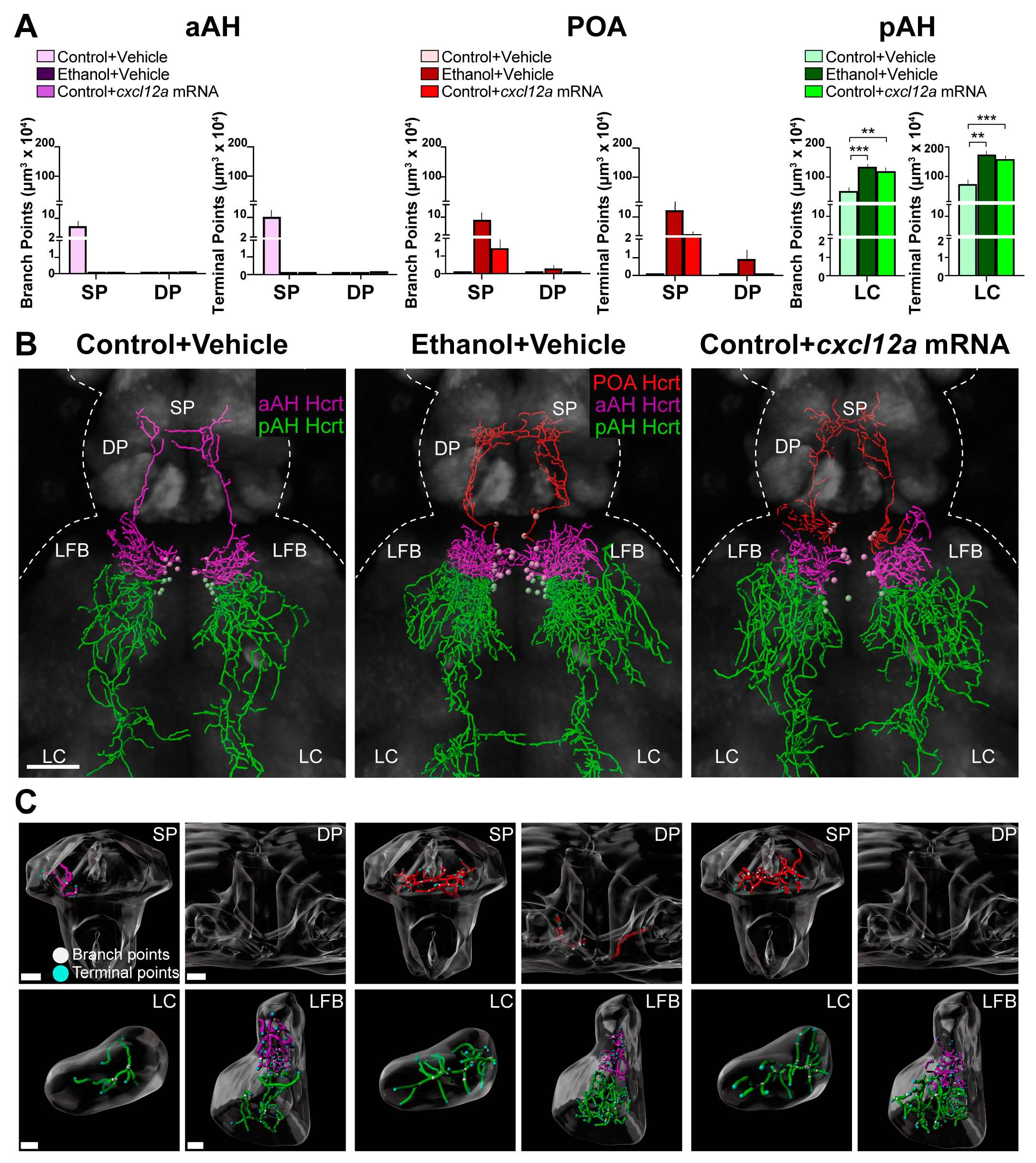
3.3. Cxcl12a Overexpression Mimics the Ethanol-Induced Stimulation of Hcrt Projections to Distant Areas in the Anterior and Posterior Brain
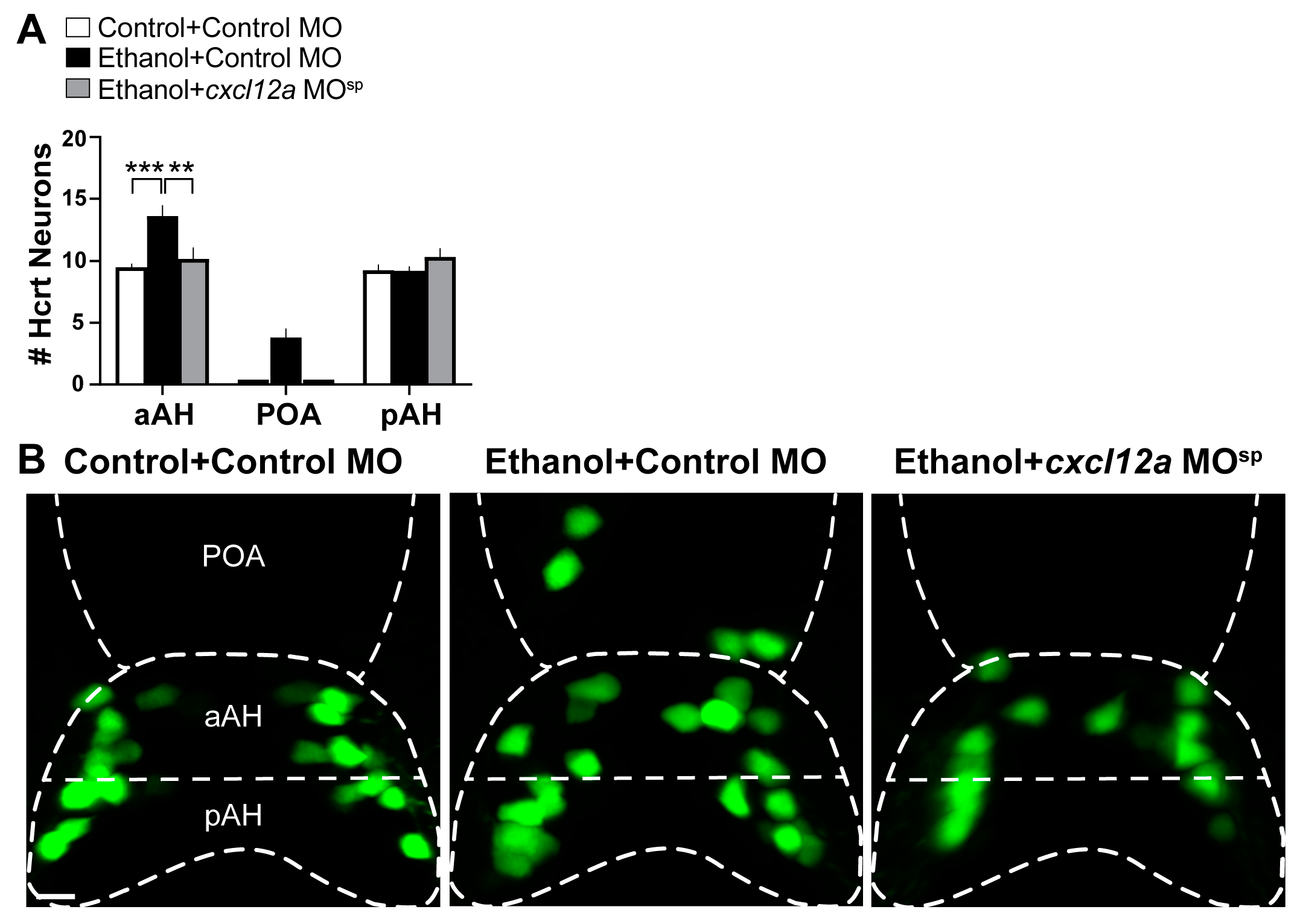
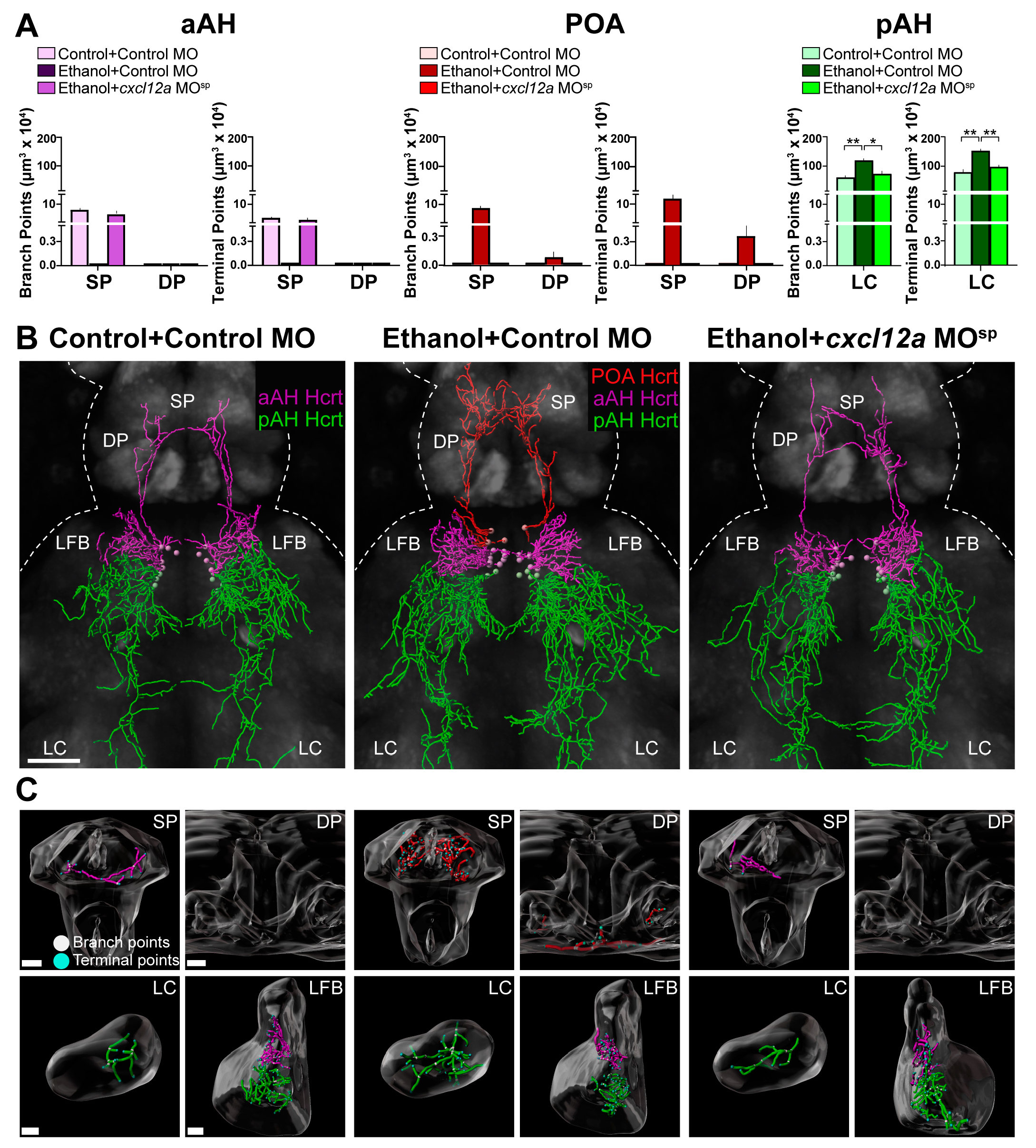
3.4. Cxcl12a Knockdown Blocks the Ethanol-Induced increase in Number of Hcrt Neurons in the aAH and POA
3.5. Cxcl12a Knockdown Blocks the Ethanol-Induced Stimulation of Hcrt Projections to Distant Areas in the Anterior and Posterior Brain
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, K.K.; Gupta, V.K.; Shirasaka, T. An Update on Fetal Alcohol Syndrome-Pathogenesis, Risks, and Treatment. Alcohol. Clin. Exp. Res. 2016, 40, 1594–1602. [Google Scholar] [CrossRef]
- Petrelli, B.; Weinberg, J.; Hicks, G.G. Effects of prenatal alcohol exposure (PAE): Insights into FASD using mouse models of PAE. Biochem. Cell Biol. 2018, 96, 131–147. [Google Scholar] [CrossRef]
- Kleiber, M.L.; Mantha, K.; Stringer, R.L.; Singh, S.M. Neurodevelopmental alcohol exposure elicits long-term changes to gene expression that alter distinct molecular pathways dependent on timing of exposure. J. Neurodev. Disord. 2013, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Cuzon, V.C.; Yeh, P.W.L.; Yanagawa, Y.; Obata, K.; Yeh, H.H. Ethanol Consumption during Early Pregnancy Alters the Disposition of Tangentially Migrating GABAergic Interneurons in the Fetal Cortex. J. Neurosci. 2008, 28, 1854–1864. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.-Q.; Karatayev, O.; Liang, S.C.; Barson, J.R.; Leibowitz, S.F. Prenatal ethanol exposure stimulates neurogenesis in hypothalamic and limbic peptide systems: Possible mechanism for offspring ethanol overconsumption. Neuroscience 2012, 222, 417–428. [Google Scholar] [CrossRef]
- Chang, G.-Q.; Karatayev, O.; Leibowitz, S.F. Prenatal exposure to ethanol stimulates hypothalamic CCR2 chemokine receptor system: Possible relation to increased density of orexigenic peptide neurons and ethanol drinking in adolescent offspring. Neuroscience 2015, 310, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.D.; Yasmin, N.; Chang, G.Q.; Karatayev, O.; Khalizova, N.; Fam, M.; Abdulai, A.R.; Yu, B.; Leibowitz, S.F. Embryonic ethanol exposure induces ectopic Hcrt and MCH neurons outside hypothalamus in rats and zebrafish: Role in ethanol-induced behavioural disturbances. Addict. Biol. 2022, 27, e13238. [Google Scholar] [CrossRef]
- Lovely, C.B.; Fernandes, Y.; Eberhart, J.K. Fishing for Fetal Alcohol Spectrum Disorders: Zebrafish as a Model for Ethanol Teratogenesis. Zebrafish 2016, 13, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, N.; Collier, A.D.; Karatayev, O.; Abdulai, A.R.; Yu, B.; Fam, M.; Khalizova, N.; Leibowitz, S.F. Subpopulations of hypocretin/orexin neurons differ in measures of their cell proliferation, dynorphin co-expression, projections, and response to embryonic ethanol exposure. Sci. Rep. 2023. in review. [Google Scholar]
- Collier, A.D.; Yasmin, N.; Karatayev, O.; Abdulai, A.R.; Yu, B.; Khalizova, N.; Fam, M.; Leibowitz, S.F. Neuronal chemokine concentration gradients mediate effects of embryonic ethanol exposure on ectopic hypocretin/orexin neurons and behavior in zebrafish. Sci. Rep. 2023, 13, 1447. [Google Scholar] [CrossRef]
- Collier, A.D.; Halkina, V.; Min, S.S.; Roberts, M.Y.; Campbell, S.D.; Camidge, K.; Leibowitz, S.F. Embryonic Ethanol Exposure Affects the Early Development, Migration, and Location of Hypocretin/Orexin Neurons in Zebrafish. Alcohol. Clin. Exp. Res. 2019, 43, 1702–1713. [Google Scholar] [CrossRef] [PubMed]
- Boric, K.; Orio, P.; Viéville, T.; Whitlock, K. Quantitative Analysis of Cell Migration Using Optical Flow. PLoS ONE 2013, 8, e69574. [Google Scholar] [CrossRef]
- Coffey, C.M.; Solleveld, P.A.; Fang, J.; Roberts, A.K.; Hong, S.-K.; Dawid, I.B.; Laverriere, C.E.; Glasgow, E. Novel Oxytocin Gene Expression in the Hindbrain Is Induced by Alcohol Exposure: Transgenic Zebrafish Enable Visualization of Sensitive Neurons. PLoS ONE 2013, 8, e53991. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.J.; Zhang, C.; Ojiaku, P.; Bell, V.; Devkota, S.; Mukhopadhyay, S. Effects of Ethanol Exposure on Nervous System Development in Zebrafish. Int. Rev. Cell Mol. Biol. 2012, 299, 255–315. [Google Scholar] [CrossRef] [PubMed]
- Chevassus-Au-Louis, N.; Represa, A. The right neuron at the wrong place: Biology of heterotopic neurons in cortical neuronal migration disorders, with special reference to associated pathologies. Cell. Mol. Life Sci. 1999, 55, 1206–1215. [Google Scholar] [CrossRef]
- Wozniak, J.R.; Riley, E.P.; Charness, M.E. Clinical presentation, diagnosis, and management of fetal alcohol spectrum disorder. Lancet Neurol. 2019, 18, 760–770. [Google Scholar] [CrossRef]
- Dunty, W.C., Jr.; Zucker, R.M.; Sulik, K.K. Hindbrain and cranial nerve dysmorphogenesis result from acute maternal ethanol administration. Dev. Neurosci. 2002, 24, 328–342. [Google Scholar] [CrossRef]
- Abbott, C.W.; Rohac, D.J.; Bottom, R.T.; Patadia, S.; Huffman, K.J. Prenatal Ethanol Exposure and Neocortical Development: A Transgenerational Model of FASD. Cereb. Cortex 2018, 28, 2908–2921. [Google Scholar] [CrossRef] [PubMed]
- Goeke, C.M.; Roberts, M.L.; Hashimoto, J.G.; Finn, D.A.; Guizzetti, M. Neonatal Ethanol and Choline Treatments Alter the Morphology of Developing Rat Hippocampal Pyramidal Neurons in Opposite Directions. Neuroscience 2018, 374, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, G.F.; Whitcher, L.T.; Klintsova, A. Postnatal binge-like alcohol exposure decreases dendritic complexity while increasing the density of mature spines in mPFC Layer II/III pyramidal neurons. Synapse 2010, 64, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, G.F.; Criss, K.J.; Klintsova, A.Y. Voluntary exercise partially reverses neonatal alcohol-induced deficits in mPFC layer II/III dendritic morphology of male adolescent rats. Synapse 2015, 69, 405–415. [Google Scholar] [CrossRef]
- Collier, A.D.; Khalizova, N.; Chang, G.Q.; Min, S.; Campbell, S.; Gulati, G.; Leibowitz, S.F. Involvement of Cxcl12a/Cxcr4b Chemokine System in Mediating the Stimulatory Effect of Embryonic Ethanol Exposure on Neuronal Density in Zebrafish Hypothalamus. Alcohol. Clin. Exp. Res. 2020, 44, 2519–2535. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Collier, A.D.; Karatayev, O.; Gulati, G.; Boorgu, D.; Leibowitz, S.F. Moderate Prenatal Ethanol Exposure Stimulates CXCL12/CXCR4 Chemokine System in Radial Glia Progenitor Cells in Hypothalamic Neuroepithelium and Peptide Neurons in Lateral Hypothalamus of the Embryo and Postnatal Offspring. Alcohol. Clin. Exp. Research. 2020, 44, 866–879. [Google Scholar] [CrossRef]
- Watson, A.E.S.; Goodkey, K.; Footz, T.; Voronova, A. Regulation of CNS precursor function by neuronal chemokines. Neurosci. Lett. 2020, 715, 134533. [Google Scholar] [CrossRef]
- Poon, K.; Barson, J.R.; Shi, H.; Chang, G.Q.; Leibowitz, S.F. Involvement of the CXCL12 System in the Stimulatory Effects of Prenatal Exposure to High-Fat Diet on Hypothalamic Orexigenic Peptides and Behavior in Offspring. Front. Behav. Neurosci. 2017, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Chédotal, A. Roles of axon guidance molecules in neuronal wiring in the developing spinal cord. Nat. Rev. Neurosci. 2019, 20, 380–396. [Google Scholar] [CrossRef]
- Festa, L.K.; Irollo, E.; Platt, B.J.; Tian, Y.; Floresco, S.; Meucci, O. CXCL12-induced rescue of cortical dendritic spines and cognitive flexibility. Elife 2020, 9, e49717. [Google Scholar] [CrossRef]
- Palevitch, O.; Abraham, E.; Borodovsky, N.; Levkowitz, G.; Zohar, Y.; Gothilf, Y. Cxcl12a–Cxcr4b signaling is important for proper development of the forebrain GnRH system in zebrafish. Gen. Comp. Endocrinol. 2010, 165, 262–268. [Google Scholar] [CrossRef]
- Li, J.; Wu, Y.; Chen, P.; Huang, X.; Liu, Y.; Peng, M.; Wu, R.-Q. CXCL12 promotes spinal nerve regeneration and functional recovery after spinal cord injury. Neuroreport 2021, 32, 450–457. [Google Scholar] [CrossRef]
- Appelbaum, L.; Wang, G.X.; Maro, G.S.; Mori, R.; Tovin, A.; Marin, W.; Yokogawa, T.; Kawakami, K.; Smith, S.J.; Gothilf, Y.; et al. Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 21942–21947. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, N.; Knaut, H.; Yoshihara, Y. Cxcl12/Cxcr4 chemokine signaling is required for placode assembly and sensory axon pathfinding in the zebrafish olfactory system. Development 2007, 134, 2459–2468. [Google Scholar] [CrossRef]
- Marsay, K.S.; Greaves, S.; Mahabaleshwar, H.; Ho, C.M.; Roehl, H.; Monk, P.N.; Carney, T.J.; Partridge, L.J. Tetraspanin Cd9b and Cxcl12a/Cxcr4b have a synergistic effect on the control of collective cell migration. PLoS ONE 2021, 16, e0260372. [Google Scholar] [CrossRef]
- Otten, A.B.C.; Kamps, R.; Lindsey, P.; Gerards, M.; Pendeville-Samain, H.; Muller, M.; van Tienen, F.H.J.; Smeets, H.J.M. Tfam Knockdown Results in Reduction of mtDNA Copy Number, OXPHOS Deficiency and Abnormalities in Zebrafish Embryos. Front. Cell Dev. Biol. 2020, 8, 381. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Isolation of Total RNA from Zebrafish Embryos and Adults. Cold Spring Harb. Protoc. 2020, 2020, 101667. [Google Scholar] [CrossRef] [PubMed]
- Dudink, I.; White, T.A.; Ardalan, M.; Mallard, C.; Ballerin, G.; Creed, S.J.; Pham, Y.; Sutherland, A.E.; Castillo-Melendez, M.; Allison, B.J.; et al. An Optimized and Detailed Step-by-Step Protocol for the Analysis of Neuronal Morphology in Golgi-Stained Fetal Sheep Brain. Dev. Neurosci. 2022, 44, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, I.; Levitas-Djerbi, T.; Appelbaum, L. The Hypocretin/Orexin Neuronal Networks in Zebrafish. Behav. Neurosci. Orexin/Hypocretin 2016, 33, 75–92. [Google Scholar] [CrossRef]
- Kaslin, J.; Nystedt, J.M.; Östergård, M.; Peitsaro, N.; Panula, P. The Orexin/Hypocretin System in Zebrafish Is Connected to the Aminergic and Cholinergic Systems. J. Neurosci. 2004, 24, 2678–2689. [Google Scholar] [CrossRef]
- Marquart, G.D.; Tabor, K.M.; Brown, M.; Strykowski, J.L.; Varshney, G.K.; LaFave, M.C.; Mueller, T.; Burgess, S.M.; Higashijima, S.-I.; Burgess, H.A. A 3D Searchable Database of Transgenic Zebrafish Gal4 and Cre Lines for Functional Neuroanatomy Studies. Front. Neural Circuits 2015, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Palasin, K.; Uechi, T.; Yoshihama, M.; Srisowanna, N.; Choijookhuu, N.; Hishikawa, Y.; Kenmochi, N.; Chotigeat, W. Abnormal development of zebrafish after knockout and knockdown of ribosomal protein L10a. Sci. Rep. 2019, 9, 18130. [Google Scholar] [CrossRef]
- Guo, R.; Li, F.; Lu, M.; Ge, K.; Gan, L.; Sheng, D. LIM Homeobox 9 knockdown by morpholino does not affect zebrafish retinal development. Biol. Open 2021, 10, bio056382. [Google Scholar] [CrossRef]
- Poon, K.; Barson, J.R.; Ho, H.T.; Leibowitz, S.F. Relationship of the Chemokine, CXCL12, to Effects of Dietary Fat on Feeding-Related Behaviors and Hypothalamic Neuropeptide Systems. Front. Behav. Neurosci. 2016, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.D.; Yasmin, N.; Khalizova, N.; Campbell, S.; Onoichenco, A.; Fam, M.; Albeg, A.S.; Leibowitz, S.F. Sexually dimorphic and asymmetric effects of embryonic ethanol exposure on hypocretin/orexin neurons as related to behavioral changes in zebrafish. Sci. Rep. 2021, 11, 16078. [Google Scholar] [CrossRef]
- Wu, Y.; Peng, H.; Cui, M.; Whitney, N.P.; Huang, Y.; Zheng, J.C. CXCL12 increases human neural progenitor cell proliferation through Akt-1/FOXO3a signaling pathway. J. Neurochem. 2009, 109, 1157–1167. [Google Scholar] [CrossRef]
- Teicher, B.A.; Fricker, S.P. CXCL12 (SDF-1)/CXCR4 pathway in cancer. Clin. Cancer Res. 2010, 16, 2927–2931. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro-da-Silva, J.; Luchiari, A.C. Embryonic ethanol exposure on zebrafish early development. Brain Behav. 2021, 11, e02062. [Google Scholar] [CrossRef] [PubMed]
- Memi, F.; Abe, P.; Cariboni, A.; MacKay, F.; Parnavelas, J.G.; Stumm, R. CXC Chemokine Receptor 7 (CXCR7) Affects the Migration of GnRH Neurons by Regulating CXCL12 Availability. J. Neurosci. 2013, 33, 17527–17537. [Google Scholar] [CrossRef] [PubMed]
- Lewellis, S.W.; Nagelberg, D.; Subedi, A.; Staton, A.; LeBlanc, M.; Giraldez, A.; Knaut, H. Precise SDF1-mediated cell guidance is achieved through ligand clearance and microRNA-mediated decay. J. Cell Biol. 2013, 200, 337–355. [Google Scholar] [CrossRef]
- Sapède, D.; Rossel, M.; Dambly-Chaudière, C.; Ghysen, A. Role of SDF1 chemokine in the development of lateral line efferent and facial motor neurons. Proc. Natl. Acad. Sci. USA 2005, 102, 1714–1718. [Google Scholar] [CrossRef]
- Poon, K.; Ho, H.T.; Barson, J.R.; Leibowitz, S.F. Stimulatory role of the chemokine CCL2 in the migration and peptide expression of embryonic hypothalamic neurons. J. Neurochem. 2014, 131, 509–520. [Google Scholar] [CrossRef] [PubMed]
- James, M.H.; Aston-Jones, G. Orexin Reserve: A Mechanistic Framework for the Role of Orexins (Hypocretins) in Addiction. Biol. Psychiatry 2022, 92, 836–844. [Google Scholar] [CrossRef]
- Lau, S.; Feitzinger, A.; Venkiteswaran, G.; Wang, J.; Lewellis, S.W.; Koplinski, C.A.; Peterson, F.C.; Volkman, B.F.; Meier-Schellersheim, M.; Knaut, H. A negative-feedback loop maintains optimal chemokine concentrations for directional cell migration. Nat. Cell Biol. 2020, 22, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Hilla, A.M.; Baehr, A.; Leibinger, M.; Andreadaki, A.; Fischer, D. CXCR4/CXCL12-mediated entrapment of axons at the injury site compromises optic nerve regeneration. Proc. Natl. Acad. Sci. USA 2021, 118, e2016409118. [Google Scholar] [CrossRef]
- Knaut, H.; Blader, P.; Strähle, U.; Schier, A.F. Assembly of Trigeminal Sensory Ganglia by Chemokine Signaling. Neuron 2005, 47, 653–666. [Google Scholar] [CrossRef]
- Belmadani, A.; Tran, P.B.; Ren, D.; Assimacopoulos, S.; Grove, E.A.; Miller, R.J. The Chemokine Stromal Cell-Derived Factor-1 Regulates the Migration of Sensory Neuron Progenitors. J. Neurosci. 2005, 25, 3995–4003. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, B.W.; Hall, Z.J.; Heuzé, A.; Joly, J.-S.; Tropepe, V.; Kaslin, J. The role of neuro-epithelial-like and radial-glial stem and progenitor cells in development, plasticity, and repair. Prog. Neurobiol. 2018, 170, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Turrero García, M.; Harwell, C.C. Radial glia in the ventral telencephalon. FEBS Lett. 2017, 591, 3942–3959. [Google Scholar] [CrossRef]
- Bertipaglia, C.; Gonçalves, J.C.; Vallee, R.B. Nuclear migration in mammalian brain development. Semin. Cell Dev. Biol. 2018, 82, 57–66. [Google Scholar] [CrossRef]
- Diotel, N.; Vaillant, C.; Gueguen, M.-M.; Mironov, S.; Anglade, I.; Servili, A.; Pellegrini, E.; Kah, O. Cxcr4 and Cxcl12 expression in radial glial cells of the brain of adult zebrafish. J. Comp. Neurol. 2010, 518, 4855–4876. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.-Q.; Karatayev, O.; Kavya, B.D.S.S.; Leibowitz, S.F. CCL2/CCR2 Chemokine System in Embryonic Hypothalamus: Involvement in Sexually Dimorphic Stimulatory Effects of Prenatal Ethanol Exposure on Peptide-Expressing Neurons. Neuroscience 2020, 424, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Louth, E.L.; Luctkar, H.D.; Heney, K.A.; Bailey, C.D.C. Developmental ethanol exposure alters the morphology of mouse prefrontal neurons in a layer-specific manner. Brain Res. 2018, 1678, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Lieberam, I.; Agalliu, D.; Nagasawa, T.; Ericson, J.; Jessell, T.M. A Cxcl12-Cxcr4 Chemokine Signaling Pathway Defines the Initial Trajectory of Mammalian Motor Axons. Neuron 2005, 47, 667–679. [Google Scholar] [CrossRef]
- Mithal, D.S.; Banisadr, G.; Miller, R.J. CXCL12 Signaling in the Development of the Nervous System. J. Neuroimmune Pharmacol. 2012, 7, 820–834. [Google Scholar] [CrossRef] [PubMed]
- Meyrath, M.; Szpakowska, M.; Zeiner, J.; Massotte, L.; Merz, M.P.; Benkel, T.; Simon, K.; Ohnmacht, J.; Turner, J.D.; Krüger, R.; et al. The atypical chemokine receptor ACKR3/CXCR7 is a broad-spectrum scavenger for opioid peptides. Nat. Commun. 2020, 11, 3033. [Google Scholar] [CrossRef]
- Hayton, S.J.; Mahoney, M.K.; Olmstead, M.C. Behavioral Traits Predicting Alcohol Drinking in Outbred Rats: An Investigation of Anxiety, Novelty Seeking, and Cognitive Flexibility. Alcohol. Clin. Exp. Res. 2012, 36, 594–603. [Google Scholar] [CrossRef]
- Karatayev, O.; Barson, J.R.; Carr, A.J.; Baylan, J.; Chen, Y.W.; Leibowitz, S.F. Predictors of ethanol consumption in adult Sprague-Dawley rats: Relation to hypothalamic peptides that stimulate ethanol intake. Alcohol 2010, 44, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Sterling, M.E.; Chang, G.-Q.; Karatayev, O.; Chang, S.Y.; Leibowitz, S.F. Effects of embryonic ethanol exposure at low doses on neuronal development, voluntary ethanol consumption and related behaviors in larval and adult zebrafish: Role of hypothalamic orexigenic peptides. Behav. Brain Res. 2016, 304, 125–138. [Google Scholar] [CrossRef]
- Pinheiro-Da-Silva, J.; Agues-Barbosa, T.; Luchiari, A.C. Embryonic Exposure to Ethanol Increases Anxiety-Like Behavior in Fry Zebrafish. Alcohol 2020, 55, 581–590. [Google Scholar] [CrossRef]
- Chang, G.Q.; Karatayev, O.; Halkina, V.; Edelstien, J.; Ramirez, E.; Leibowitz, S.F. Hypothalamic CCL2/CCR2 Chemokine System: Role in Sexually Dimorphic Effects of Maternal Ethanol Exposure on Melanin-Concentrating Hormone and Behavior in Adolescent Offspring. J. Neurosci. 2018, 38, 9072–9090. [Google Scholar] [CrossRef] [PubMed]
- James, M.H.; Mahler, S.V.; Moorman, D.E.; Aston-Jones, G. A decade of orexin/hypocretin and addiction: Where are we now? In Behavioral Neuroscience of Orexin/Hypocretin; Lawrence, A.J., De Lucea, L., Eds.; Springer: New York, NY, USA, 2016; Volume 33, pp. 247–281. [Google Scholar]
- Sagi, D.; de Lecea, L.; Appelbaum, L. Heterogeneity of Hypocretin/Orexin Neurons. Front Neurol. Neurosci. 2021, 45, 61–74. [Google Scholar] [CrossRef]
- Muschamp, J.W.; Hollander, J.A.; Thompson, J.L.; Voren, G.; Hassinger, L.C.; Onvani, S.; Kamenecka, T.M.; Borgland, S.L.; Kenny, P.J.; Carlezon, W.A., Jr. Hypocretin (orexin) facilitates reward by attenuating the antireward effects of its cotransmitter dynorphin in ventral tegmental area. Proc. Natl. Acad. Sci. USA 2014, 111, E1648–E1655. [Google Scholar] [CrossRef]
- Wee, C.L.; Song, E.; Nikitchenko, M.; Herrera, K.J.; Wong, S.; Engert, F.; Kunes, S. Social isolation modulates appetite and avoidance behavior via a common oxytocinergic circuit in larval zebrafish. Nat. Commun. 2022, 13, 2573. [Google Scholar] [CrossRef]
- Paredes-Zúñiga, S.; Ormeño, F.; Allende, M.L. Triadimefon triggers circling behavior and conditioned place preference/aversion in zebrafish in a dose dependent manner. Neurotoxicol. Teratol. 2021, 86, 106979. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.E.; Watt, M.J.; Forster, G.L.; Øverli; Bockholt, C.; Renner, K.J.; Summers, C.H. Corticotropin releasing factor induces anxiogenic locomotion in trout and alters serotonergic and dopaminergic activity. Horm. Behav. 2007, 52, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Von Trotha, J.W.; Vernier, P.; Bally-Cuif, L. Emotions and motivated behavior converge on an amygdala-like structure in the zebrafish. Eur. J. Neurosci. 2014, 40, 3302–3315. [Google Scholar] [CrossRef]
- Mathuru, A.S.; Kibat, C.; Cheong, W.F.; Shui, G.; Wenk, M.R.; Friedrich, R.W.; Jesuthasan, S. Chondroitin Fragments Are Odorants that Trigger Fear Behavior in Fish. Curr. Biol. 2012, 22, 538–544. [Google Scholar] [CrossRef]
- Singh, C.; Rihel, J.; Prober, D.A. Neuropeptide Y Regulates Sleep by Modulating Noradrenergic Signaling. Curr. Biol. 2017, 27, 3796–3811.e3795. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Altermatt, M.; Zhang, R.-W.; Coughlin, G.M.; Montz, C.; Gradinaru, V.; Prober, D.A. The Serotonergic Raphe Promote Sleep in Zebrafish and Mice. Neuron 2019, 103, 686–701.e688. [Google Scholar] [CrossRef]
- Singh, C.; Oikonomou, G.; Prober, D.A. Norepinephrine is required to promote wakefulness and for hypocretin-induced arousal in zebrafish. Elife 2015, 4, e07000. [Google Scholar] [CrossRef]
- López-Muciño, L.A.; García-García, F.; Cueto-Escobedo, J.; Acosta-Hernández, M.; Venebra-Muñoz, A.; Rodríguez-Alba, J.C. Sleep loss and addiction. Neurosci. Biobehav. Rev. 2022, 141, 104832. [Google Scholar] [CrossRef]
- Gyawali, U.; James, M.H. Sleep disturbance in substance use disorders: The orexin (hypocretin) system as an emerging pharmacological target. Neuropsychopharmacology 2023, 48, 228–229. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hcrt Projection Branch and Terminal Points in the LFB | ||||
|---|---|---|---|---|
| One-way ANOVA | ||||
| Brain Areas | Measures | df | F | pValue |
| aAH | Branch Points | 2, 10 | 3.88 | 0.05 |
| Terminal Points | 2, 10 | 2.91 | 1.00 | |
| pAH | Branch Points | 2, 10 | 0.47 | 0.63 |
| Terminal Points | 2, 10 | 0.29 | 0.75 | |
| Post-Hoc Tests | ||||
| Brain Areas | Measures | Control + Vehicle | Ethanol + Vehicle | pValue |
| aAH | Branch Points | 8.23 ± 1.79 | 11.08 ± 0.74 | 0.35 |
| Terminal Points | 11.70 ± 1.67 | 16.02 ± 1.49 | 0.19 | |
| pAH | Branch Points | 15.16 ± 2.00 | 11.88 ± 2.04 | 0.59 |
| Terminal Points | 21.93 ± 3.78 | 18.26 ± 2.51 | 0.71 | |
| Brain Areas | Measures | Control + Vehicle | Control + cxcl12amRNA | pValue |
| aAH | Branch Points | 8.23 ± 1.79 | 5.85 ± 1.17 | 0.38 |
| Terminal Points | 11.70 ± 1.67 | 10.87 ± 0.63 | 0.67 | |
| pAH | Branch Points | 15.16 ± 2.00 | 13.93 ± 3.26 | 0.93 |
| Terminal Points | 21.93 ± 3.78 | 19.32 ± 4.35 | 0.71 | |
| Hcrt Projection Branch and Terminal Points in the LFB | ||||
|---|---|---|---|---|
| One-way ANOVA | ||||
| Brain Areas | Measures | df | F | pValue |
| aAH | Branch Points | 2, 10 | 3.88 | 0.05 |
| Terminal Points | 2, 10 | 2.91 | 1.00 | |
| pAH | Branch Points | 2, 10 | 0.47 | 0.63 |
| Terminal Points | 2, 10 | 0.29 | 0.75 | |
| Post-Hoc Tests | ||||
| Brain Areas | Measures | Control + Control MO | Ethanol + Control MO | pValue |
| aAH | Branch Points | 10.44 ± 2.99 | 8.11 ± 1.44 | 0.97 |
| Terminal Points | 14.59 ± 3.12 | 11.81 ± 1.83 | 0.52 | |
| pAH | Branch Points | 12.21 ± 1.51 | 10.60 ± 1.83 | 0.69 |
| Terminal Points | 20.84 ± 1.59 | 19.03 ± 3.02 | 0.64 | |
| Brain Areas | Measures | Ethanol + Control MO | Ethanol + cxcl12aMOsp | pValue |
| aAH | Branch Points | 8.11 ± 1.44 | 2.99 ± 0.77 | 0.15 |
| Terminal Points | 11.81 ± 1.83 | 5.15 ± 0.1.51 | 0.20 | |
| pAH | Branch Points | 10.60 ± 1.83 | 7.68 ± 1.97 | 0.57 |
| Terminal Points | 19.03 ± 3.02 | 11.96 ± 2.59 | 0.16 | |
| Hcrt Neuron Number | Two-Way ANOVA Post-Hoc Tests | ||||
|---|---|---|---|---|---|
| Brain Areas | Measures | Control + Control MO | Control + cxcl12aMOsp | pValue | |
| aAH | # Hcrt neurons | 9.50 ± 0.28 | 10.00 ± 0.41 | 0.85 | |
| POA | # Hcrt neurons | 0 | 0 | NA | |
| pAH | # Hcrt neurons | 9.25 ± 0.47 | 9.25 ± 0.75 | 0.99 | |
| Hcrt Projection Branch and Terminal Points | ttest results | ||||
| Brain Areas | Projection Areas | Measures | Control + Control MO | Control + cxcl12aMOsp | pValue |
| aAH | SP | Branch Points | 3.30 ± 0.45 | 0.54 ± 0.34 | 0.003 ** |
| Terminal Points | 7.14 ± 1.03 | 3.00 ± 1.30 | 0.047 * | ||
| DP | Branch Points | 0 | 0 | NA | |
| Terminal Points | 0 | 0 | NA | ||
| LFB | Branch Points | 10.44 ± 2.99 | 6.05 ± 1.66 | 0.24 | |
| Terminal Points | 14.59 ± 3.12 | 16.01 ± 1.52 | 0.19 | ||
| POA | SP | Branch Points | 0 | 0 | NA |
| Terminal Points | 0 | 0 | NA | ||
| DP | Branch Points | 0 | 0 | NA | |
| Terminal Points | 0 | 0 | NA | ||
| pAH | LC | Branch Points | 60.52 ± 1.51 | 92.43 ± 23.18 | 0.32 |
| Terminal Points | 80.98 ± 8.95 | 111.24 ± 24.73 | 0.16 | ||
| LFB | Branch Points | 12.21 ± 1.51 | 8.50 ± 1.50 | 0.13 | |
| Terminal Points | 20.84 ± 1.59 | 14.77 ± 2.51 | 0.08 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasmin, N.; Collier, A.D.; Abdulai, A.R.; Karatayev, O.; Yu, B.; Fam, M.; Leibowitz, S.F. Role of Chemokine Cxcl12a in Mediating the Stimulatory Effects of Ethanol on Embryonic Development of Subpopulations of Hypocretin/Orexin Neurons and Their Projections. Cells 2023, 12, 1399. https://doi.org/10.3390/cells12101399
Yasmin N, Collier AD, Abdulai AR, Karatayev O, Yu B, Fam M, Leibowitz SF. Role of Chemokine Cxcl12a in Mediating the Stimulatory Effects of Ethanol on Embryonic Development of Subpopulations of Hypocretin/Orexin Neurons and Their Projections. Cells. 2023; 12(10):1399. https://doi.org/10.3390/cells12101399
Chicago/Turabian StyleYasmin, Nushrat, Adam D. Collier, Abdul R. Abdulai, Olga Karatayev, Boyi Yu, Milisia Fam, and Sarah F. Leibowitz. 2023. "Role of Chemokine Cxcl12a in Mediating the Stimulatory Effects of Ethanol on Embryonic Development of Subpopulations of Hypocretin/Orexin Neurons and Their Projections" Cells 12, no. 10: 1399. https://doi.org/10.3390/cells12101399