
Acute and Chronic Ethanol Effects during Adolescence on Neuroimmune Responses: Consequences and Potential Pharmacologic Interventions

Abstract
:1. Introduction
2. Acute and Chronic Effects of Adolescent Binge Drinking on Toll-like Receptors
3. Acute and Chronic Effects of Adolescent Binge Drinking on Cytokines and Chemokines
4. Acute and Chronic Effects of Adolescent Binge Drinking on Microglia
5. Acute and Chronic Effects of Adolescent Binge Drinking on Astrocytes
6. Alcohol Use Interventions and Drug Treatments
6.1. Anticonvulsant Drugs
6.2. Acetylcholinesterase Inhibitors
6.3. Opioid Antagonist
6.4. Lipid-Lowering Agents
6.5. Classic Anti-Inflammatory Drugs
6.6. Exercise
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinberg, L. A Social Neuroscience Perspective on Adolescent Risk-Taking. Dev. Rev. 2008, 28, 78–106. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H. Smoking and adolescent health. Korean J. Pediatr. 2011, 54, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Ritchwood, T.D.; Ford, H.; DeCoster, J.; Sutton, M.; Lochman, J.E. Risky Sexual Behavior and Substance Use among Adolescents: A Meta-analysis. Child. Youth Serv. Rev. 2015, 52, 74–88. [Google Scholar] [CrossRef]
- Chung, T.; Creswell, K.G.; Bachrach, R.; Clark, D.B.; Martin, C.S. Adolescent Binge Drinking. Alcohol Res. 2018, 39, 5–15. [Google Scholar] [PubMed]
- Substance Abuse and Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2019 National Survey on Drug Use and Health 2020; Center for Behavioral Health Statistics and Quality, Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2020.
- Sudhinaraset, M.; Wigglesworth, C.; Takeuchi, D.T. Social and Cultural Contexts of Alcohol Use: Influences in a Social-Ecological Framework. Alcohol Res. 2016, 38, 35–45. [Google Scholar]
- Crews, F.T.; Zou, J.; Qin, L. Induction of innate immune genes in brain create the neurobiology of addiction. Brain Behav. Immun. 2011, 25 (Suppl. S1), S4–S12. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, A.T.; Nadim, H.; Devine, L.; Jatlow, P.; O’Malley, S.S. Acute alcohol consumption alters the peripheral cytokines IL-8 and TNF-α. Alcohol 2020, 85, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Hanke, M.L.; Kielian, T. Toll-like receptors in health and disease in the brain: Mechanisms and therapeutic potential. Clin. Sci. 2011, 121, 367–387. [Google Scholar] [CrossRef]
- Alfonso-Loeches, S.; Pascual-Lucas, M.; Blanco, A.M.; Sanchez-Vera, I.; Guerri, C. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. J. Neurosci. 2010, 30, 8285–8295. [Google Scholar] [CrossRef]
- Blanco, A.M.; Perez-Arago, A.; Fernandez-Lizarbe, S.; Guerri, C. Ethanol mimics ligand-mediated activation and endocytosis of IL-1RI/TLR4 receptors via lipid rafts caveolae in astroglial cells. J. Neurochem. 2008, 106, 625–639. [Google Scholar] [CrossRef]
- Fernandez-Lizarbe, S.; Pascual, M.; Gascon, M.S.; Blanco, A.; Guerri, C. Lipid rafts regulate ethanol-induced activation of TLR4 signaling in murine macrophages. Mol. Immunol. 2008, 45, 2007–2016. [Google Scholar] [CrossRef]
- Wu, R.; Li, J.X. Toll-Like Receptor 4 Signaling and Drug Addiction. Front. Pharmacol. 2020, 11, 603445. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.H.W.; Buisman-Pijlman, F.T.; Mustafa, S.; Rice, K.C.; Hutchinson, M.R. Antagonising TLR4-TRIF signalling before or after a low-dose alcohol binge during adolescence prevents alcohol drinking but not seeking behaviour in adulthood. Neuropharmacology 2018, 128, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Deschamps, C.; Uyttersprot, F.; Debris, M.; Marie, C.; Fouquet, G.; Marcq, I.; Vilpoux, C.; Naassila, M.; Pierrefiche, O. Anti-inflammatory drugs prevent memory and hippocampal plasticity deficits following initial binge-like alcohol exposure in adolescent male rats. Psychopharmacology 2022, 239, 2245–2262. [Google Scholar] [CrossRef] [PubMed]
- Vetreno, R.P.; Lawrimore, C.J.; Rowsey, P.J.; Crews, F.T. Persistent Adult Neuroimmune Activation and Loss of Hippocampal Neurogenesis Following Adolescent Ethanol Exposure: Blockade by Exercise and the Anti-inflammatory Drug Indomethacin. Front. Neurosci. 2018, 12, 200. [Google Scholar] [CrossRef]
- Vetreno, R.P.; Crews, F.T. Adolescent binge drinking increases expression of the danger signal receptor agonist HMGB1 and Toll-like receptors in the adult prefrontal cortex. Neuroscience 2012, 226, 475–488. [Google Scholar] [CrossRef]
- Crews, F.T.; Walter, T.J.; Coleman, L.G., Jr.; Vetreno, R.P. Toll-like receptor signaling and stages of addiction. Psychopharmacology 2017, 234, 1483–1498. [Google Scholar] [CrossRef]
- Silva-Gotay, A.; Davis, J.; Tavares, E.R.; Richardson, H.N. Alcohol drinking during early adolescence activates microglial cells and increases frontolimbic Interleukin-1 beta and Toll-like receptor 4 gene expression, with heightened sensitivity in male rats compared to females. Neuropharmacology 2021, 197, 108698. [Google Scholar] [CrossRef]
- Li, Q.; Liu, D.; Pan, F.; Ho, C.S.H.; Ho, R.C.M. Ethanol Exposure Induces Microglia Activation and Neuroinflammation through TLR4 Activation and SENP6 Modulation in the Adolescent Rat Hippocampus. Neural Plast. 2019, 2019, 1648736. [Google Scholar] [CrossRef]
- Orio, L.; Anton, M.; Rodriguez-Rojo, I.C.; Correas, A.; Garcia-Bueno, B.; Corral, M.; de Fonseca, F.R.; Garcia-Moreno, L.M.; Maestu, F.; Cadaveira, F. Young alcohol binge drinkers have elevated blood endotoxin, peripheral inflammation and low cortisol levels: Neuropsychological correlations in women. Addict. Biol. 2018, 23, 1130–1144. [Google Scholar] [CrossRef]
- Peng, H.; Nixon, K. Microglia Phenotypes Following the Induction of Alcohol Dependence in Adolescent Rats. Alcohol. Clin. Exp. Res. 2021, 45, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Galán-Llario, M.; Rodríguez-Zapata, M.; Gramage, E.; Vicente-Rodríguez, M.; Fontán-Baselga, T.; Ovejero-Benito, M.C.; Pérez-García, C.; Carrasco, J.; Moreno-Herradón, M.; Sevillano, J.; et al. Receptor protein tyrosine phosphatase β/ζ regulates loss of neurogenesis in the mouse hippocampus following adolescent acute ethanol exposure. NeuroToxicology 2023, 94, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Barney, T.M.; Vore, A.S.; Deak, T. Acute Ethanol Challenge Differentially Regulates Expression of Growth Factors and miRNA Expression Profile of Whole Tissue of the Dorsal Hippocampus. Front. Neurosci. 2022, 16, 884197. [Google Scholar] [CrossRef] [PubMed]
- Fuster, J.J.; Walsh, K. The good, the bad, and the ugly of interleukin-6 signaling. EMBO J. 2014, 33, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Bellozi, P.M.Q.; Pelicao, R.; Santos, M.C.; Lima, I.V.A.; Saliba, S.W.; Vieira, E.L.M.; Campos, A.C.; Teixeira, A.L.; de Oliveira, A.C.P.; Nakamura-Palacios, E.M.; et al. URB597 ameliorates the deleterious effects induced by binge alcohol consumption in adolescent rats. Neurosci. Lett. 2019, 711, 134408. [Google Scholar] [CrossRef]
- Gomez, G.I.; Falcon, R.V.; Maturana, C.J.; Labra, V.C.; Salgado, N.; Rojas, C.A.; Oyarzun, J.E.; Cerpa, W.; Quintanilla, R.A.; Orellana, J.A. Heavy Alcohol Exposure Activates Astroglial Hemichannels and Pannexons in the Hippocampus of Adolescent Rats: Effects on Neuroinflammation and Astrocyte Arborization. Front. Cell. Neurosci. 2018, 12, 472. [Google Scholar] [CrossRef]
- Gonzalez-Portilla, M.; Montagud-Romero, S.; Navarrete, F.; Gasparyan, A.; Manzanares, J.; Minarro, J.; Rodriguez-Arias, M. Pairing Binge Drinking and a High-Fat Diet in Adolescence Modulates the Inflammatory Effects of Subsequent Alcohol Consumption in Mice. Int. J. Mol. Sci. 2021, 22, 5279. [Google Scholar] [CrossRef]
- Pfefferbaum, A.; Rosenbloom, M.; Deshmukh, A.; Sullivan, E. Sex differences in the effects of alcohol on brain structure. Am. J. Psychiatry 2001, 158, 188–197. [Google Scholar] [CrossRef]
- Nwachukwu, K.N.; Healey, K.L.; Swartzwelder, H.S.; Marshall, S.A. The Influence of Sex on Hippocampal Neurogenesis and Neurotrophic Responses on the Persistent Effects of Adolescent Intermittent Ethanol Exposure into Adulthood. Neuroscience 2022, 506, 68–79. [Google Scholar] [CrossRef]
- Vore, A.S.; Barney, T.M.; Gano, A.; Varlinskaya, E.I.; Deak, T. Adolescent intermittent ethanol (AIE) produces sex specific alterations in adult neuroimmune gene expression and ethanol sensitivity that are independent of ethanol metabolism. Neuropharmacology 2021, 195, 108635. [Google Scholar] [CrossRef]
- Saijo, K.; Glass, C.K. Microglial cell origin and phenotypes in health and disease. Nat. Rev. Immunol. 2011, 11, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.J.; Xue, Q.S. Life and death of microglia. J. Neuroimmune Pharmacol. 2009, 4, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Grifasi, I.R.; Evans, W.A.; Rexha, A.D.; Sako, L.W.; Marshall, S.A. A comparison of hippocampal microglial responses in aged and young rodents following dependent and non-dependent binge drinking. Int. Rev. Neurobiol. 2019, 148, 305–343. [Google Scholar] [PubMed]
- Grifasi, I.R.; McIntosh, S.E.; Thomas, R.D.; Lysle, D.T.; Thiele, T.E.; Marshall, S.A. Characterization of the Hippocampal Neuroimmune Response to Binge-Like Ethanol Consumption in the Drinking in the Dark Model. Neuroimmunomodulation 2019, 26, 19–32. [Google Scholar] [CrossRef]
- Mouihate, A.; Galic, M.A.; Ellis, S.L.; Spencer, S.J.; Tsutsui, S.; Pittman, Q.J. Early life activation of toll-like receptor 4 reprograms neural anti-inflammatory pathways. J. Neurosci. 2010, 30, 7975–7983. [Google Scholar] [CrossRef]
- Lacagnina, M.J.; Rivera, P.D.; Bilbo, S.D. Glial and Neuroimmune Mechanisms as Critical Modulators of Drug Use and Abuse. Neuropsychopharmacology 2017, 42, 156–177. [Google Scholar] [CrossRef]
- Bilbo, S.D.; Schwarz, J.M. Early-life programming of later-life brain and behavior: A critical role for the immune system. Front. Behav. Neurosci. 2009, 3, 14. [Google Scholar] [CrossRef]
- Marshall, S.A.; McClain, J.A.; Wooden, J.I.; Nixon, K. Microglia Dystrophy Following Binge-Like Alcohol Exposure in Adolescent and Adult Male Rats. Front. Neuroanat. 2020, 14, 52. [Google Scholar] [CrossRef]
- Ji, Z.; Yuan, L.; Lu, X.; Ding, H.; Luo, J.; Ke, Z.J. Binge Alcohol Exposure Causes Neurobehavioral Deficits and GSK3beta Activation in the Hippocampus of Adolescent Rats. Sci. Rep. 2018, 8, 3088. [Google Scholar] [CrossRef]
- Vilpoux, C.; Fouquet, G.; Deschamps, C.; Lefebvre, E.; Gosset, P.; Antol, J.; Zabijak, L.; Marcq, I.; Naassila, M.; Pierrefiche, O. Astrogliosis and compensatory neurogenesis after the first ethanol binge drinking-like exposure in the adolescent rat. Alcohol. Clin. Exp. Res. 2022, 46, 207–220. [Google Scholar] [CrossRef]
- Guo, Y.; Yan, M.; Li, L.; Zhao, L.; Li, Y. Treadmill Exercise Prevents Cognitive Impairments in Adolescent Intermittent Ethanol Rats by Reducing the Excessive Activation of Microglia Cell in the Hippocampus. Int. J. Mol. Sci. 2022, 23, 14701. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, K.N.; King, D.M.; Healey, K.L.; Swartzwelder, H.S.; Marshall, S.A. Sex-specific effects of adolescent intermittent ethanol exposure-induced dysregulation of hippocampal glial cells in adulthood. Alcohol 2022, 100, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, D.; Zhang, Y.; Cai, Z.; Ye, T.; Tong, L.; Xu, X.; Lu, J.; Liu, F.; Lu, X.; et al. Apoptosis-triggered decline in hippocampal microglia mediates adolescent intermittent alcohol exposure-induced depression-like behaviors in mice. Neuropharmacology 2020, 170, 108054. [Google Scholar] [CrossRef]
- Guo, J.; Qiu, T.; Wang, L.; Shi, L.; Ai, M.; Xia, Z.; Peng, Z.; Zheng, A.; Li, X.; Kuang, L. Microglia Loss and Astrocyte Activation Cause Dynamic Changes in Hippocampal [(18)F]DPA-714 Uptake in Mouse Models of Depression. Front. Cell. Neurosci. 2022, 16, 802192. [Google Scholar] [CrossRef]
- Sanchez-Alavez, M.; Nguyen, W.; Mori, S.; Wills, D.N.; Otero, D.; Aguirre, C.A.; Singh, M.; Ehlers, C.L.; Conti, B. Time Course of Blood and Brain Cytokine/Chemokine Levels Following Adolescent Alcohol Exposure and Withdrawal in Rats. Alcohol. Clin. Exp. Res. 2019, 43, 2547–2558. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.; Rizor, A.; Lee, J.; Aschner, M.; Lee, E. The role of astrocytic glutamate transporters GLT-1 and GLAST in neurological disorders: Potential targets for neurotherapeutics. Neuropharmacology 2019, 161, 107559. [Google Scholar] [CrossRef]
- Vainchtein, I.D.; Molofsky, A.V. Astrocytes and Microglia: In Sickness and in Health. Trends Neurosci. 2020, 43, 144–154. [Google Scholar] [CrossRef]
- Crews, F.T.P.D. Alcohol-Related Neurodegeneration and Recovery: Mechanisms from Animal Models. Alcohol Res. Health 2008, 31, 377–388. [Google Scholar]
- Scheetz, A.J.; Markham, J.A.; Fifkova, E. Astrocyte proliferation precedes a decrease in basket cells in the dentate fascia following chronic ethanol treatment in mice. Brain Res. 1988, 460, 246–252. [Google Scholar] [CrossRef]
- Adermark, L.; Bowers, M.S. Disentangling the Role of Astrocytes in Alcohol Use Disorder. Alcohol. Clin. Exp. Res. 2016, 40, 1802–1816. [Google Scholar] [CrossRef]
- Fernandes, L.M.P.; Cartagenes, S.C.; Barros, M.A.; Carvalheiro, T.; Castro, N.C.F.; Schamne, M.G.; Lima, R.R.; Prediger, R.D.; Monteiro, M.C.; Fontes-Junior, E.A.; et al. Repeated cycles of binge-like ethanol exposure induce immediate and delayed neurobehavioral changes and hippocampal dysfunction in adolescent female rats. Behav. Brain Res. 2018, 350, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Loeches, S.; Pascual, M.; Guerri, C. Gender differences in alcohol-induced neurotoxicity and brain damage. Toxicology 2013, 311, 27–34. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, C.J.; Hashimoto, J.G.; Roberts, M.L.; Bloom, S.H.; Andrew, M.R.; Wiren, K.M. Astrocyte Dysfunction Induced by Alcohol in Females but Not Males. Brain Pathol. 2016, 26, 433–451. [Google Scholar] [CrossRef] [PubMed]
- Brewton, H.W.; Robinson, S.L.; Thiele, T.E. Astrocyte expression in the extended amygdala of C57BL/6J mice is sex-dependently affected by chronic intermittent and binge-like ethanol exposure. Alcohol 2023, 108, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Del Riotao, I.; Puente, N.; Penasco, S.; Rico, I.; Gutierrez-Rodriotaguez, A.; Elezgarai, I.; Ramos, A.; Reguero, L.; Gerrikagoitia, I.; Christie, B.R.; et al. Adolescent ethanol intake alters cannabinoid type-1 receptor localization in astrocytes of the adult mouse hippocampus. Addict. Biol. 2019, 24, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.L.; Bell, A.; Scofield, M.D.; Swartzwelder, H.S. Adolescent intermittent ethanol exposure reduces astrocyte-synaptic proximity in the adult medial prefrontal cortex in rats: Reversal by gabapentin. Addict. Neurosci. 2022, 4, 100047. [Google Scholar] [CrossRef]
- Healey, K.L.; Kibble, S.; Hodges, S.; Reissner, K.J.; Testen, A.; Wills, T.A.; Acheson, S.K.; Siemsen, B.M.; McFaddin, J.A.; Scofield, M.D.; et al. Enduring alterations in hippocampal astrocytesynaptic proximity following adolescent alcohol exposure: Reversal by gabapentin. Neural Regen. Res. 2020, 15, 1496–1501. [Google Scholar]
- Walker, C.D.; Sexton, H.G.; Hyde, J.; Greene, B.; Risher, M.L. Diverging Effects of Adolescent Ethanol Exposure on Tripartite Synaptic Development across Prefrontal Cortex Subregions. Cells 2022, 11, 3111. [Google Scholar] [CrossRef]
- Healey, K.L.; Kibble, S.; Bell, A.; Hodges, S.; Swartzwelder, H.S. Effects of adolescent intermittent ethanol on hippocampal expression of glutamate homeostasis and astrocyte-neuronal tethering proteins in male and female rats. J. Neurosci. Res. 2021, 99, 1908–1921. [Google Scholar] [CrossRef]
- Xing, L.; Yang, T.; Cui, S.; Chen, G. Connexin Hemichannels in Astrocytes: Role in CNS Disorders. Front. Mol. Neurosci. 2019, 12, 23. [Google Scholar] [CrossRef]
- Nwachukwu, K.N.; Evans, W.A.; Sides, T.R.; Trevisani, C.P.; Davis, A.; Marshall, S.A. Chemogenetic manipulation of astrocytic signaling in the basolateral amygdala reduces binge-like alcohol consumption in male mice. J. Neurosci. Res. 2021, 99, 1957–1972. [Google Scholar] [CrossRef] [PubMed]
- Blednov, Y.A.; Benavidez, J.M.; Black, M.; Harris, R.A. Inhibition of phosphodiesterase 4 reduces ethanol intake and preference in C57BL/6J mice. Front. Neurosci. 2014, 8, 129. [Google Scholar] [CrossRef]
- Bell, R.L.; Lopez, M.F.; Cui, C.; Egli, M.; Johnson, K.W.; Franklin, K.M.; Becker, H.C. Ibudilast reduces alcohol drinking in multiple animal models of alcohol dependence. Addict. Biol. 2013, 20, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; Casachahua, J.D.; Rinker, J.A.; Blose, A.K.; Lysle, D.T.; Thiele, T.E. IL-1 receptor signaling in the basolateral amygdala modulates binge-like ethanol consumption in male C57BL/6J mice. Brain Behav. Immun. 2016, 51, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Cabrera, M.A.; Chastain, L.G.; Boyadjieva, N.I.; Jabbar, S.; Franklin, T.; Sarkar, D.K. Mu-opioid receptor and delta-opioid receptor differentially regulate microglial inflammatory response to control proopiomelanocortin neuronal apoptosis in the hypothalamus: Effects of neonatal alcohol. J. Neuroinflamm. 2017, 14, 83. [Google Scholar] [CrossRef] [PubMed]
- Erickson, E.K.; Grantham, E.K.; Warden, A.S.; Harris, R.A. Neuroimmune signaling in alcohol use disorder. Pharmacol. Biochem. Behav. 2019, 177, 34–60. [Google Scholar] [CrossRef]
- Yang, J.L.; Xu, B.; Li, S.S.; Zhang, W.S.; Xu, H.; Deng, X.M.; Zhang, Y.Q. Gabapentin reduces CX3CL1 signaling and blocks spinal microglial activation in monoarthritic rats. Mol. Brain 2012, 5, 18. [Google Scholar] [CrossRef]
- Li, X.; Wang, B.; Yu, N.; Yang, L.; Nan, C.; Sun, Z.; Guo, L.; Zhao, Z. Gabapentin Alleviates Brain Injury in Intracerebral Hemorrhage Through Suppressing Neuroinflammation and Apoptosis. Neurochem. Res. 2022, 47, 3063–3075. [Google Scholar] [CrossRef]
- Anton, R.F.; Latham, P.; Voronin, K.; Book, S.; Hoffman, M.; Prisciandaro, J.; Bristol, E. Efficacy of Gabapentin for the Treatment of Alcohol Use Disorder in Patients with Alcohol Withdrawal Symptoms: A Randomized Clinical Trial. JAMA Intern. Med. 2020, 180, 728–736. [Google Scholar] [CrossRef]
- Furieri, F.A.; Nakamura-Palacios, E.M. Gabapentin reduces alcohol consumption and craving: A randomized, double-blind, placebo-controlled trial. J. Clin. Psychiatry 2007, 68, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Swartzwelder, H.S.; Park, M.H.; Acheson, S. Adolescent Ethanol Exposure Enhances NMDA Receptor-Mediated Currents in Hippocampal Neurons: Reversal by Gabapentin. Sci. Rep. 2017, 7, 13133. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, P.J.; Teppen, T.L.; Miller, K.M.; Sexton, H.G.; Pandey, S.C.; Swartzwelder, H.S. Donepezil Reverses Dendritic Spine Morphology Adaptations and Fmr1 Epigenetic Modifications in Hippocampus of Adult Rats After Adolescent Alcohol Exposure. Alcohol. Clin. Exp. Res. 2018, 42, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Macht, V.; Vetreno, R.; Elchert, N.; Crews, F. Galantamine prevents and reverses neuroimmune induction and loss of adult hippocampal neurogenesis following adolescent alcohol exposure. J. Neuroinflamm. 2021, 18, 212. [Google Scholar] [CrossRef]
- Montesinos, J.; Gil, A.; Guerri, C. Nalmefene Prevents Alcohol-Induced Neuroinflammation and Alcohol Drinking Preference in Adolescent Female Mice: Role of TLR4. Alcohol. Clin. Exp. Res. 2017, 41, 1257–1270. [Google Scholar] [CrossRef]
- Tournier, N.; Pottier, G.; Caille, F.; Coulon, C.; Goislard, M.; Jego, B.; Negroni, J.; Leroy, C.; Saba, W. Nalmefene alleviates the neuroimmune response to repeated binge-like ethanol exposure: A TSPO PET imaging study in adolescent rats. Addict. Biol. 2021, 26, e12962. [Google Scholar] [CrossRef]
- Rivera-Meza, M.; Muñoz, D.; Jerez, E.; Quintanilla, M.E.; Salinas-Luypaert, C.; Fernandez, K.; Karahanian, E. Fenofibrate Administration Reduces Alcohol and Saccharin Intake in Rats: Possible Effects at Peripheral and Central Levels. Front. Behav. Neurosci. 2017, 11, 133. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Benavidez, J.M.; Black, M.; Ferguson, L.B.; Schoenhard, G.L.; Goate, A.M.; Edenberg, H.J.; Wetherill, L.; Hesselbrock, V.; Foroud, T.; et al. Peroxisome proliferator-activated receptors α and γ are linked with alcohol consumption in mice and withdrawal and dependence in humans. Alcohol. Clin. Exp. Res. 2015, 39, 136–145. [Google Scholar] [CrossRef]
- Villavicencio-Tejo, F.; Flores-Bastias, O.; Marambio-Ruiz, L.; Perez-Reytor, D.; Karahanian, E. Fenofibrate (a PPAR-alpha Agonist) Administered During Ethanol Withdrawal Reverts Ethanol-Induced Astrogliosis and Restores the Levels of Glutamate Transporter in Ethanol-Administered Adolescent Rats. Front. Pharmacol. 2021, 12, 653175. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Kumar, S.; Sehgal, A.; Singh, S.; Kumari, S.; Brisc, M.C.; Munteanu, M.A.; Brisc, C.; Buhas, C.L.; Judea-Pusta, C.; et al. Rice bran, an off-shoot to newer therapeutics in neurological disorders. Biomed. Pharmacother. 2021, 140, 111796. [Google Scholar] [CrossRef]
- Berger, A.; Rein, D.; Schäfer, A.; Monnard, I.; Gremaud, G.; Lambelet, P.; Bertoli, C. Similar cholesterol–lowering propertiesof rice bran oil, with varied γ–oryzanol, in mildly hypercholesterolemic men*. Eur. J. Nutr. 2005, 44, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Jolfaie, N.R.; Rouhani, M.H.; Surkan, P.J.; Siassi, F.; Azadbakht, L. Rice Bran Oil Decreases Total and LDL Cholesterol in Humans: A Systematic Review and Meta-Analysis of Randomized Controlled Clinical Trials. Horm. Metab. Res. 2016, 48, 417–426. [Google Scholar] [CrossRef]
- Akter, S.; Uddin, K.R.; Sasaki, H.; Lyu, Y.; Shibata, S. Gamma oryzanol impairs alcohol-induced anxiety-like behavior in mice via upregulation of central monoamines associated with Bdnf and Il-1beta signaling. Sci. Rep. 2020, 10, 10677. [Google Scholar] [CrossRef] [PubMed]
- Vetreno, R.P.; Crews, F.T. Adolescent binge ethanol-induced loss of basal forebrain cholinergic neurons and neuroimmune activation are prevented by exercise and indomethacin. PLoS ONE 2018, 13, e0204500. [Google Scholar] [CrossRef] [PubMed]
- Macht, V.; Vetreno, R.; Elchert, N.; Fisher, R.; Crews, F. Indomethacin restores loss of hippocampal neurogenesis and cholinergic innervation and reduces innate immune expression and reversal learning deficits in adult male and female rats following adolescent ethanol exposure. Alcohol. Clin. Exp. Res. 2023, 47, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Grodin, E.N.; Bujarski, S.; Towns, B.; Burnette, E.; Nieto, S.; Lim, A.; Lin, J.; Miotto, K.; Gillis, A.; Irwin, M.R.; et al. Ibudilast, a neuroimmune modulator, reduces heavy drinking and alcohol cue-elicited neural activation: A randomized trial. Transl. Psychiatry 2021, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Ray, L.A.; Bujarski, S.; Shoptaw, S.; Roche, D.J.; Heinzerling, K.; Miotto, K. Development of the Neuroimmune Modulator Ibudilast for the Treatment of Alcoholism: A Randomized, Placebo-Controlled, Human Laboratory Trial. Neuropsychopharmacology 2017, 42, 1776–1788. [Google Scholar] [CrossRef]
- Avila, D.V.; Myers, S.A.; Zhang, J.; Kharebava, G.; McClain, C.J.; Kim, H.Y.; Whittemore, S.R.; Gobejishvili, L.; Barve, S. Phosphodiesterase 4b expression plays a major role in alcohol-induced neuro-inflammation. Neuropharmacology 2017, 125, 376–385. [Google Scholar] [CrossRef]
- Garrido-Mesa, N.; Zarzuelo, A.; Gálvez, J. Minocycline: Far beyond an antibiotic. Br. J. Pharmacol. 2013, 169, 337–352. [Google Scholar] [CrossRef]
- Doremus-Fitzwater, T.L.; Buck, H.M.; Bordner, K.; Richey, L.; Jones, M.E.; Deak, T. Intoxication- and withdrawal-dependent expression of central and peripheral cytokines following initial ethanol exposure. Alcohol. Clin. Exp. Res. 2014, 38, 2186–2198. [Google Scholar] [CrossRef]
- Agrawal, R.G.; Hewetson, A.; George, C.M.; Syapin, P.J.; Bergeson, S.E. Minocycline reduces ethanol drinking. Brain Behav. Immun. 2011, 25 (Suppl. S1), S165–S169. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lousberg, E.L.; Moldenhauer, L.M.; Hayball, J.D.; Robertson, S.A.; Coller, J.K.; Watkins, L.R.; Somogyi, A.A.; Hutchinson, M.R. Attenuation of microglial and IL-1 signaling protects mice from acute alcohol-induced sedation and/or motor impairment. Brain Behav. Immun. 2011, 25 (Suppl. S1), S155–S164. [Google Scholar] [CrossRef] [PubMed]
- Vetreno, R.P.; Bohnsack, J.P.; Kusumo, H.; Liu, W.; Pandey, S.C.; Crews, F.T. Neuroimmune and epigenetic involvement in adolescent binge ethanol-induced loss of basal forebrain cholinergic neurons: Restoration with voluntary exercise. Addict. Biol. 2020, 25, e12731. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, M.E.; Tampier, L.; Sapag, A.; Gerdtzen, Z.; Israel, Y. Sex differences, alcohol dehydrogenase, acetaldehyde burst, and aversion to ethanol in the rat: A systems perspective. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E531–E537. [Google Scholar] [CrossRef]
- Thomasson, H.R. Gender differences in alcohol metabolism. Physiological responses to ethanol. Recent Dev. Alcohol 1995, 12, 163–179. [Google Scholar]
- Marshall, S.A.; Geil, C.R.; Nixon, K. Prior Binge Ethanol Exposure Potentiates the Microglial Response in a Model of Alcohol-Induced Neurodegeneration. Brain Sci. 2016, 6, 16. [Google Scholar] [CrossRef]
- Zhao, Y.N.; Wang, F.; Fan, Y.X.; Ping, G.F.; Yang, J.Y.; Wu, C.F. Activated microglia are implicated in cognitive deficits, neuronal death, and successful recovery following intermittent ethanol exposure. Behav. Brain Res. 2013, 236, 270–282. [Google Scholar] [CrossRef]
- Downs, A.M.; Catavero, C.M.; Kasten, M.R.; McElligott, Z.A. Tauopathy and alcohol consumption interact to alter locus coeruleus excitatory transmission and excitability in male and female mice. Alcohol 2023, 107, 97–107. [Google Scholar] [CrossRef]
- Tucker, A.E.; Alicea Pauneto, C.D.M.; Barnett, A.M.; Coleman, L.G., Jr. Chronic Ethanol Causes Persistent Increases in Alzheimer’s Tau Pathology in Female 3xTg-AD Mice: A Potential Role for Lysosomal Impairment. Front. Behav. Neurosci. 2022, 16, 886634. [Google Scholar] [CrossRef]
- Hoffman, J.L.; Faccidomo, S.; Kim, M.; Taylor, S.M.; Agoglia, A.E.; May, A.M.; Smith, E.N.; Wong, L.C.; Hodge, C.W. Alcohol drinking exacerbates neural and behavioral pathology in the 3xTg-AD mouse model of Alzheimer’s disease. In International Review of Neurobiology; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Barnett, A.; David, E.; Rohlman, A.; Nikolova, V.D.; Moy, S.S.; Vetreno, R.P.; Coleman, L.G., Jr. Adolescent Binge Alcohol Enhances Early Alzheimer’s Disease Pathology in Adulthood Through Proinflammatory Neuroimmune Activation. Front. Pharmacol. 2022, 13, 884170. [Google Scholar] [CrossRef]


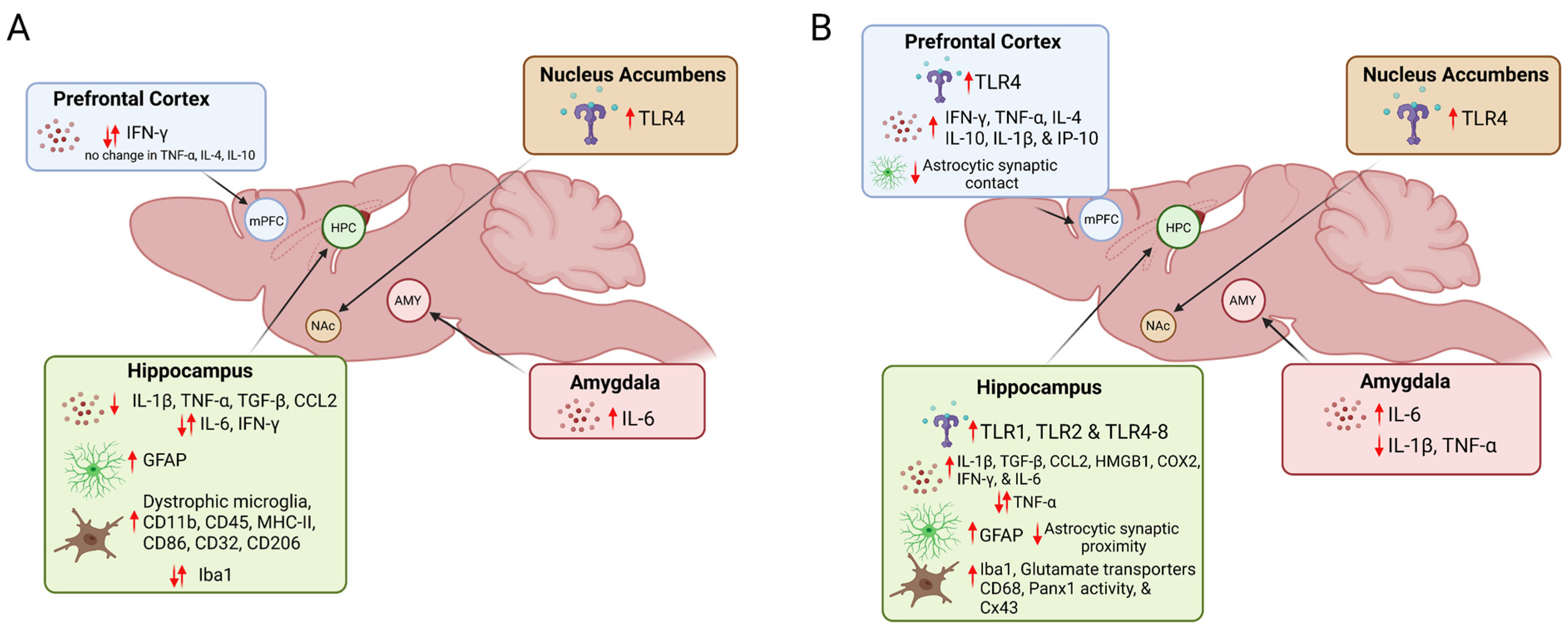{kind=link}
| Duration | Administration | Species/Strain | Toll-like Receptor Effects | Brain Region |
|---|---|---|---|---|
| Acute | ||||
| PND 22–25 | Intragastric gavage (0.5 g/kg–3.5 g/kg/day) | Male & Female Balb/c mice | ↑ TLR4 † | Nucleus Accumbens |
| Chronic | ||||
| PND 28–42 | Operant self-administration (~3.3 g/kg/day) | Male & Female Wistar Rats | ↑ TLR4 * | Medial prefrontal cortex & Hippocampus |
| PND 25–55 [16] | Intragastric gavage (5 g/kg/day) | Male Wistar Rats | ↑ TLR1, ↑ TLR2, -- TLR3, ↑ TLR4, ↑ TLR5, ↑ TLR6, ↑ TLR7, ↑ TLR8, -- TLR9 | Hippocampus |
| PND ~30–51 [20] | Free access self-administration (~9.6 g/kg/day) | Male Wistar Rats | ↑ TLR4 | Hippocampus |
| Duration | Administration | Species/Strain | Cytokine & Chemokine Effects | Brain Region | |
|---|---|---|---|---|---|
| Acute | Cytokines | Chemokines | |||
| PND 33–36 | Intragastric gavage (~12 g/kg/day) | Male Sprague Dawley Rats | ↓ IL-1β, ↓ IL-6, ↓ TNF-α, ↓ TGF-β | ↓ CCL2 | Hippocampus & Entorhinal Cortex |
| PND 32–33 [24] | Intraperitoneal (3.5–4 g/kg) | Male & Female Sprague Dawley Rats | ↑IL-6 | - | Hippocampus & Amygdala |
| PND 30–32 | Intragastric gavage (3 or 6 g/kg/day) | Male Wistar Rats | ↑↓ IFN-γ †, -- TNF-α, IL-4, or IL-10 | - | Prefrontal Cortex & Hippocampus |
| Chronic | |||||
| PND 29–41 | Intraperitoneal (1.25 g/kg) | Male Oncins France-1 Mice | ↑ IL-6 | ↑ CX3CL1 | Striatum |
| ~PND 30–56 | Intragastric gavage (3 or 6 g/kg/day) | Male Wistar Rats | ↑ IFN-γ, ↑ TNF-α, ↑ IL-4 (PFC), ↑ IL-10 (PFC) | - | Prefrontal Cortex & Hippocampus |
| PND 25–54 | Intragastric gavage (5 g/kg/day) | Male Wistar Rats | - | ↑ CCL2, ↑ HMGB1, ↑ COX2 | Hippocampus |
| PND 30–52 | Intragastric gavage (5 g/kg/day) | Male & Female Sprague Dawley Rats | -- TGF-β1, or IL-10, ↑ TNF-α *, ↑ IL-1β * | - | Hippocampus |
| PND 25–55 [16] | Intragastric gavage (5 g/kg/day) | Male Wistar Rats | ↑ TNF-α, -- IL-4, TGF-β, or IL-10 | ↑ HMGB1 | Whole Brain |
| PND 28–48 [31] | Intraperitoneal (4 g/kg/day) | Male & Female Sprague Dawley Rats | ↓ TNF-α, ↑ IL-6, ↑ IL-1β ** (Hipp), ↓ IL-1β * (Amy) | - | Hippocampus & Amygdala |
| PND ~30–51 [20] | Free access self-administration (~9.6 g/kg/day) | Male Wistar Rats | ↑ IL-1β, ↑ TNF-α, -- IL-6 | - | Hippocampus |
| PND 25–38 | Intraperitoneal (3.0 g/kg) | Male Sprague Dawley Rats | ↑ IL-1β, ↑ TNF-α, ↑ IL-6 | - | Hippocampus |
| PND 22–58 | Ethanol Vapor (14 h/day) | Male Wistar Rats | ↑ IL-1β, -- IL-4, IL-5, IL-13, IL-1α, IL12, IL-17, IL-18, IL-10, or TNF-α | ↑ IP-10, -- CX3CL1, CCL11, LIX, or RANTES | Frontal Cortex |
| Duration | Administration | Species/Strain | Glial Effects | Brain Region | |
|---|---|---|---|---|---|
| Acute | Astrocytes | Microglia | |||
| PND 35–39 | Intragastric gavage (3 g/kg/day) | Female Wistar Rats | ↑ GFAP | - | Hippocampus |
| ~PND 35–39 | Intragastric gavage (5 g/kg/day) | Sprague Dawley Rats # | -- GFAP | ↑ Iba1 | Hippocampus |
| PND 35–38 [39] | Intragastric gavage (5 g/kg) | Male Sprague-Dawley rats | - | ↓ Iba1, ↑ Dystrophic microglia | Hippocampus, Perirhinal & Entorhinal Cortices |
| PND 33–36 | Intragastric gavage (5 g/kg) | Male Sprague-Dawley rats | - | ↑ CD11b, ↑ CD45, ↑ MHC-II, ↑ CD86 ↑ CD32, ↑ CD206 | Hippocampus and Entorhinal Cortex |
| PND 40–42 | Intraperitoneal (3 g/kg) | Male Sprague-Dawley rats | -- GFAP | -- Iba1, -- Microglia TLR4 expression | Hippocampus |
| Chronic | |||||
| ~PND 42–74 | DID self-administration (~3.5 g/kg/day) | Male & Female C57BL/6 N mice | - | ↑ Iba1 | Hippocampus |
| ~PND 28–46 | DID self-administration # | Male C57BL/6 Mice | ↑ GFAP | - | Hippocampus |
| PND 31–46 | Intragastric gavage (5 g/kg/day) | Male & Female Sprague Dawley Rats | ↑ GFAP | ↑ Iba1 ** | Hippocampus |
| ~PND 30–58 | Intragastric gavage (1–2 g/kg/day) | Male Sprague Dawley Rats | ↑ GFAP (Hipp/HT), -- GFAP (PFC) | - | Hippocampus, Hypothalamus, Prefrontal Cortex |
| PND 24–55 | Free access self-administration # | Male Syrian Mice | ↑ GFAP | - | Hippocampus |
| PND 28–50 | Intragastric gavage (5 g/kg) | Male Sprague Dawley Rats | ↓ astrocytic-synaptic contact, -- GLT-1, GLAST, α2-δ1, or PSD-95 | - | Medial prefrontal cortex |
| PND 30–46 | Intragastric gavage (5 g/kg) | Male Sprague Dawley Rats | During Withdrawal-- Astrocyte volume or synaptic contact | - | Medial prefrontal cortex, Orbitofrontal cortex, Anterior cingulate cortex |
| Persisting Effects-- Astrocyte Volume ↓ astrocytic-synaptic contact (ACC/OFC) | |||||
| ~PND 32–84 (start & duration varied) | DID self-administration (~2.5 g/kg/day) | Male C57BL/6 Mice | ↓ Astrocytic CB1 and number of processes ↑ Area | - | Hippocampus |
| PND 31–46 | Intragastric gavage (5 g/kg) | Male Sprague Dawley Rats | ↓ Astrocytic-synaptic proximity | - | Hippocampus |
| PND 25–38 | Intraperitoneal (3 g/kg) | Male Sprague Dawley Rats | ↑ Cx43 and Panx1 activity ↑ Arborization | - | Hippocampus |
| PND 35 -58 | Intragastric gavage (3 g/kg/day) | Female Wistar Rats | ↑ GFAP | - | Hippocampus |
| PND 30–46 | Intragastric gavage (5 g/kg) | Male & Female Sprague Dawley Rats | ↑ GLT-1 ↑ xCT and GLAST * | - | Hippocampus |
| PND 28–55 | Intragastric gavage (5 g/kg) | Male Wistar Rats | - | ↑ Iba1 and CD68 | Hippocampus |
| PND 28–49 | Free access self-administration (~9.6 g/kg/day) | Male Wistar Rats | - | ↑ Iba1 | Hippocampus |
| ~PND 25–53 | Free access self-administration (~13.5 g/kg/day) | Male C57BL/6 Mice | - | ↑ CD68 | Dorsal horn, amygdala, anterior cingulate cortex, medullary raphe, thalamus, & hypothalamus |
| PND 28–43 | Intragastric gavage (3.5 g/kg) | Male C57BL/6 Mice | - | ↓ Iba1 | Hippocampus |
| PND 28–42 | Operant self-administration (~3.3 g/kg/day) | Male & Female Wistar Rats | - | ↑ Ibal, ↓ Ramified ** | Hippocampus & prefrontal cortex |
| PND 22–58 | Ethanol Vapor (14 h/day) | Male Wistar Rats | - | ↑ Iba1 | Amygdala, frontal cortex, hippocampus, and substantia nigra |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwachukwu, K.N.; Mohammed, H.E.; Mebane, D.R.; Barber, A.W.; Swartzwelder, H.S.; Marshall, S.A. Acute and Chronic Ethanol Effects during Adolescence on Neuroimmune Responses: Consequences and Potential Pharmacologic Interventions. Cells 2023, 12, 1423. https://doi.org/10.3390/cells12101423
Nwachukwu KN, Mohammed HE, Mebane DR, Barber AW, Swartzwelder HS, Marshall SA. Acute and Chronic Ethanol Effects during Adolescence on Neuroimmune Responses: Consequences and Potential Pharmacologic Interventions. Cells. 2023; 12(10):1423. https://doi.org/10.3390/cells12101423
Chicago/Turabian StyleNwachukwu, Kala N., Hassan E. Mohammed, DaQuan R. Mebane, Andrew W. Barber, H. Scott Swartzwelder, and S. Alex Marshall. 2023. "Acute and Chronic Ethanol Effects during Adolescence on Neuroimmune Responses: Consequences and Potential Pharmacologic Interventions" Cells 12, no. 10: 1423. https://doi.org/10.3390/cells12101423