
Dysregulated PI3K Signaling in B Cells of CVID Patients

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
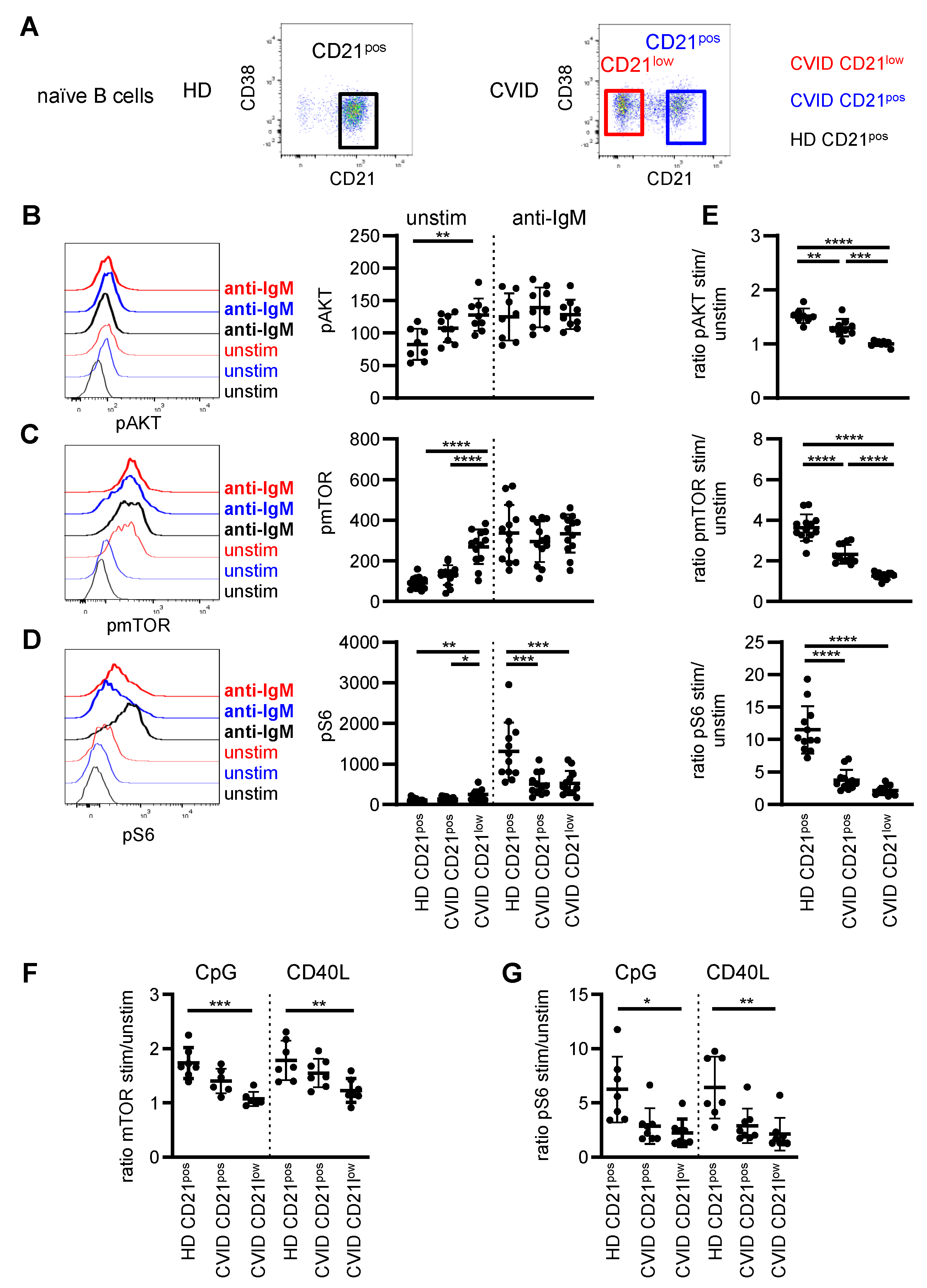
3.1. AKT, mTOR and S6 Phosphorylation in B-Cell Subpopulations of CVID Patients
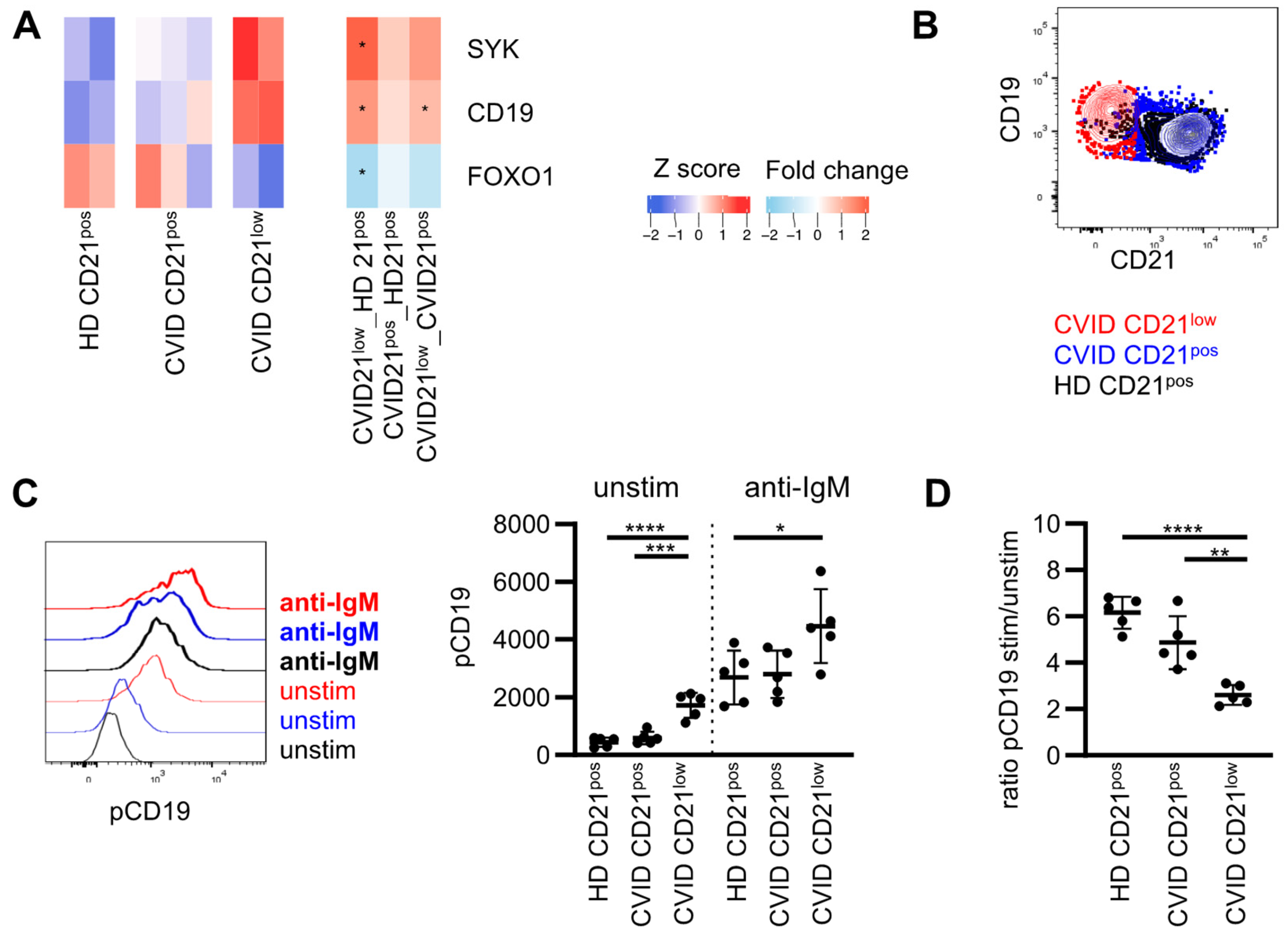
3.2. Expression of Molecules Involved in PI3K Signaling
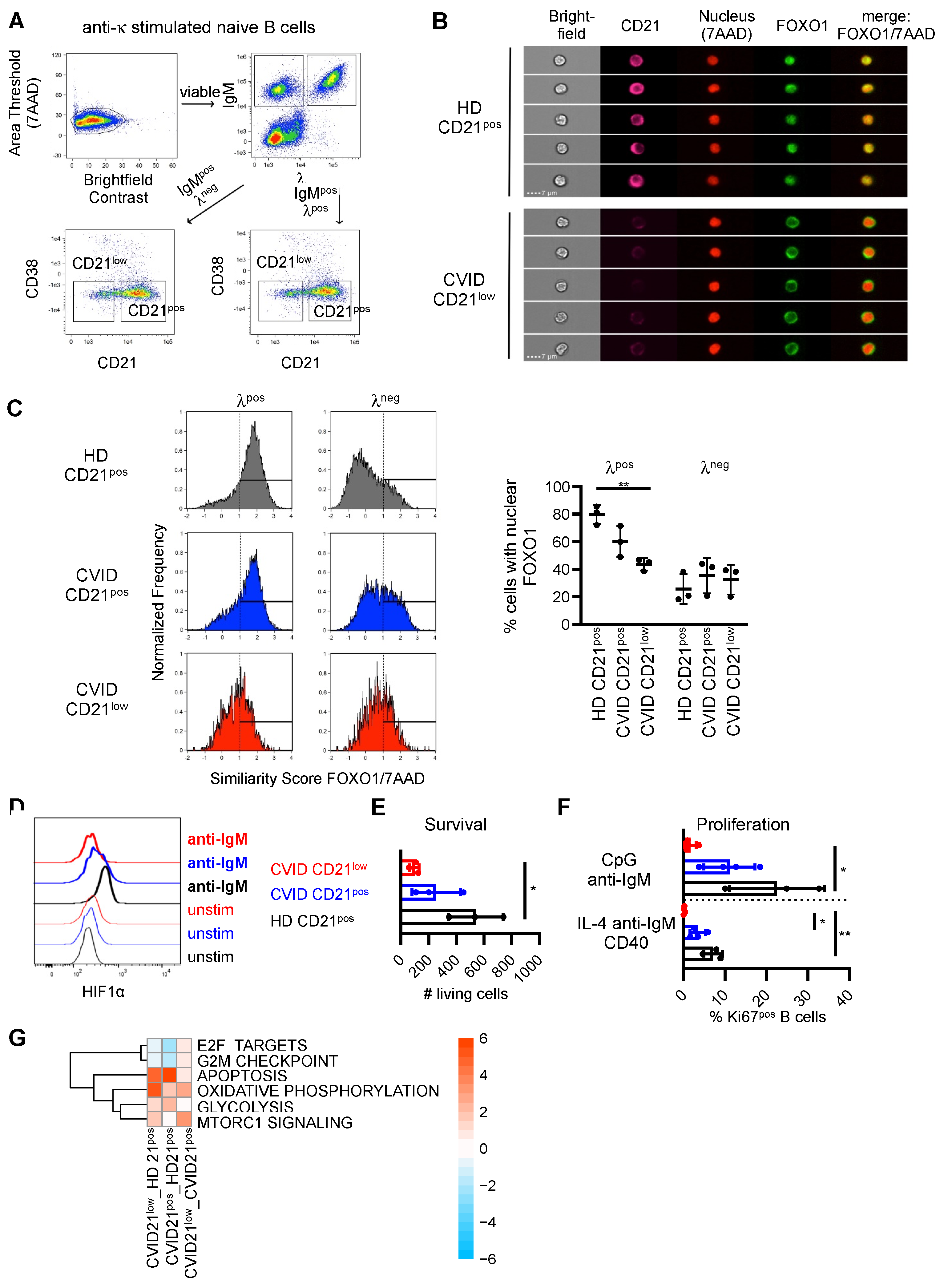
3.3. Downstream Signaling und Target Genes
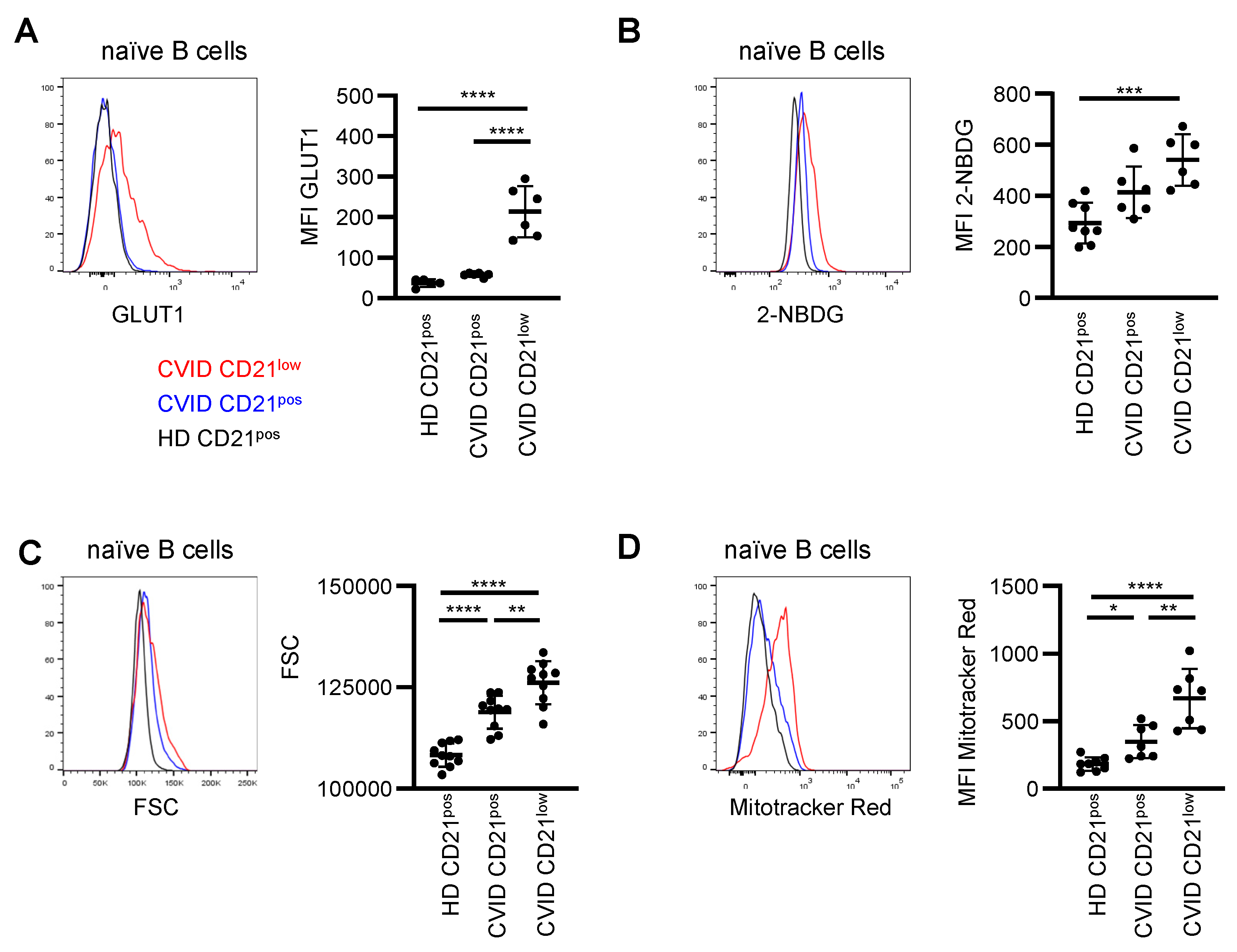
3.4. Metabolic Characteristics
3.5. Correlation with Clinical and Immunological Features
3.6. Delineation from Activated PI3Kδ Syndrome (APDS)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seidel, M.G.; Kindle, G.; Gathmann, B.; Quinti, I.; Buckland, M.; Van Montfrans, J.; Scheible, R.; Rusch, S.; Gasteiger, L.M.; Grimbacher, B.; et al. The European Society for Immunodeficiencies (ESID) Registry Working Definitions for the Clinical Diagnosis of Inborn Errors of Immunity. J. Allergy Clin. Immunol. Pract. 2019, 7, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Chapel, H.; Lucas, M.; Lee, M.; Bjorkander, J.; Webster, D.; Grimbacher, B.; Fieschi, C.; Thon, V.; Abedi, M.R.; Hammarstrom, L. Common Variable Immunodeficiency Disorders: Division into Distinct Clinical Phenotypes. Blood 2008, 112, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham-Rundles, C. The Many Faces of Common Variable Immunodeficiency. Hematology/the Education Program of the American Society of Hematology. Am. Soc. Hematol. Educ. Program 2012, 2012, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.-E.; Cunningham-Rundles, C. Non-Infectious Complications of Common Variable Immunodeficiency: Updated Clinical Spectrum, Sequelae, and Insights to Pathogenesis. Front. Immunol. 2020, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Mouillot, G.; Carmagnat, M.; Gerard, L.; Garnier, J.L.; Fieschi, C.; Vince, N.; Karlin, L.; Viallard, J.F.; Jaussaud, R.; Boileau, J.; et al. B-Cell and T-Cell Phenotypes in CVID Patients Correlate with the Clinical Phenotype of the Disease. J. Clin. Immunol. 2010, 30, 746–755. [Google Scholar] [CrossRef]
- Boileau, J.; Mouillot, G.; Gérard, L.; Carmagnat, M.; Rabian, C.; Oksenhendler, E.; Pasquali, J.-L.; Korganow, A.-S. Autoimmunity in Common Variable Immunodeficiency: Correlation with Lymphocyte Phenotype in the French DEFI study. J. Autoimmun. 2011, 36, 25–32. [Google Scholar] [CrossRef]
- Wehr, C.; Kivioja, T.; Schmitt, C.; Ferry, B.; Witte, T.; Eren, E.; Vlkova, M.; Hernandez-Gonzalez, M.; Detkova, D.; Bos, P.R.; et al. The EUROclass Trial: Defining Subgroups in Common Variable Immunodeficiency. Blood 2008, 111, 77–85. [Google Scholar] [CrossRef]
- Warnatz, K.; Wehr, C.; Drager, R.; Schmidt, S.; Eibel, H.; Schlesier, M.; Peter, H.H. Expansion of cd19(hi)cd21(lo/neg) B Cells in Common Variable Immunodeficiency (cvid) Patients with Autoimmune Cytopenia. Immunobiology 2002, 206, 502–513. [Google Scholar] [CrossRef]
- Moir, S.; Malaspina, A.; Ogwaro, K.M.; Donoghue, E.T.; Hallahan, C.W.; Ehler, L.A.; Liu, S.; Adelsberger, J.; Lapointe, R.; Hwu, P.; et al. HIV-1 Induces Phenotypic and Functional Perturbations of B Cells in Chronically Infected Individuals. Proc. Natl. Acad. Sci. USA 2001, 98, 10362–10367. [Google Scholar] [CrossRef] [Green Version]
- Charles, E.D.; Brunetti, C.; Marukian, S.; Ritola, K.D.; Talal, A.H.; Marks, K.; Jacobson, I.M.; Rice, C.M.; Dustin, L.B. Clonal B Cells in Patients with Hepatitis C Virus–Associated Mixed Cryoglobulinemia Contain an Expanded Anergic CD21low B-Cell Subset. Blood 2011, 117, 5425–5437. [Google Scholar] [CrossRef]
- Weiss, G.E.; Crompton, P.D.; Li, S.; Walsh, L.A.; Moir, S.; Traore, B.; Kayentao, K.; Ongoiba, A.; Doumbo, O.K.; Pierce, S.K. Atypical Memory B Cells Are Greatly Expanded in Individuals Living in a Malaria-Endemic Area. J. Immunol. 2009, 183, 2176–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnardi, I.; Ng, Y.-S.; Menard, L.; Meyers, G.; Saadoun, D.; Srdanovic, I.; Samuels, J.; Berman, J.; Buckner, J.H.; Cunningham-Rundles, C.; et al. Complement Receptor 2/CD21−Human Naive B Cells Contain Mostly Autoreactive Unresponsive Clones. Blood 2010, 115, 5026–5036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehr, C.; Eibel, H.; Masilamani, M.; Illges, H.; Schlesier, M.; Peter, H.-H.; Warnatz, K. A New CD21low B Cell Population in the Peripheral Blood of Patients with SLE. Clin. Immunol. 2004, 113, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Zumaquero, E.; Stone, S.L.; Scharer, C.D.; Jenks, S.A.; Nellore, A.; Mousseau, B.; Rosal-Vela, A.; Botta, D.; Bradley, J.E.; Wojciechowski, W.; et al. Ifngamma Induces Epigenetic Programming of Human t-bet(hi) B Cells and Promotes tlr7/8 and il-21 Induced Differentiation. Elife 2019, 8, e41641. [Google Scholar] [CrossRef] [PubMed]
- Tipton, C.M.; Fucile, C.F.; Darce, J.; Chida, A.S.; Ichikawa, T.; Gregoretti, I.; Schieferl, S.M.; Hom, J.; Jenks, S.A.; Feldman, R.J.; et al. Diversity, Cellular Origin and Autoreactivity of Antibody-Secreting Cell Population Expansions in Acute Systemic Lupus Erythematosus. Nat. Immunol. 2015, 16, 755–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenks, S.A.; Cashman, K.S.; Zumaquero, E.; Marigorta, U.M.; Patel, A.V.; Wang, X.; Tomar, D.; Woodruff, M.C.; Simon, Z.; Bugrovsky, R.; et al. Distinct Effector B Cells Induced by Unregulated Toll-like Receptor 7 Contribute to Pathogenic Responses in Systemic Lupus Erythematosus. Immunity 2018, 49, 725–739.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, B.; Strohmeier, V.; Harder, I.; Unger, S.; Payne, K.J.; Andrieux, G.; Boerries, M.; Felixberger, P.T.; Landry, J.J.M.; Nieters, A.; et al. The Expansion of Human T-Bet High CD21 Low B Cells is T Cell Dependent. Sci. Immunol. 2021, 6. [Google Scholar] [CrossRef]
- Knox, J.J.; Kaplan, D.E.; Betts, M.R. T-Bet-Expressing B Cells during HIV and HCV Infections. Cell. Immunol. 2017, 321, 26–34. [Google Scholar] [CrossRef]
- Obeng-Adjei, N.; Portugal, S.; Holla, P.; Li, S.; Sohn, H.; Ambegaonkar, A.; Skinner, J.; Bowyer, G.; Doumbo, O.K.; Traore, B.; et al. Malaria-Induced Interferon-Gamma Drives the Expansion of Tbethi Atypical Memory B Cells. PLoS Pathog. 2017, 13, e1006576. [Google Scholar] [CrossRef] [Green Version]
- Rakhmanov, M.; Keller, B.; Gutenberger, S.; Foerster, C.; Hoenig, M.; Driessen, G.; van der Burg, M.; Van Dongen, J.; Wiech, E.; Visentini, M.; et al. Circulating CD21low B Cells in Common Variable Immunodeficiency Resemble Tissue Homing, Innate-Like B Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 13451–13456. [Google Scholar] [CrossRef] [Green Version]
- Portugal, S.; Tipton, C.M.; Sohn, H.; Kone, Y.; Wang, J.; Li, S.; Skinner, J.; Virtaneva, K.; Sturdevant, D.E.; Porcella, S.F.; et al. Malaria-Associated Atypical Memory B Cells Exhibit Markedly Reduced B Cell Receptor Signaling and Effector Function. eLife 2015, 4, e07218. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, M.W.; Dooley, M.A.; Hogan, S.L.; Anolik, J.; Looney, J.; Sanz, I.; Clarke, S.H. A Novel Subset of Memory B Cells is Enriched in Autoreactivity and Correlates with Adverse Outcomes in SLE. Clin. Immunol. 2008, 126, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, B.; Stumpf, I.; Strohmeier, V.; Usadel, S.; Verhoeyen, E.; Eibel, H.; Warnatz, K. High SYK Expression Drives Constitutive Activation of CD21low B Cells. J. Immunol. 2017, 198, 4285–4292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, B.; Cseresnyes, Z.; Stumpf, I.; Wehr, C.; Fliegauf, M.; Bulashevska, A.; Usadel, S.; Grimbacher, B.; Rizzi, M.; Eibel, H.; et al. Disturbed Canonical Nuclear Factor of Kappa Light Chain Signaling in B Cells of Patients with Common Variable ImmunoDeficiency. J. Allergy Clin. Immunol. 2017, 139, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Kardava, L.; Sohn, H.; Youn, C.; Austin, J.W.; Wang, W.; Buckner, C.M.; Justement, J.S.; Melson, V.A.; Roth, G.E.; Hand, M.A.; et al. IgG3 Regulates Tissue-Like Memory B Cells in HIV-Infected Individuals. Nat. Immunol. 2018, 19, 1001–1012. [Google Scholar] [CrossRef]
- Freudenhammer, M.; Voll, R.E.; Binder, S.C.; Keller, B.; Warnatz, K. Naive- and Memory-like CD21low B Cell Subsets Share Core Phenotypic and Signaling Characteristics in Systemic Autoimmune Disorders. J. Immunol. 2020, 205, 2016–2025. [Google Scholar] [CrossRef]
- Foerster, C.; Voelxen, N.; Rakhmanov, M.; Keller, B.; Gutenberger, S.; Goldacker, S.; Thiel, J.; Feske, S.; Peter, H.-H.; Warnatz, K. B Cell Receptor-Mediated Calcium Signaling Is Impaired in B Lymphocytes of Type Ia Patients with Common Variable Immunodeficiency. J. Immunol. 2010, 184, 7305–7313. [Google Scholar] [CrossRef] [Green Version]
- Burton, A.R.; Pallett, L.J.; McCoy, L.E.; Suveizdytė, K.; Amin, O.E.; Swadling, L.; Alberts, E.; Davidson, B.R.; Kennedy, P.T.; Gill, U.S.; et al. Circulating and Intrahepatic Antiviral B Cells are Defective in Hepatitis, B.J. Clin. Investig. 2018, 128, 4588–4603. [Google Scholar] [CrossRef] [Green Version]
- Ramadani, F.; Bolland, D.J.; Garcon, F.; Emery, J.L.; Vanhaesebroeck, B.; Corcoran, A.E.; Okkenhaug, K. The Pi3k Isoforms P110alpha and P110delta are Essential for Pre-B Cell Receptor Signaling and B Cell Development. Sci. Signal. 2010, 3, ra50. [Google Scholar] [CrossRef] [Green Version]
- Okkenhaug, K. Signaling by the Phosphoinositide 3-Kinase Family in Immune Cells. Annu. Rev. Immunol. 2013, 31, 675–704. [Google Scholar] [CrossRef] [Green Version]
- Omori, S.A.; Cato, M.H.; Anzelon-Mills, A.; Puri, K.D.; Shelef, M.; Calame, K.; Rickert, R.C. Regulation of Class-Switch Recombination and Plasma Cell Differentiation by Phosphatidylinositol 3-Kinase Signaling. Immunity 2006, 25, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The Pi3k Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilancio, A.; Okkenhaug, K.; Camps, M.; Emery, J.L.; Ruckle, T.; Rommel, C.; Vanhaesebroeck, B. Key Role of the P110delta Isoform of Pi3k in B-Cell Antigen and il-4 Receptor Signaling: Comparative Analysis of Genetic and Pharmacologic Interference with P110delta Function in B Cells. Blood 2006, 107, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Santos, C.J.; Uzel, G.; Rosenzweig, S.D. PI3K Pathway Defects Leading to Immunodeficiency and Immune Dysregulation. J. Allergy Clin. Immunol. 2019, 143, 1676–1687. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Deenick, E.K.; Tangye, S.G. Phosphatidylinositol 3-Kinase Signaling and Immune Regulation: Insights into Disease Pathogenesis and Clinical Implications. Expert Rev. Clin. Immunol. 2021, 17, 905–914. [Google Scholar] [CrossRef]
- Lucas, C.L.; Kuehn, H.S.; Zhao, F.; Niemela, J.E.; Deenick, E.K.; Palendira, U.; Avery, D.T.; Moens, L.; Cannons, J.L.; Biancalana, M.; et al. Dominant-Activating Germline Mutations in the Gene Encoding the Pi(3)k Catalytic Subunit P110delta Result in T Cell Senescence and Human Immunodeficiency. Nat. Immunol 2014, 15, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Angulo, I.; Vadas, O.; Garcon, F.; Banham-Hall, E.; Plagnol, V.; Leahy, T.R.; Baxendale, H.; Coulter, T.; Curtis, J.; Wu, C.; et al. Phosphoinositide 3-Kinase Delta Gene Mutation Predisposes to Respiratory Infection and Airway Damage. Science 2013, 342, 866–871. [Google Scholar] [CrossRef] [Green Version]
- Asano, T.; Okada, S.; Tsumura, M.; Yeh, T.W.; Mitsui-Sekinaka, K.; Tsujita, Y.; Ichinose, Y.; Shimada, A.; Hashimoto, K.; Wada, T.; et al. Enhanced Akt Phosphorylation of Circulating B Cells in Patients with Activated Pi3kdelta Syndrome. Front. Immunol. 2018, 9, 568. [Google Scholar] [CrossRef] [Green Version]
- Avery, D.T.; Kane, A.; Nguyen, T.; Lau, A.; Nguyen, A.; Lenthall, H.; Payne, K.; Shi, W.; Brigden, H.; French, E.; et al. Germline-Activating Mutations in PIK3CD Compromise B Cell Development and Function. J. Exp. Med. 2018, 215, 2073–2095. [Google Scholar] [CrossRef]
- Olbrich, P.; Lorenz, M.; Cura Daball, P.; Lucena, J.M.; Rensing-Ehl, A.; Sanchez, B.; Fuhrer, M.; Camacho-Lovillo, M.; Melon, M.; Schwarz, K.; et al. Activated Pi3kdelta Syndrome Type 2: Two Patients, a Novel Mutation, and Review of the Literature. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2016, 27, 640–644. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Friedman, M.S.; Shedden, K.; Hankenson, K.D.; Woolf, P.J. GAGE: Generally Applicable Gene Set Enrichment for Pathway Analysis. BMC Bioinform. 2009, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischer, S.J.; Giesecke, C.; Mei, H.E.; Lipsky, P.E.; Daridon, C.; Dörner, T. Increased Frequency of a Unique Spleen Tyrosine Kinase Bright Memory B Cell Population in Systemic Lupus Erythematosus. Arthritis Rheumatol. 2014, 66, 3424–3435. [Google Scholar] [CrossRef] [PubMed]
- Tedder, T.F.; Inaoki, M.; Sato, S. The CD19–CD21 Complex Regulates Signal Transduction Thresholds Governing Humoral Immunity and Autoimmunity. Immunity 1997, 6, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, L.; Sasaki, Y.; Calado, D.P.; Zhang, B.; Paik, J.H.; DePinho, R.A.; Kutok, J.L.; Kearney, J.F.; Otipoby, K.L.; Rajewsky, K. PI3 Kinase Signals BCR-Dependent Mature B Cell Survival. Cell 2009, 139, 573–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limon, J.J.; Fruman, D.A. Akt and mTOR in B Cell Activation and Differentiation. Front. Immunol. 2012, 3, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beitz, L.O.; Fruman, D.A.; Kurosaki, T.; Cantley, L.C.; Scharenberg, A.M. SYK Is Upstream of Phosphoinositide 3-Kinase in B Cell Receptor Signaling. J. Biol. Chem. 1999, 274, 32662–32666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S. CD19 is a Central Response Regulator of B Lymphocyte Signaling Thresholds Governing Autoimmunity. J. Dermatol. Sci. 1999, 22, 1–10. [Google Scholar] [CrossRef]
- Irish, J.M.; Doxie, D.B. High-Dimensional Single-Cell Cancer Biology. Curr. Top. Microbiol. Immunol. 2014, 377, 1–21. [Google Scholar] [PubMed] [Green Version]
- Cortez, V.C.R.; Molina, L.D.P.; Rodriguez-Ubreva, J.; Ciudad, L.; Gomez-Cabrero, D.; Company, C.; Urquiza, J.M.; Tegnér, J.; Rodríguez-Gallego, C.; López-Granados, E.; et al. Monozygotic Twins Discordant for Common variable Immunodeficiency Reveal Impaired DNA Demethylation during Naïve-to-Memory B-Cell Transition. Nat. Commun. 2015, 6, 7335. [Google Scholar] [CrossRef]
- Fleischer, S.J.; Daridon, C.; Fleischer, V.; Lipsky, P.E.; Dörner, T. Enhanced Tyrosine Phosphatase Activity Underlies Dysregulated B-Cell Receptor Signaling and Promotes Survival of Human Lupus B-Cells. Arthritis Rheumatol. 2016, 68, 1210–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmond, R.J.; Emery, J.; Okkenhaug, K.; Zamoyska, R. MAPK, Phosphatidylinositol 3-Kinase, and Mammalian Target of Rapamycin Pathways Converge at the Level of Ribosomal Protein S6 Phosphorylation to Control Metabolic Signaling in CD8 T Cells. J. Immunol. 2009, 183, 7388–7397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyuhas, O. Ribosomal Protein s6 Phosphorylation: Four Decades of Research. Int. Rev. Cell Mol. Biol. 2015, 320, 41–73. [Google Scholar] [PubMed]
- Shlomchik, M.J.; Luo, W.; Weisel, F. Linking Signaling and Selection in the Germinal Center. Immunol. Rev. 2019, 288, 49–63. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Cyster, J.G. Transcriptional Regulation of Memory B Cell Differentiation. Nat. Rev. Immunol. 2020, 21, 209–220. [Google Scholar] [CrossRef]
- Jellusova, J.; Rickert, R.C. The Pi3k Pathway in B Cell Metabolism. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 359–378. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Pallikkuth, S.; Pahwa, S. Metabolic Phenotype of B Cells from Young and Elderly HIV Individuals. Immun. Ageing 2021, 18, 35. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef]
- Zhu, J.; Thompson, C.B. Metabolic Regulation of Cell Growth and Proliferation. Nat. Rev. Mol. Cell Biol. 2019, 20, 436–450. [Google Scholar] [CrossRef]
- Jacobi, A.M.; Reiter, K.; Mackay, M.; Aranow, C.; Hiepe, F.; Radbruch, A.; Hansen, A.; Burmester, G.-R.; Diamond, B.; Lipsky, P.E.; et al. Activated Memory B Cell Subsets Correlate with Disease Activity in Systemic Lupus Erythematosus: Delineation by Expression of CD27, IgD, and CD95. Arthritis Care Res. 2008, 58, 1762–1773. [Google Scholar] [CrossRef]
- Ahye, M.K.H.; Golding, A. Cytoplasmic FOXO1 Identifies a Novel Disease-Activity Associated B Cell Phenotype in SLE. Lupus Sci. Med. 2018, 5, e000296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Jhunjhunwala, S.; Benner, C.; Heinz, S.; Welinder, E.; Mansson, R.; Sigvardsson, M.; Hagman, J.; Espinoza, C.A.; Dutkowski, J.; et al. A Global Network of Transcription Factors, Involving E2A, EBF1 and Foxo1, that Orchestrates B Cell Fate. Nat. Immunol. 2010, 11, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, I.; Zhu, X.; Kharas, M.G.; Chen, J.; Fruman, D.A. Optimal B-Cell Proliferation Requires Phosphoinositide 3-Kinase–Dependent Inactivation of FOXO Transcription Factors. Blood 2004, 104, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Grötsch, B.; Luo, Y.; Knaup, K.X.; Wiesener, M.S.; Chen, X.-X.; Jantsch, J.; Fillatreau, S.; Schett, G.; Bozec, A. Hypoxia-Inducible Factor-1α is a Critical Transcription Factor for IL-10-Producing B Cells in Autoimmune Disease. Nat. Commun. 2018, 9, 251. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.-N.; Ye, Y.-X.; Niu, J.-W.; Li, Y.; Li, X.; You, X.; Chen, H.; Zhao, L.-D.; Zeng, X.-F.; Zhang, F.-C.; et al. Defective PTEN Regulation Contributes to B Cell Hyperresponsiveness in Systemic Lupus Erythematosus. Sci. Transl. Med. 2014, 6, 246ra99. [Google Scholar] [CrossRef]
- Taher, T.E.; Parikh, K.; Flores-Borja, F.; Mletzko, S.; Isenberg, D.A.; Peppelenbosch, M.P.; Mageed, R.A. Protein Phosphorylation and Kinome Profiling Reveal Altered Regulation of Multiple Signaling Pathways in B Lymphocytes from Patients with Systemic Lupus Erythematosus. Arthritis Care Res. 2010, 62, 2412–2423. [Google Scholar] [CrossRef]
- Wu, C.; Fu, Q.; Guo, Q.; Chen, S.; Goswami, S.; Sun, S.; Li, T.; Cao, X.; Chu, F.; Chen, Z.; et al. Lupus-Associated Atypical Memory B Cells are mTORC1-Hyperactivated and Functionally Dysregulated. Ann. Rheum. Dis. 2019, 78, 1090–1100. [Google Scholar] [CrossRef] [Green Version]
- Yazdani, R.; Ganjalikhani-Hakemi, M.; Esmaeili, M.; Abolhassani, H.; Vaeli, S.; Rezaei, A.; Sharifi, Z.; Azizi, G.; Rezaei, N.; Aghamohammadi, A. Impaired Akt phosphorylation in B-Cells of Patients with Common Variable Immunodeficiency. Clin. Immunol. 2017, 175, 124–132. [Google Scholar] [CrossRef]
- Del Pino-Molina, L.; Canizales, J.M.T.; Rodríguez-Pena, R.; López-Granados, E. Evaluation of B-Cell Intracellular Signaling by Monitoring the PI3K-Akt Axis in Patients with Common Variable Immunodeficiency and Activated Phosphoinositide 3-Kinase Delta Syndrome. Cytom. Part B Clin. Cytom. 2020. [Google Scholar] [CrossRef]
- Ersching, J.; Efeyan, A.; Mesin, L.; Jacobsen, J.T.; Pasqual, G.; Grabiner, B.C.; Dominguez-Sola, D.; Sabatini, D.M.; Victora, G.D. Germinal Center Selection and Affinity Maturation Require Dynamic Regulation of mTORC1 Kinase. Immunity 2017, 46, 1045–1058.e6. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.H.; Raybuck, A.L.; Stengel, K.; Wei, M.; Beck, T.C.; Volanakis, E.; Thomas, J.W.; Hiebert, S.; Haase, V.H.; Boothby, M.R. Germinal Centre Hypoxia and Regulation of Antibody Qualities by a Hypoxia Response System. Nature 2016, 537, 234–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Weisel, F.; Shlomchik, M.J. B Cell Receptor and CD40 Signaling Are Rewired for Synergistic Induction of the c-Myc Transcription Factor in Germinal Center B Cells. Immunity 2018, 48, 313–326.e315. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harder, I.; Münchhalfen, M.; Andrieux, G.; Boerries, M.; Grimbacher, B.; Eibel, H.; Maccari, M.E.; Ehl, S.; Wienands, J.; Jellusova, J.; et al. Dysregulated PI3K Signaling in B Cells of CVID Patients. Cells 2022, 11, 464. https://doi.org/10.3390/cells11030464
Harder I, Münchhalfen M, Andrieux G, Boerries M, Grimbacher B, Eibel H, Maccari ME, Ehl S, Wienands J, Jellusova J, et al. Dysregulated PI3K Signaling in B Cells of CVID Patients. Cells. 2022; 11(3):464. https://doi.org/10.3390/cells11030464
Chicago/Turabian StyleHarder, Ina, Matthias Münchhalfen, Geoffroy Andrieux, Melanie Boerries, Bodo Grimbacher, Hermann Eibel, Maria Elena Maccari, Stephan Ehl, Jürgen Wienands, Julia Jellusova, and et al. 2022. "Dysregulated PI3K Signaling in B Cells of CVID Patients" Cells 11, no. 3: 464. https://doi.org/10.3390/cells11030464