
Special Microbial Communities Enhanced the Role of Aged Biochar in Reducing Cd Accumulation in Rice
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Biochar Production
2.2. Experimental Design
2.3. Soil and Plant Analysis
2.4. Rhizosphere Soil DNA Extraction and Sequencing
2.5. Data and Statistical Analysis
3. Results
3.1. Plant Cd Uptake
3.2. Changes in Soil Chemical Properties
3.3. Soil Available Cd Concentration
3.4. Effect of Biochar on the Rrhizosphere Microbial Community
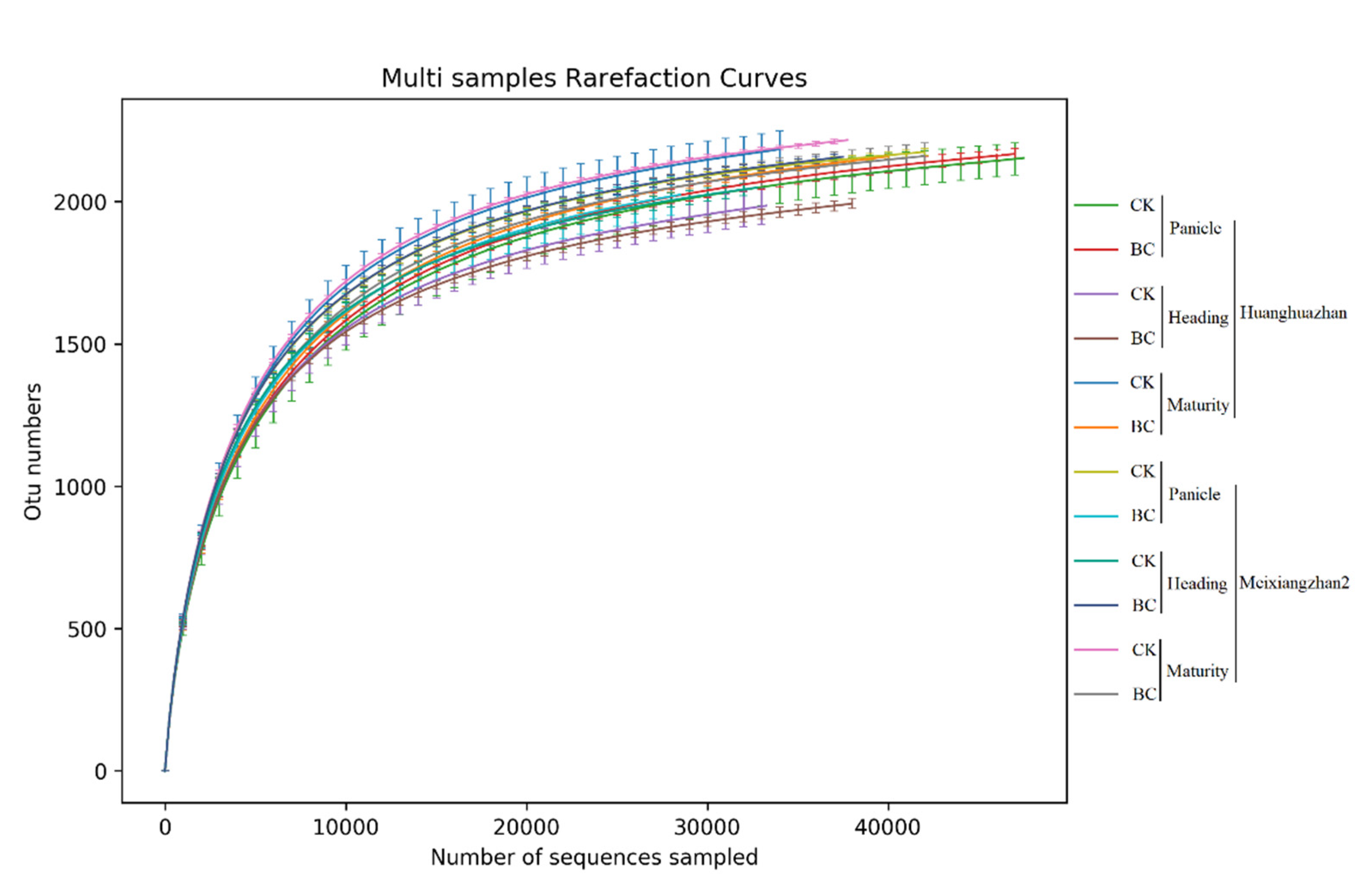
3.4.1. Bacterial Community Diversity and Abundance
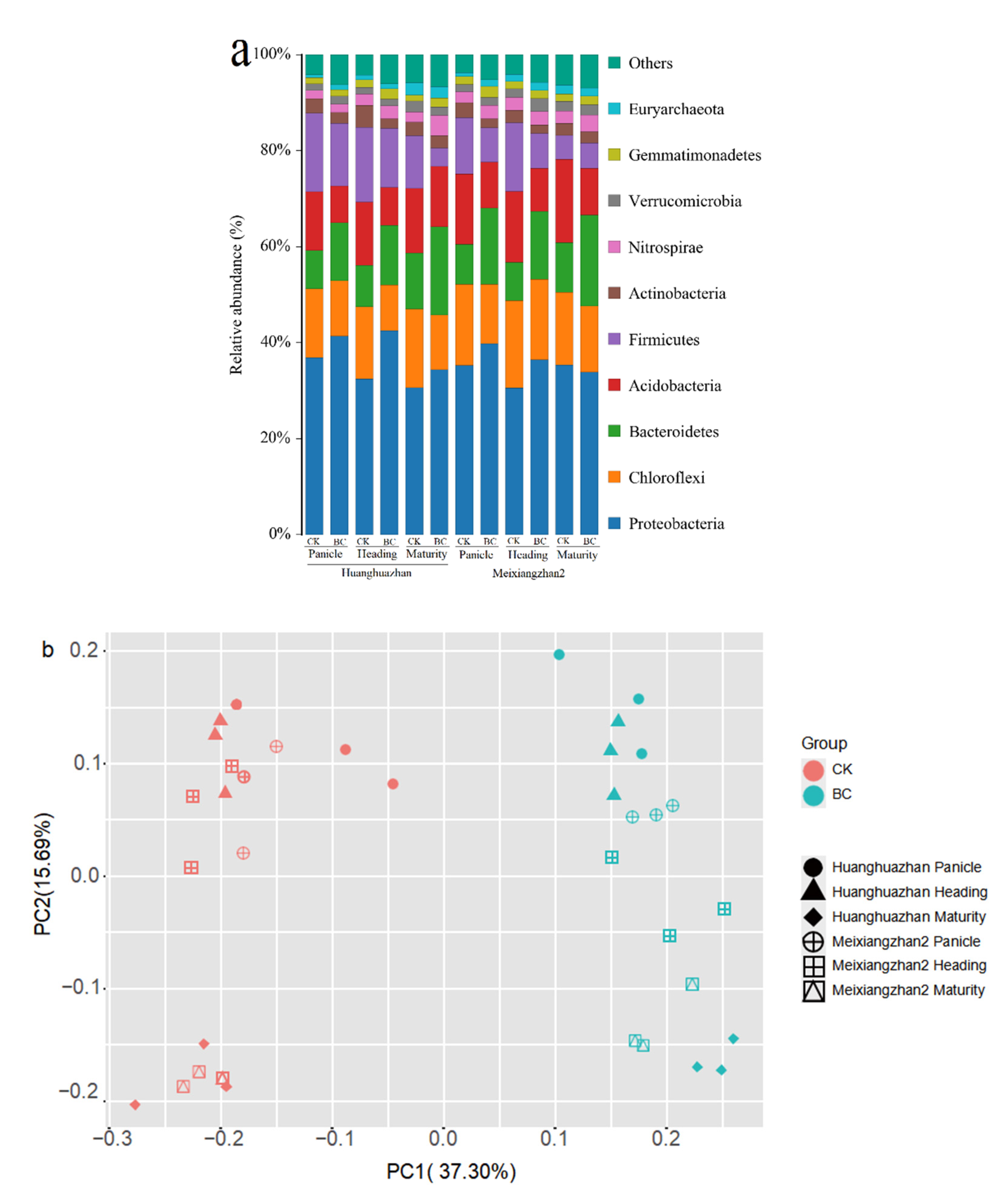
3.4.2. Bacterial Community Structure
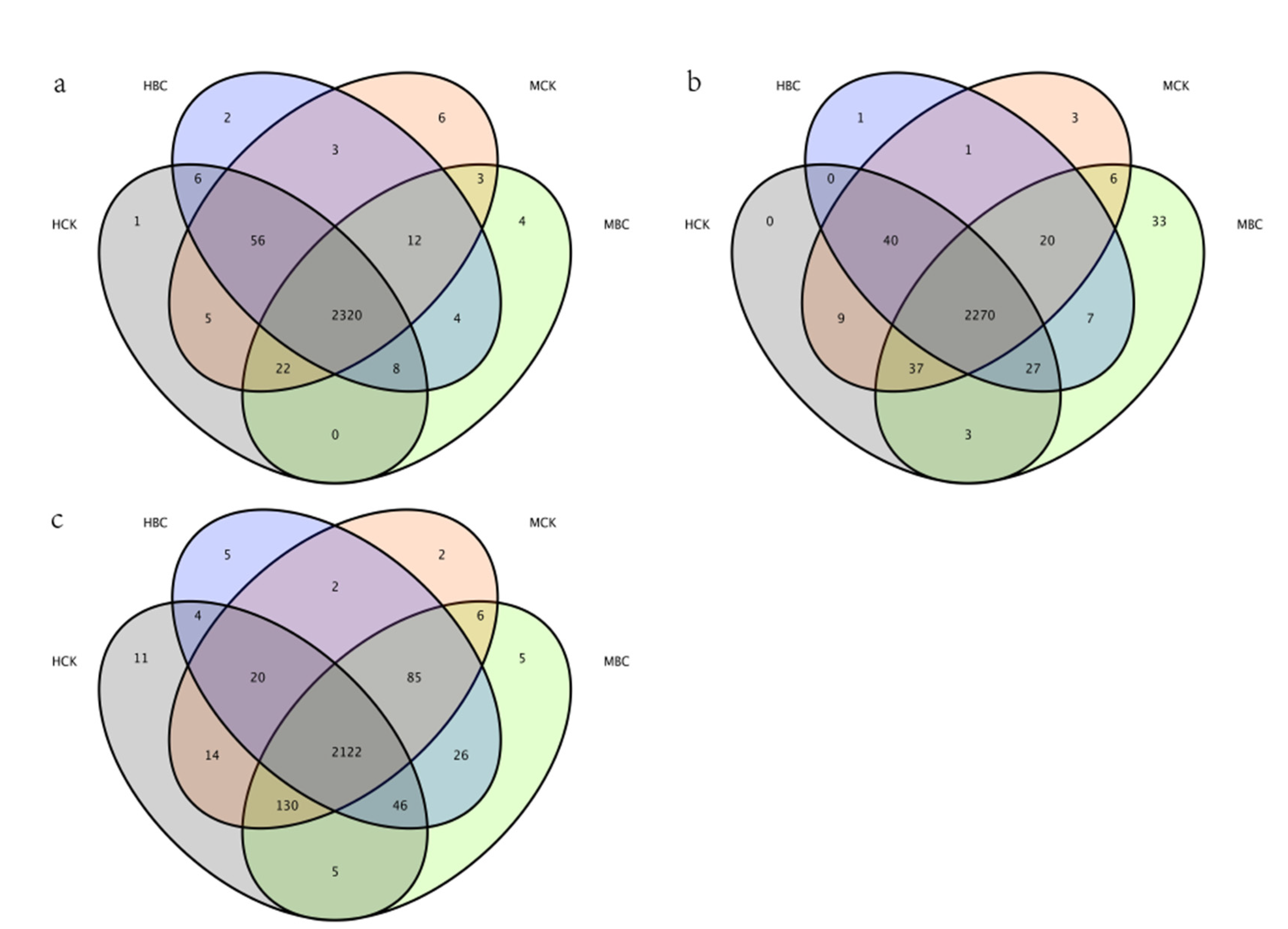
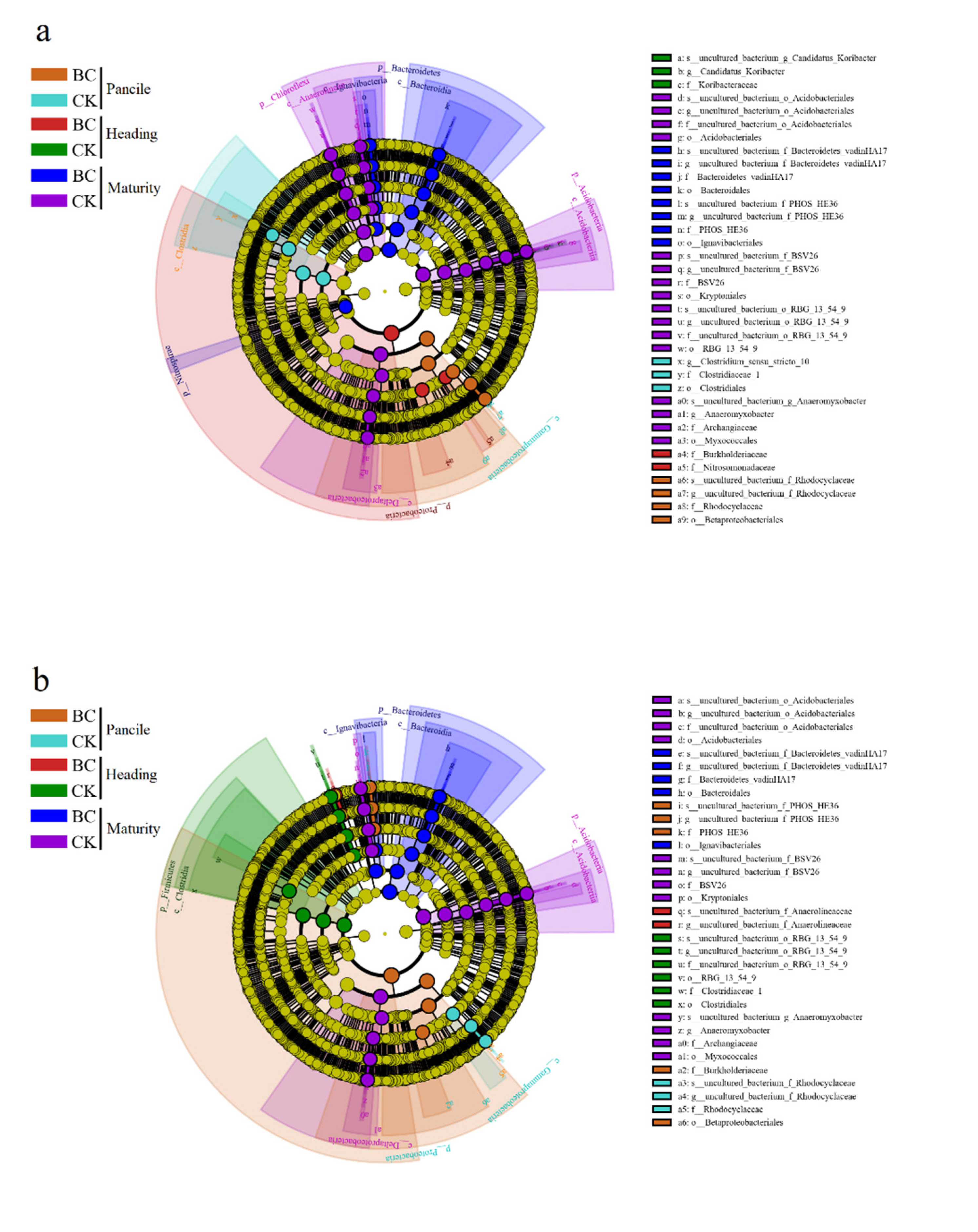
3.4.3. Specific Bacterial Assemblages in the Rhizosphere
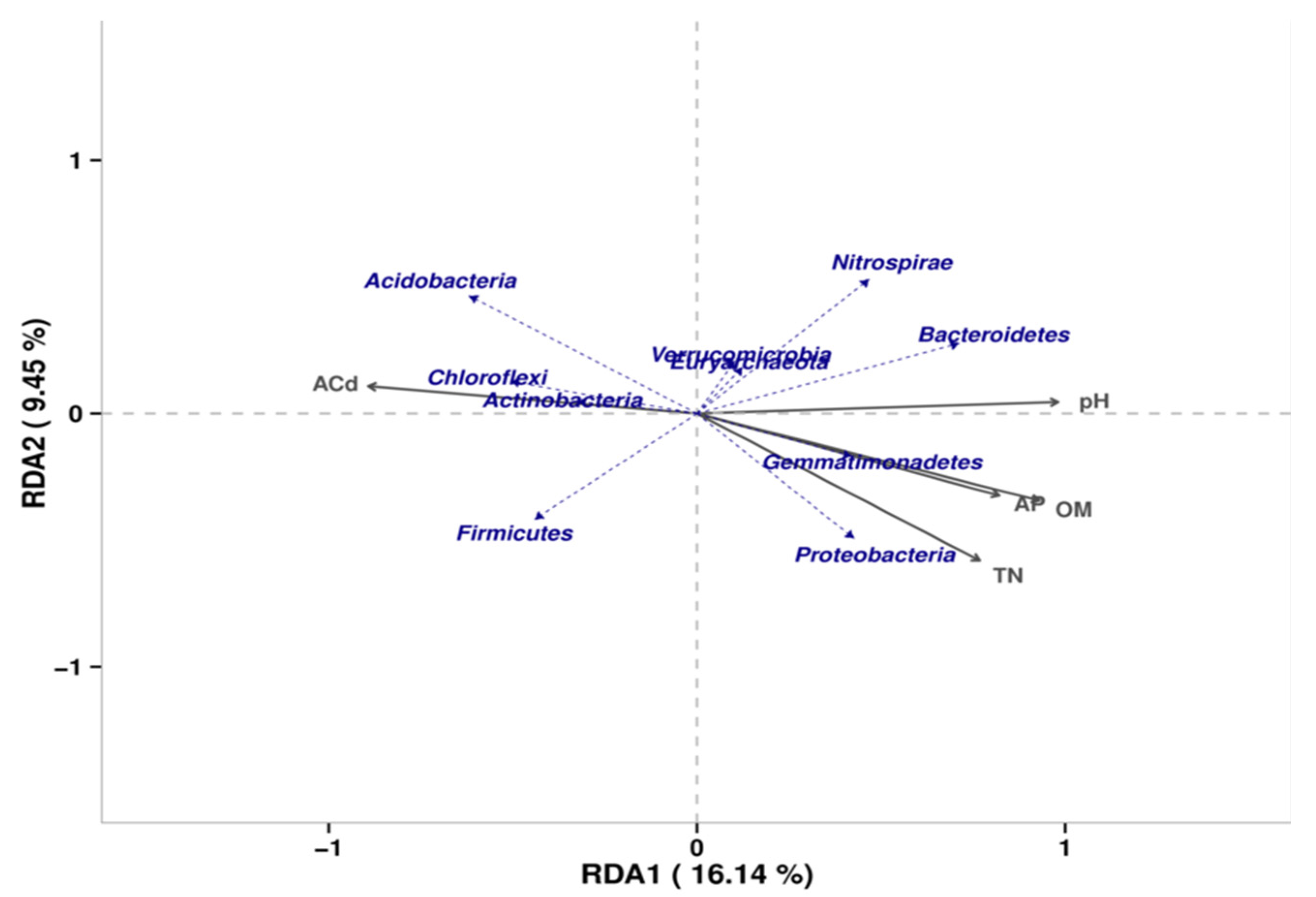
3.4.4. Environmental Factors Associated with the Microbial Community
4. Discussion
4.1. Effects of Biochar Aging on Soil Physicochemical Properties and Available Cd Concentration
4.2. Effects of Biochar Aging on Rhizosphere Microbial Community Structure Composition
4.3. Effects of Aging Biochar on Rice Uptake of Cd
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, H.; Tang, Z.; Wang, P.; Zhao, F. Geographical variations of cadmium and arsenic concentrations and arsenic speciation in Chinese rice. Environ. Pollut. 2018, 238, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Li, D.; Tang, B.; Man, S.; Jia, Y.; Xu, H. Vermicompost and biochar as bio-conditioners to immobilize heavy metal and improve soil fertility on cadmium contaminated soil under acid rain stress. Sci. Total Environ. 2018, 621, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Chen, S.; Liu, J.; Chen, L.; Song, N.; Li, N.; Liu, B. Variation of Cd concentration in various rice cultivars and derivation of cadmium toxicity thresholds for paddy soil by species-sensitivity distribution. J. Integr. Agric. 2015, 14, 1845–1854. [Google Scholar] [CrossRef]
- Jia, X.; Zhou, C.; Dong, S. Effects of Pb2+/Cd2+ on the number of microorganisms in the rhizosphere of winter wheat seedings. J. Shenyang Agric. Univ. 2011, 42, 443–448. [Google Scholar]
- Shi, Z.; Carey, M.; Meharg, C.; Williams, P.N.; Signes-Pastor, A.J.; Triwardhani, E.A.; Pandiangan, F.I.; Campbell, K.; Elliott, C.; Marwa, E.M.; et al. Rice Grain Cadmium Concentrations in the Global Supply-Chain. Expo. Health 2020, 12, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Zhou, S.; Zhou, Y.; Jia, Z.; Guo, T.; Wang, J. Cadmium pollution of soil-rice ecosystems in rice cultivation dominated regions in China: A review. Environ. Pollut. 2021, 280, 116965. [Google Scholar] [CrossRef]
- Cheng, Z.; Shi, J.; He, Y.; Wu, L.; Xu, J. Assembly of root-associated bacterial community in cadmium contaminated soil following five-year consecutive application of soil amendments: Evidences for improved soil health. J. Hazard. Mater. 2022, 426, 128095. [Google Scholar] [CrossRef]
- Shahid, M.; Ali, S.; Shabir, G.; Siddique, M.; Rizwan, M.; Seleiman, M.F.; Afzal, M. Comparing the performance of four macrophytes in bacterial assisted floating treatment wetlands for the removal of trace metals (Fe, Mn, Ni, Pb, and Cr) from polluted river water. Chemosphere 2019, 243, 125353. [Google Scholar] [CrossRef]
- Lehmann, J. A handful of carbon. Nature 2007, 447, 143–144. [Google Scholar] [CrossRef]
- Bandara, T.; Franks, A.; Xu, J.; Bolan, N.; Wang, H.; Tang, C. Chemical and biological immobilization mechanisms of potentially toxic elements in biochar-amended soils. Crit. Rev. Environ. Sci. Technol. 2020, 50, 903–978. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Alotaibi, M.A.; Alhammad, B.A.; Alharbi, B.M.; Refay, Y.; Badawy, S.A. Effects of ZnO nanoparticles and biochar of rice straw and cow Manure on characteristics of contaminated soil and sunflower productivity, oil quality, and heavy metals uptake. Agronomy 2020, 10, 790. [Google Scholar] [CrossRef]
- Li, H.; Dong, X.; Da, S.; Oliveira, L.; Chen, Y.; Ma, L. Mechanisms of metal sorption by biochars: Biochar characteristics and modifications. Chemosphere 2017, 178, 466–478. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.; Thies, J.; Masiello, C.; Hockaday, W.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Gorovtsov, A.V.; Minkina, T.M.; Mandzhieva, S.S.; Perelomov, L.V.; Soja, G.; Zamulina, I.V.; Rajput, V.D.; Sushkova, S.N.; Mohan, D.; Yao, J. The mechanisms of biochar interactions with microorganisms in soil. Environ. Geochem. Health 2020, 42, 2495–2518. [Google Scholar] [CrossRef]
- Jiang, X.; Tan, X.; Cheng, J.; Haddix, M.L.; Cotrufo, M.F. Interactions between aged biochar, fresh low molecular weight carbon and soil organic carbon after 3.5 years soil-biochar incubations. Geoderma 2019, 333, 99–107. [Google Scholar] [CrossRef]
- De la Rosa, J.; Rosado, M.; Paneque, M.; Miller, A.; Knicker, H. Effects of aging under field conditions on biochar structure and composition: Implications for biochar stability in soils. Sci. Total Environ. 2018, 613–614, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Pei, J.; Li, J.; Mia, S.; Singh, B.; Wu, J.; Dijkstra, F.A. Biochar aging increased microbial carbon use efficiency but decreased biomass turnover time. Geoderma 2020, 382, 114710. [Google Scholar] [CrossRef]
- Quan, G.; Fan, Q.; Zimmerman, A.R.; Sun, J.; Cui, L.; Wang, H.; Gao, B.; Yan, J. Effects of laboratory biotic aging on the characteristics of biochar and its water-soluble organic products. J. Hazard. Mater. 2020, 382, 121071. [Google Scholar] [CrossRef]
- Fan, S.; Zuo, J.; Dong, H. Changes in Soil Properties and Bacterial Community Composition with Biochar Amendment after Six Years. Agronomy 2020, 10, 746. [Google Scholar] [CrossRef]
- Li, K.; Yin, G.; Xu, Q.; Yan, J.; Hseu, Z.; Zhu, L.; Lin, Q. Influence of Aged Biochar Modified by Cd2+ on Soil Properties and Microbial Community. Sustainability 2020, 12, 4868. [Google Scholar] [CrossRef]
- Hu, J.; Wei, Z.; Kowalchuk, G.; Xu, Y.; Shen, Q.; Jousset, A. Rhizosphere microbiome functional diversity and pathogen invasion resistance build up during plant development. Environ. Microbiol. 2020, 22, 5005–5018. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.; Lee, S.; Zhang, M.; Tsang, Y.; Kim, K. Heavy metals in food crops: Health risks, fate, mechanisms, and management. Environ. Int. 2019, 125, 365–385. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Wang, R.; Gao, X.; Wang, K.; Lin, Z.; Ge, J.; Liu, T.; Wei, S.; Chen, W.; Xie, R.; et al. Cultivar-specific response of bacterial community to cadmium contamination in the rhizosphere of rice (Oryza sativa L.). Environ. Pollut. 2018, 241, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, C.; Wang, P.; Kretzschmar, R.; Zhao, F. Control of arsenic mobilization in paddy soils by manganese and iron oxides. Environ. Pollut. 2017, 231, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; Zhao, S.; Li, S.; Lei, X.; Qin, L.; Sun, X.; Chen, S. Manganese facilitates cadmium stabilization through physicochemical dynamics and amino acid accumulation in rice rhizosphere under flood-associated low pe+ pH. J. Hazard. Mater. 2021, 416, 126079. [Google Scholar] [CrossRef]
- Wang, B.; Lehmann, J.; Hanley, K.; Hestrin, R.; Enders, A. Adsorption and desorption of ammonium by maple wood biochar as a function of oxidation and pH. Chemosphere 2015, 138, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Guo, L.; Liao, P.; Xiong, Q.; Deng, X.; Pan, X.; Zeng, Y.; Zhang, H. Effects of biochar on the dynamic immobilization of Cd and Cu and rice accumulation in soils with different acidity levels. J. Clean. Prod. 2022, 372, 133730. [Google Scholar] [CrossRef]
- Chen, L.; Guo, L.; Liao, P.; Xiong, Q.; Deng, X.; Gao, H.; Wei, H.; Dai, Q.; Pan, X.; Zeng, Y.; et al. Rice Straw Biochar Reduces Cd Accumulation and promotes Cu accumulation in rice: Irrigation regime is the driving factor. J. Soils Sediments 2022. [Google Scholar] [CrossRef]
- Edwards, J.; Santos-Medellín, C.; Sundaresan, V. Extraction and 16S rRNA sequence analysis of microbiomes associated with rice Roots. Bio-Protocol 2018, 8, e2884. [Google Scholar] [CrossRef]
- Lindsay, W.; Norvell, W. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Xiao, Q.; Wang, Y.; Lü, Q.; Wen, H.; Han, B.; Chen, S.; Zheng, X.; Lin, R. Responses of glutathione and phytochelatins biosysthesis in a cadmium accumulator of Perilla frutescens (L.) Britt. under cadmium contaminated conditions. Ecotoxicol. Environ. Saf. 2020, 201, 110805. [Google Scholar] [CrossRef]
- Huang, D.; Xu, Y.; Lei, F.; Yu, X.; Ouyang, Z.; Chen, Y.; Jia, H.; Guo, X. Degradation of polyethylene plastic in soil and effects on microbial community composition. J. Hazard. Mater. 2021, 416, 126173. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson AJ, A.; Holmes, S.P. DADA2, High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Battaglia, M.L.; Shami, A.; Jalal, R.S.; Alhammad, B.A.; Almutairi, K.F.; Al-Saif, A.M. Biochar and its broad impacts in soil quality and fertility, nutrient leaching and crop productivity: A Review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Li, Z.; Liang, Y.; Hu, H.; Shaheen, S.; Zhong, H.; Tack, F.; Wu, M.; Li, Y.-F.; Gao, Y.; Rinklebe, J.; et al. Speciation, transportation, and pathways of cadmium in soil-rice systems: A review on the environmental implications and remediation approaches for food safety. Environ. Int. 2021, 156, 106749. [Google Scholar] [CrossRef]
- Futa, B.; Oleszczuk, P.; Andruszczak, S.; Kwiecińska-Poppe, E.; Kraska, P. Effect of Natural Aging of Biochar on Soil Enzymatic Activity and Physicochemical Properties in Long-Term Field Experiment. Agronomy 2020, 10, 449. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chen, D.; Shen, J.; Yuan, Q.; Fan, F.; Wei, W.-X.; Li, Y.; Wu, J. Biochar alters soil microbial communities and potential functions 3-4 years after amendment in a double rice cropping system. Agric. Ecosyst. Environ. 2021, 311, 107291. [Google Scholar] [CrossRef]
- Bian, R.; Joseph, S.; Cui, L.; Pan, G.; Li, L.; Liu, X.; Zhang, A.; Rutlidge, H.; Wong, S.; Chia, C.; et al. A three-year experiment confirms continuous immobilization of cadmium and lead in contaminated paddy field with biochar amendment. J. Hazard. Mater. 2014, 272, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Zhang, X.; Tang, C.; Muhammad, N.; Wu, J.; Brookes, P.C.; Xu, J. Potential role of biochars in decreasing soil acidification—A critical review. Sci. Total Environ. 2017, 581–582, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, M.; Wang, J.; Liu, X.; Guo, W.; Zheng, J.; Bian, R.; Wang, G.; Zhang, X.; Cheng, K.; et al. The responses of soil organic carbon mineralization and microbial communities to fresh and aged biochar soil amendments. Glob. Change Biol. Bioenergy 2019, 11, 1408–1420. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Yu, J.; Li, Z.; Wu, L.; Christie, P. Effects of biochar on the migration andtransformation of metal species in a highly acid soil contaminated with multiplemetals and leached with solutions of different pH. Chemosphere 2021, 278, 130344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Q.; Zeng, Y.; Zhang, J.; Lu, G.; Dang, Z.; Guo, C. Bioaccumulation and distribution of cadmium by Burkholderia cepacia GYP1 under oligotrophic condition and mechanism analysis at proteome level. Ecotoxicol. Environ. Saf. 2019, 176, 162–169. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, J.; Pan, G.; Liu, X.; Zhang, X.; Li, L.; Bian, R.; Cheng, K.; Jinwei, Z. Biochar decreased microbial metabolic quotient and shifted community composition four years after a single incorporation in a slightly acid rice paddy from southwest China. Sci. Total Environ. 2016, 571, 206–217. [Google Scholar] [CrossRef]
- Kirby, R. Actinomycetes and lignin degradation. Adv. Appl. Microbiol. 2009, 58, 125. [Google Scholar]
- Wang, R.; Wei, S.; Jia, P.; Liu, T.; Hou, D.; Xie, R.; Lin, Z.; Ge, J.; Qiao, Y.; Chang, X.; et al. Biochar significantly alters rhizobacterial communities and reduces Cd concentration in rice grains grown on Cd-contaminated soils. Sci. Total Environ. 2019, 676, 627–638. [Google Scholar] [CrossRef]
- Soothar, M.K.; Hamani, A.K.M.; Sardar, M.F.; Sootahar, M.K.; Fu, Y.; Rahim, R.; Soothar, J.K.; Bhatti, S.M.; Abubakar, S.A.; Gao, Y.; et al. Maize (Zea mays L.) Seedlings Rhizosphere Microbial Community as Responded to Acidic Biochar Amendment Under Saline Conditions. Front. Microbiol. 2021, 12, 789235. [Google Scholar] [CrossRef]
- Wang, C.; Huang, Y.; Yang, X.; Xue, W.; Zhang, X.; Zhang, Y.; Pang, J.; Liu, Y.; Liu, Z. Burkholderia sp. Y4 inhibits cadmium accumulation in rice by increasing essential nutrient uptake and preferentially absorbing cadmium. Chemosphere 2020, 252, 126603. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Tang, T. The effect of different straws on anaerobic removal of ficiency and mechanism of polycyclic aromatic hydroc-arbons in sludge by co-metabolism pathway. Master's Thesis, Guizhou University, Guiyang, China, 2019; p. P72. [Google Scholar]
- Xie, L.; Hao, P.; Cheng, Y.; Ahmed, I.; Cao, F. Effect of combined application of lead, cadmium, chromium and copper on grain, leaf and stem heavy metal contents at different growth stages in rice. Ecotoxicol. Environ. Saf. 2018, 168, 71–76. [Google Scholar] [CrossRef]
- Yue, L.; Lian, F.; Han, Y.; Bao, Q.L.; Wang, Z.Y.; Xing, B.S. The effect of biochar nanoparticles on rice plant growth and the uptake of heavy metals: Implications for agronomic benefits and potential risk. Sci. Total Environ. 2019, 656, 9–18. [Google Scholar] [CrossRef]
- Khan, S.; Chao, C.; Waqas, M.; Arp, H.P.H.; Zhu, Y.G. Sewage Sludge Biochar Influence upon Rice (Oryza sativa L.) Yield, Metal Bioaccumulation and Greenhouse Gas Emissions from Acidic Paddy Soil. Environ. Sci Technol 2013, 47, 8624–8632. [Google Scholar] [CrossRef]
- Lv, G.; Yang, T.; Chen, Y.; Hou, H.; Liu, X.; Li, J.; Wei, L.; Li, J. Biochar-based fertilizer enhanced Cd immobilization and soil quality in soil-rice system. Ecol. Eng. 2021, 171, 106396. [Google Scholar] [CrossRef]
- Yin, D.; Wang, X.; Peng, B.; Tan, C.; Ma, L.Q. Effect of biochar and Fe-biochar on Cd and As mobility and transfer in soil-rice system. Cheosphere 2017, 186, 928–937. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, H.; Tie, B.; Li, D.; Liu, S.; Lei, M.; Du, H. The long-term effectiveness of ferromanganese biochar in soil Cd stabilization and reduction of Cd bioaccumulation in rice. Biochar 2021, 3, 499–509. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Stage | Treatment | Cd Content (mg kg−1) | ||||
|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Rice Husk | Milled Rice | |||
| Huanghuazhan | Panicle initiation stage | CK | 2.304bcd | 0.207b | 0.432a | - | - |
| BC | 2.85b | 0.070d | 0.136d | - | - | ||
| Heading stage | CK | 2.496bc | 0.161bc | 0.43a | - | - | |
| BC | 1.589ef | 0.089d | 0.254c | - | - | ||
| Maturity stage | CK | 1.458f | 0.173b | 0.394ab | 0.048ab | 0.047b | |
| BC | 1.697def | 0.102d | 0.156d | 0.025c | 0.032c | ||
| Meixiangzhan2 | Panicle initiation stage | CK | 4.475a | 0.203b | 0.407a | - | - |
| BC | 2.239bcde | 0.077d | 0.121d | - | - | ||
| Heading stage | CK | 2.023cdef | 0.178b | 0.423a | - | - | |
| BC | 2.361bcd | 0.109cd | 0.14d | - | - | ||
| Maturity stage | CK | 2.159bc | 0.269a | 0.333b | 0.055a | 0.064a | |
| BC | 1.351f | 0.095d | 0.165d | 0.031bc | 0.042bc | ||
| Varieties | Stage | Treatment | pH | Total N Concentration (g.kg−1) | Organic Matter (g.kg−1) | Available P (mg.kg−1) |
|---|---|---|---|---|---|---|
| Huanghuazhan | Panicle initiation stage | CK | 5.74g | 1.72cd | 29.43d | 10.97c |
| BC | 6.61bc | 2.02a | 64.351a | 31.97a | ||
| Heading stage | CK | 5.81fg | 1.66cd | 28.91d | 10.28c | |
| BC | 6.73bc | 2.06a | 65.58a | 24.69b | ||
| Maturity stage | CK | 5.89ef | 1.63cd | 28.29d | 8.8c | |
| BC | 6.84ab | 1.89b | 59.1b | 20.5b | ||
| Meixiangzhan2 | Panicle initiation stage | CK | 6de | 1.68cd | 28.86d | 12.34c |
| BC | 6.63bc | 2.1a | 63.56ab | 29.14a | ||
| Heading stage | CK | 5.7g | 1.67cd | 28.38d | 9.43c | |
| BC | 6.58c | 2.05a | 65.31a | 23.05b | ||
| Maturity stage | CK | 6.02d | 1.61d | 28.06d | 9.34c | |
| BC | 6.87a | 1.74c | 49.35c | 21.79b |
| Varieties | Stage | Treatment | Total Cd Concentration (mg.kg−1) | Available Cd Concentration (mg.kg−1) |
|---|---|---|---|---|
| Huanghuazhan | Panicle initiation stage | CK | 0.615c | 0.365a |
| BC | 0.521d | 0.257b | ||
| Heading stage | CK | 0.859a | 0.362a | |
| BC | 0.677bc | 0.286b | ||
| Maturity stage | CK | 0.854a | 0.366a | |
| BC | 0.642c | 0.244b | ||
| Meixiangzhan2 | Panicle initiation stage | CK | 0.597c | 0.344a |
| BC | 0.496d | 0.267b | ||
| Heading stage | CK | 0.725b | 0.388a | |
| BC | 0.605c | 0.269b | ||
| Maturity stage | CK | 0.878a | 0.357a | |
| BC | 0.676bc | 0.248b |
| Varieties | Stage | Treatment | Richness Index | Diversity Index | ||
|---|---|---|---|---|---|---|
| ACE | Chao1 | Simpson | Shannon | |||
| Huanghuazhan | Panicle initiation stage | CK | 2302.599ab | 2336.273a | 0.997a | 9.7145a |
| BC | 2289.064abc | 2310.65ab | 0.9965a | 9.5742a | ||
| Heading stage | CK | 2252.6464abc | 2263.174ab | 0.9966a | 9.5353a | |
| BC | 2269.816abc | 2290.751ab | 0.997a | 9.6272a | ||
| Maturity stage | CK | 2121.454d | 2149.383c | 0.997a | 9.5964a | |
| BC | 2114.756d | 2140.713c | 0.9965a | 9.5967a | ||
| Meixiangzhan2 | Panicle initiation stage | CK | 2271.278abc | 2293.013ab | 0.9975a | 9.7879a |
| BC | 2200.051bcd | 2214.644bc | 0.997a | 9.6706a | ||
| Heading stage | CK | 2314.535a | 2334.605a | 0.9975a | 9.8159a | |
| BC | 2266.177abc | 2283.945ab | 0.9965a | 9.6309a | ||
| Maturity stage | CK | 2182.848cd | 2204.73bc | 0.9969a | 9.6999a | |
| BC | 2266.117abc | 2292.072ab | 0.9967a | 9.7393a | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X.; Long, C.; Chen, L.; Du, Y.; Zhang, Z.; Gan, L.; Zeng, Y. Special Microbial Communities Enhanced the Role of Aged Biochar in Reducing Cd Accumulation in Rice. Agronomy 2023, 13, 81. https://doi.org/10.3390/agronomy13010081
Deng X, Long C, Chen L, Du Y, Zhang Z, Gan L, Zeng Y. Special Microbial Communities Enhanced the Role of Aged Biochar in Reducing Cd Accumulation in Rice. Agronomy. 2023; 13(1):81. https://doi.org/10.3390/agronomy13010081
Chicago/Turabian StyleDeng, Xueyun, Changzhi Long, Le Chen, Yanli Du, Zhengwei Zhang, Li Gan, and Yongjun Zeng. 2023. "Special Microbial Communities Enhanced the Role of Aged Biochar in Reducing Cd Accumulation in Rice" Agronomy 13, no. 1: 81. https://doi.org/10.3390/agronomy13010081