
Biostimulants Using Humic Substances and Plant-Growth-Promoting Bacteria: Effects on Cassava (Manihot esculentus) and Okra (Abelmoschus esculentus) Yield
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant-Growth-Promoting Bacteria PGPB
2.2. Humic Acids
- -
- The hydrophobic index is the ratio of signal intensities found in chemical shift intervals for apolar alkyl and aromatic C components over those of hydrophilic C molecules:
- (1)
- HB/HI = Σ[(0–45) + (45–60)/2 + (110–160)]/Σ[(45–60)/2 + (60–110) + (160–190)];
- -
- The alkyl ratio determines the relative contribution of apolar versus polar components of aliphatic molecules:
- (2)
- A/OA = (0–45)/(60–110)
- -
- The aromaticity index compares the area assigned to aromatic compounds to that of alkyl fractions:
- (3)
- ARM = Σ[(45–60)/2 + (110–145) + (145–160)]/Σ[ (0–45) + (45–60)/2 + (60–110)]
- -
- The lignin ratio, relates the area of methoxyl-C+N-alkyl groups to that of O-aryl-C:
- (4)
- LigR = (45–60)/(145–160)
2.3. Inoculant Preparation
2.4. Pot Assay Experiment
2.5. Field Experiment
3. Results
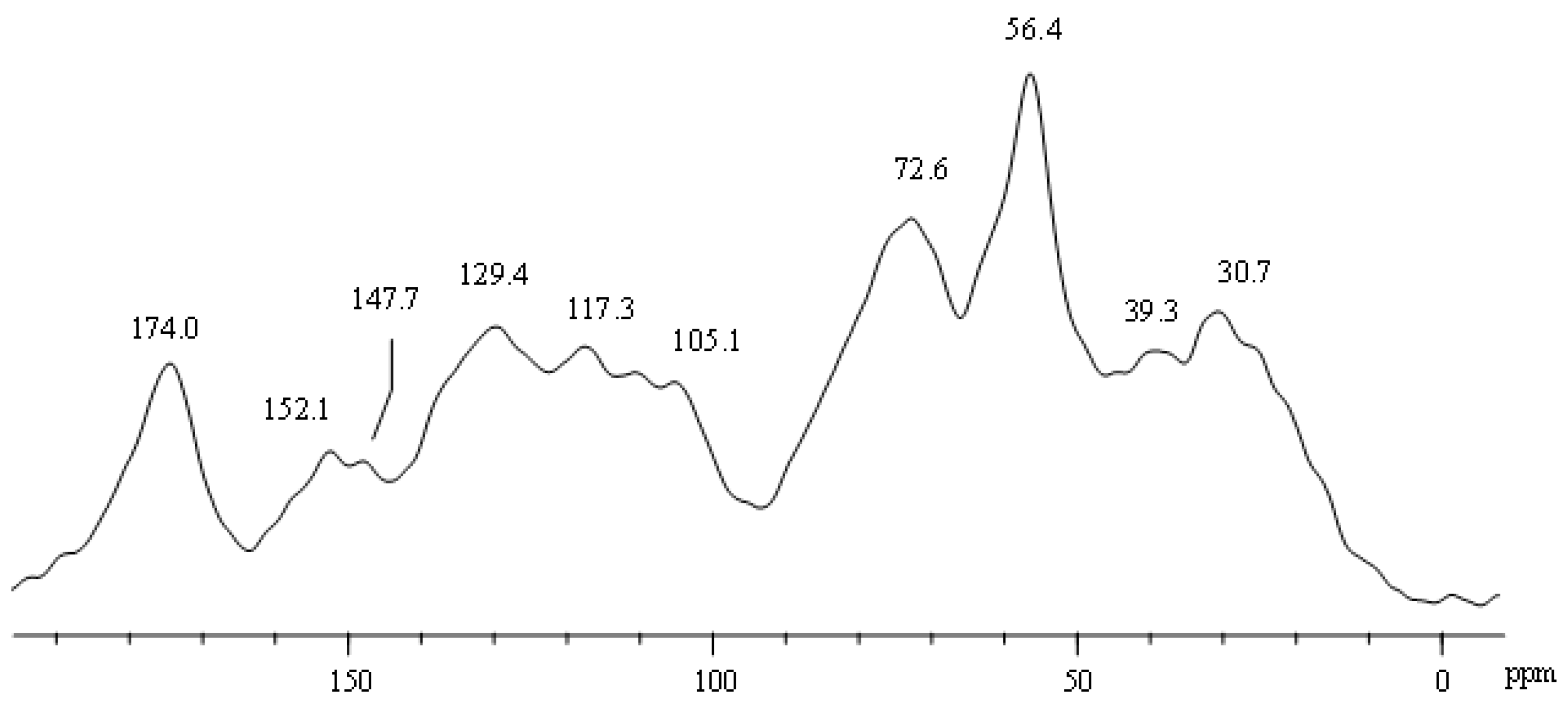
3.1. Chemical Characteristics of Humic Acids Used as a Vehicle for PGPB
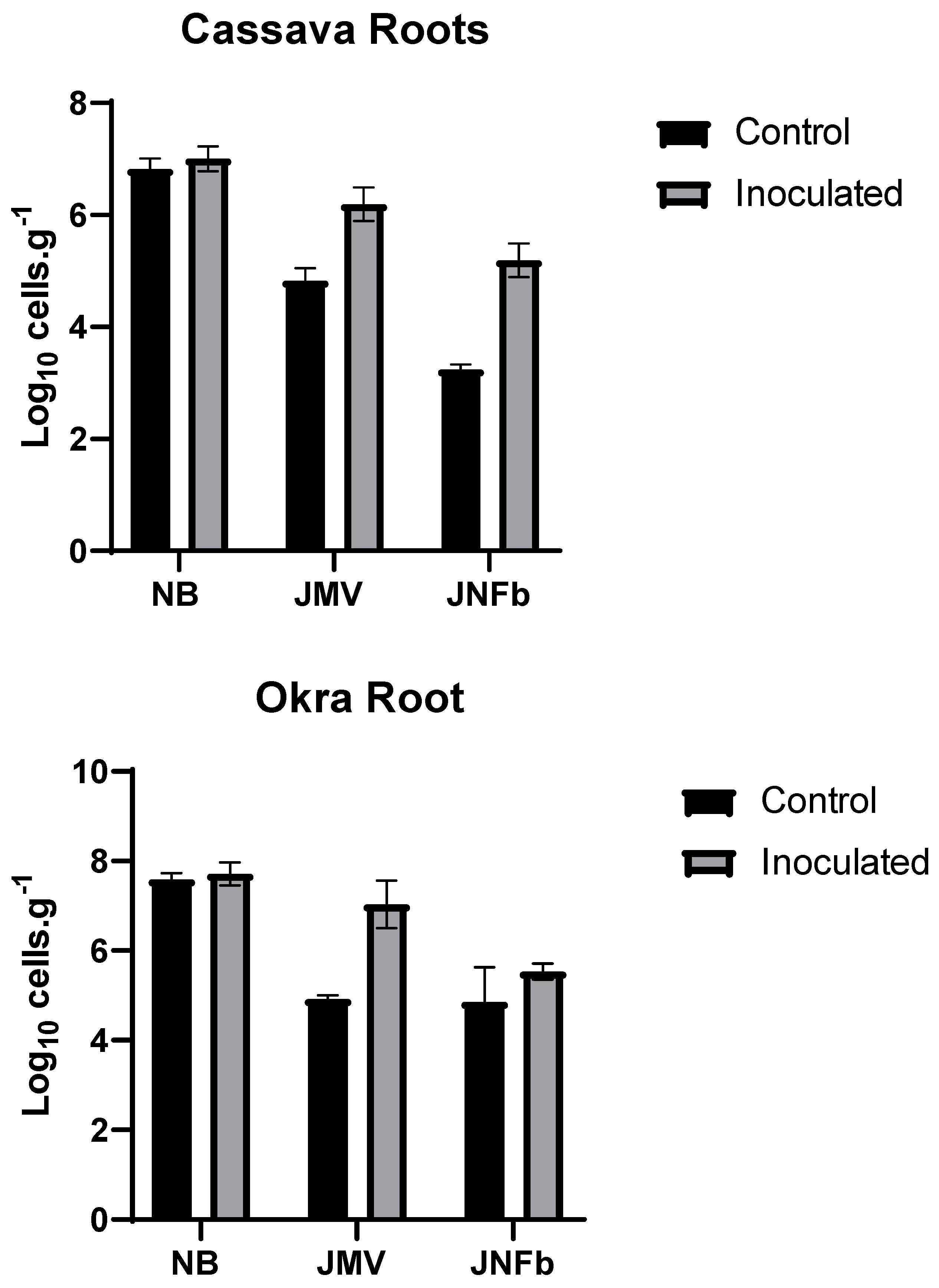
3.2. Initial Plant Growth and Diazotrophic Bacteria Population (Pot Assay)
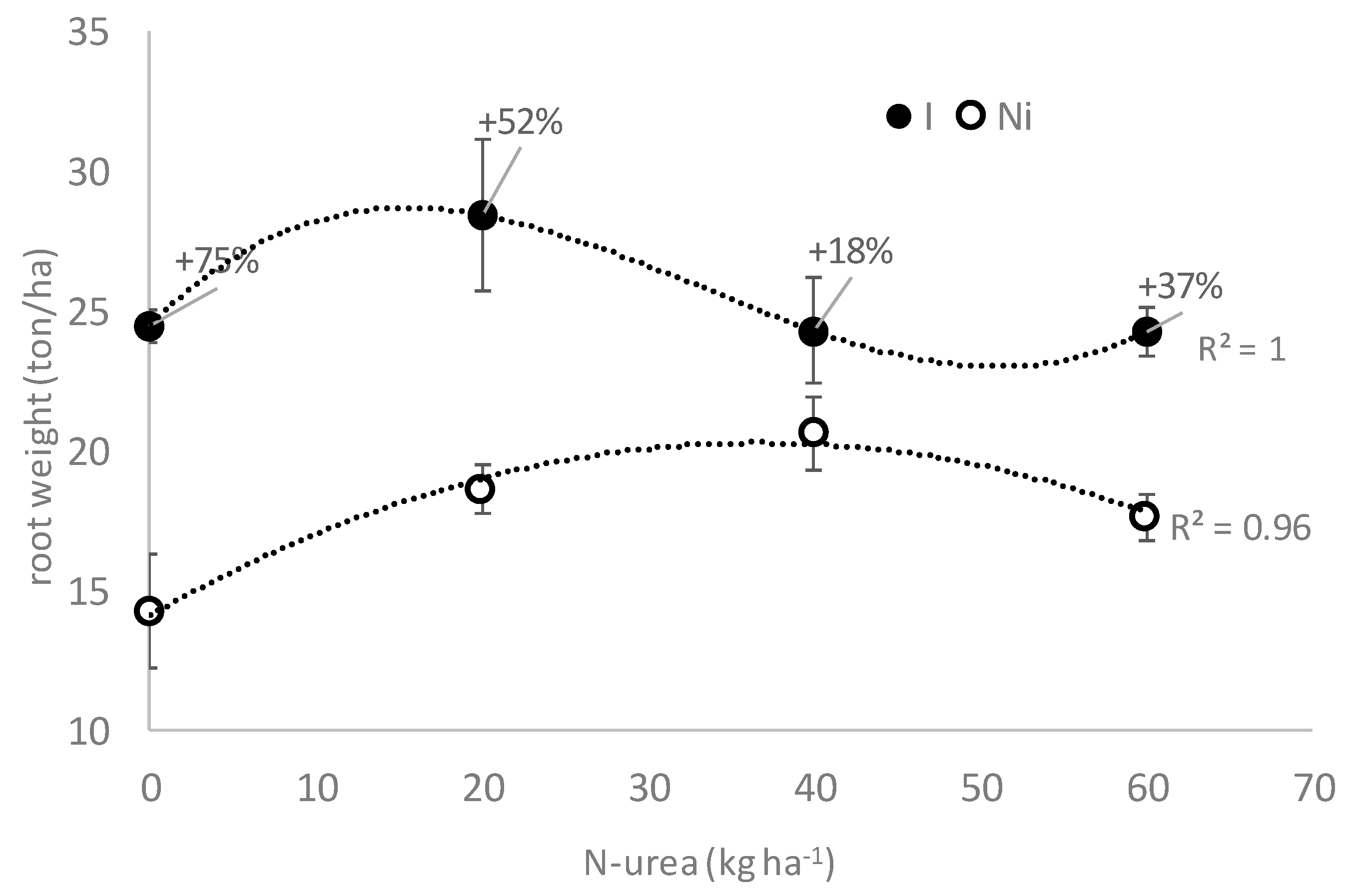
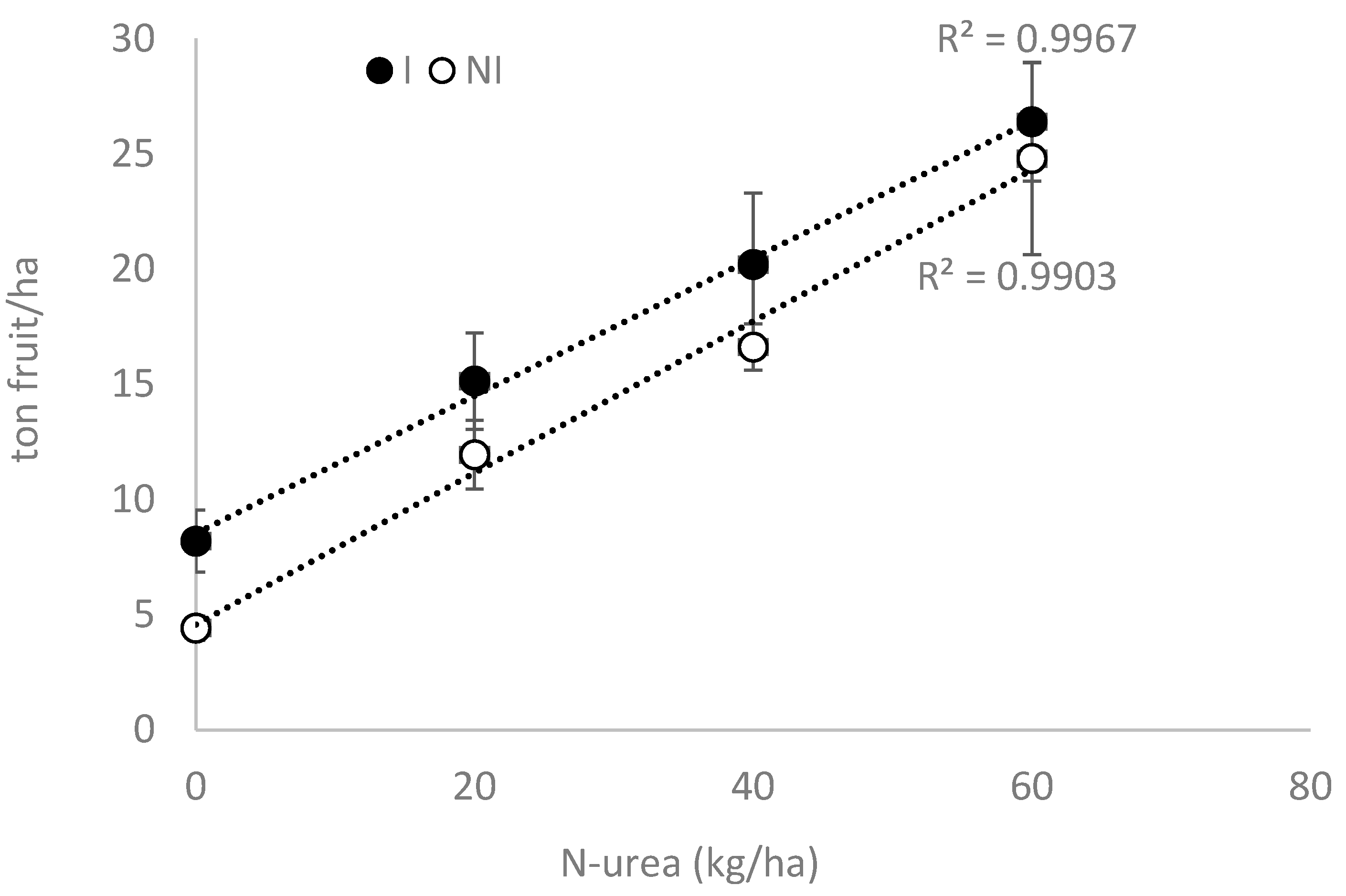
3.3. Field Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rede Brasileira em Pesquisa em Soberania e Segurança Alimentar e Nutricional (PEESAN): Inquérito Nacional Sobre Insegurança Alimentar no contexto da pandemia da COVID-19 no Brasil. Available online: https://olheparaafome.com.br/ (accessed on 15 October 2022).
- da Silva, J.G.; Sergio Gómez, E.; Rodrigo Castañeda, S. Agricultural boom and persistence of rural poverty in Latin America. Some reflections. Rev. Austral De Cienc. Soc. 2010, 18, 5–20. [Google Scholar]
- Albuquerque, A.C.S.; Silva, A.G. (Eds.) Agricultura Tropical: Quatro Décadas de Inovações Tecnológicas, Institucionais e Políticas—Portal Embrapa. 2008. Available online: https://www.embrapa.br/acre/busca-de-publicacoes/-/publicacao/507674/agricultura-tropical-quatro-decadas-de-inovacoes-tecnologicas-institucionais-e-politicas. (accessed on 17 June 2021).
- Patel, R. The Long Green Revolution. J. Peas Stud. 2013, 40, 1–63. [Google Scholar] [CrossRef]
- Mapbiomas. Available online: https://mapbiomas-br-site.s3.amazonaws.com/Fact-Sheet-Colecao7.pdf (accessed on 20 September 2022).
- Instituto Nacional de Pesquisas Espaciais—INPE/DETER (2022). Sistema de Detecção do Desmatamento na Amazônia Legal em Tempo Real (DETER)—Conjuntos de dados—Portal Brasileiro de Dados Abertos. Available online: https://dados.gov.br/dataset/deter (accessed on 1 October 2022).
- Altieri, M.A.; Toledo, V.M. The Agroecological Revolution in Latin America: Rescuing Nature, Ensuring Food Sovereignty and Empowering Peasants. J. Peas Stud. 2011, 38, 587–612. [Google Scholar] [CrossRef]
- van der Ploeg, J.A. O Sistema Alimentar em Tempos de COVID-19: Ensinamentos Para o Futuro CADERNOS PARA DEBATE n. 3. Available online: https://outraspalavras.net/wp-content/uploads/2021/10/211001-VanderPloegCriseAgricola.pdf (accessed on 7 September 2022).
- Cascudo, L.C. História da alimentação no Brasil: Editora Itatitaia; Global Editora: Belo Horizonte, Brazil, 2011; 900p. [Google Scholar]
- Aluko, M.; Kehinde-Fadare, A.B.; Oluwole, O.S. Growth and yield of okra (Abelmoschus esculentus (L.) Moench) under the influence of different organic fertilizers. IJRIAS 2020, 5, 162–166. [Google Scholar]
- Aboyeji, C.M.; Dahunsi, S.O.; Olaniyan, D.O.; Dunsin, O.; Adekiya, A.O.; Olayanju, A. Performance and quality attributes of okra (Abelmoschus esculentus (L.) Moench) fruits grown under soil-applied Zn-fertiliser, green biomass and poultry manure. Sci. Rep. 2021, 11, 8291. [Google Scholar] [CrossRef]
- Sabitha, V.; Ramachandran, S.; Naveen, K.R.; Panneerselvam, K. Antidiabetic and antihyperlipidemic potential of Abelmoschus esculentus (L.) Moench. in streptozotocin-induced diabetic rats. J. Pharm. Bioallied. Sci. 2011, 3, 397–402. [Google Scholar]
- Kamalesh, P.; Subrata, D.; Asraf, A.K.; Pranabesh, C. Phytochemical investigation and hypoglycaemic effect of Abelmoschus esculentus. Res. J. Pharm. Technol. 2016, 9, 162–164. [Google Scholar]
- da Visses, F.; Sentelhas, P.C.; Pereira, A.B. Yield gap of cassava crop as a measure of food security—An example for the main Brazilian producing regions. Food Sec. 2018, 10, 1191–1202. [Google Scholar] [CrossRef]
- de Oliveira Aparecido, L.E.; da Silva Cabral de Moraes, J.R.; de Meneses, K.C.; Lorençone, P.A.; Lorençone, J.A.; de Olanda Souza, G.H.; Torsoni, G.B. Agricultural zoning as tool for expansion of cassava in climate change scenarios. Theor. Appl. Climatol. 2020, 142, 1085–1095. [Google Scholar] [CrossRef]
- Silva, E.H.C.; Franco, C.A.; dos Santos Candido, W.; Braz, L.T. Morphoagronomic characterisation and genetic diversity of a Brazilian okra (Abelmoschus esculentus (L.) Moench) panel. Genet. Resour. Crop Evol. 2021, 68, 371–380. [Google Scholar] [CrossRef]
- Mohammed, L.; Mohammed, W.; Shiferaw, E. Phenotypic diversity assessment of okra (Abelmoschus Esculentus (L.) Moench) genotypes in Ethiopia using multivariate analysis. Scientifica 2022, 2022, 3306793. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Martínez-Balmori, D.; Médici, L.O.; Aguiar, N.O.; Campostrini, E.; Rosa, R.C.; Facanha, A.; Olivares, F.L. A combinateion of humic substances and Herbaspirillum seropedicae inoculation enhances the growth of maise (Zea mays L.). Plant Soil 2013, 366, 119–132. [Google Scholar] [CrossRef]
- Olivares, F.L.; Busato, J.G.; Paula, A.M.; Lima, L.S.; Aguiar, N.O.; Canellas, L.P. Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol. Technol. Agric. 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 3. [Google Scholar] [CrossRef]
- Olaetxea, M.; de Hita, D.; Calderin Garcia, A.; Fuentesa, M.; Baigorri, R.; Mora, V.; Garnica, M.; Urrutia, O.; Erro, J.; Zamarreño, A.M.; et al. Hypothetical framework integrating the main mechanisms involved in the promoting action of rhizospheric humic substances on plant root- and shoot growth. Appl. Soil Ecol. 2018, 123, 521–537. [Google Scholar] [CrossRef]
- Nardi, S.; Schiavon, M.; Francioso, O. Chemical structure and biological activity of humic substances define their role as plant growth promoters. Molecules 2021, 26, 2256. [Google Scholar] [CrossRef]
- Olivares, F.L.; Baldani, V.L.D.; Reis, V.M.; Baldani, J.I.; Döbereiner, J. Occurrence of the endophytic diazotrophs Herbaspirillum spp. in roots, stems and leaves predominantly of Gramineae. Biol. Fertil. Soils 1996, 21, 197–200. [Google Scholar] [CrossRef]
- Martinez-Balmori, D.; Spaccini, R.; Aguiar, N.O.; Novotny, E.H.; Olivares, F.L.; Canellas, L.P. Molecular characteristics of humic acids isolated from vermicomposts and their relationship to bioactivity. J. Agric. Food Chem. 2014, 62, 11412–11419. [Google Scholar] [CrossRef]
- de Aquino, A.M.; Canellas, L.P.; da Silva, A.P.S.; Canellas, N.A.O.; da S. Lima, L.; Olivares, F.L.; Piccolo, A.; Spaccini, R. Evaluation of molecular properties of humic acids from vermicompost by 13 CCPMAS-NMR spectroscopy and thermochemolysis–GC–MS. J. Anal. App. Pyrol. 2019, 141, 104634. [Google Scholar] [CrossRef]
- Aguiar, N.O.; Novotny, E.H.; Oliveira, A.L.; Rumjanek, V.M.; Olivares, F.L.; Canellas, L.P. Prediction of humic acids bioactivity using spectroscopy and multivariate analysis. J. Geochem. Expl. 2013, 129, 95–102. [Google Scholar] [CrossRef]
- Martinez-Balmori, D.; Olivares, F.L.; Spaccini, R.; Aguiar, K.P.; Araújo, M.F.; Aguiar, N.O.; Guridi, F.; Canellas, L.P. Molecular characteristics of vermicompost and their relationship to preservation of inoculated nitrogen-fixing bacteria. J. Anal. Appl. Pyrol. 2013, 104, 540–550. [Google Scholar] [CrossRef]
- EMBRAPA. 2021. Available online: http://www.cnpmf.embrapa.br/Base_de_Dados/index_pdf/dados/brasil/mandioca/b1_mandioca.pdf (accessed on 20 September 2022).
- Tsay, J.S.; Fukai, S.; Wilson, G.L. The response of cassava (Manihot esculenta) to spatial arrangement and to soybean intercrop. Field Crops Res. 1987, 16, 19–31. [Google Scholar] [CrossRef]
- Borsoi, T.N.; Freitas, S.J.; Souza, P.M.; Brito, P.M.B.; da Silva, W.S.; Soares, L.P.S.; Medeiros, S.R.; da Silva, L.C. Technological analyze of cassava production in the city of Campos Dos Goytacazes—RJ. J. Exp. Agric. Int. 2019, 35, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Omotose, S.O.; Shittu, O.S. Effect of NPK Fertilizer rates and method of application on growth and yield of okra (Abelmoschus esculentus L. Moench) at Ado-Ekiti Southwestern. Int. J. Agric. Res. 2007, 2, 614–619. [Google Scholar]
- Howeler, R.R. Cassava agronomy research in asia: Has it benefitted cassava farmers? Cent. Int. De Agric. Trop. 2017, 345–382. [Google Scholar]
- Pimentel, D.; Pimentel, M. Comment: Adverse Environmental Consequences of the Green Revolution. Pop Dev. Rev 1990, 16, 329. [Google Scholar] [CrossRef]
- Cabral, L.; Pandey, P.; Xu, X. Epic narratives of the Green Revolution in Brazil, China, and India. Agric. Hum. Values 2022, 39, 249–267. [Google Scholar] [CrossRef]
- Chilom, G.; Bruns, A.S.; Rice, J.A. Aggregation of humic acid in solution: Contribution of different fractions. Org. Geochem. 2013, 40, 455–460. [Google Scholar] [CrossRef]
- Chernysheva, M.G.; Badun, G.; Kulikova, N.; Perminova, I.V. Behavior of humic substances in the liquid-liquid system directly measured using tritium label. Chemosphere 2020, 238, 124646. [Google Scholar] [CrossRef]
- Terashima, M.; Fukushima, M.; Tanaka, S. Influence of pH on the surface activity of humic acid: Micelle-like aggregate formation and interfacial adsorption. Colloids Surf. A Physicochem. Eng. Asp. 2004, 247, 77–83. [Google Scholar] [CrossRef]
- Tian, S.; Tan, W.; Wang, X.; Huang, N.; Bai, Y. Surface activity of humic acid and its sub-fractions from forest soil. Sustainability 2021, 13, 8122. [Google Scholar] [CrossRef]
- da Silva, S.F.; Olivares, F.L.; Canellas, L.P. The biostimulant manufactured using diazotrophic endophytic bacteria and humates is effective to increase sugarcane yield. Chem. Biol. Technol. Agric. 2017, 4, 24. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hort. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Khillari, S.; Biostimulant Market Analysis. Recent Industry Trends Report 2026. 2020. Available online: https://www.fortunebusinessinsights.com/industry-reports/biostimulants-market-100414 (accessed on 20 September 2022).
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.; Cavagnaro, T.R. A meta-analysis and review of plant-growth response to humic substances: Practical implications for agriculture. Adv. Agron. 2014, 124, 37–89. [Google Scholar]
- Arancon, N.Q.; Owens, J.D.; Converse, C. The effects of vermicompost tea on the growth and yield of lettuce and tomato in a non-circulating hydroponics system. J. Plant Nutr. 2019, 42, 2447–2458. [Google Scholar] [CrossRef]
- Mal, B.P.; Mahapatra, P.; Mohanty, S. Effect of diazotrophs and chemical fertilizers on production and economics of okra (Abelmoschus esculentus, L.) Cultivars. Am. J. Plant Sci. 2014, 5, 168–174. [Google Scholar] [CrossRef]
- Balota, E.L.; Lopes, E.S.; Hungria, M.; Döberainer, J. Ocorrência de bactérias diazotróficas e fungos micorrízicos arbusculares na cultura da mandioca. Pesq. Agropec. Bras 1999, 34, 1265–1276. [Google Scholar] [CrossRef]
- MacLaren, C.; Mead, A.; van Balen, D.; Claessens, L.; Etana, A.; de Haan, J.; Haagsma, W.; Jäck, O.; Keller, T.; Labuschagne, J.; et al. Long-term evidence for ecological intensification as a pathway to sustainable agriculture. Nat. Sustain. 2022, 5, 770–779. [Google Scholar] [CrossRef]
- Duru, M.; Therond, O.; Fares, M. Designing agroecological transitions; A review. Agron. Sustain. Dev. 2015, 35, 1237–1257. [Google Scholar] [CrossRef] [Green Version]
- Song, X.Y.; Spaccini, R.; Pan, G.; Piccolo, A. Stabilization by hydrophobic protection as a molecular mechanism for organic carbon sequestration in maize-amended rice paddy soils. Sci. Total Environ. 2013, 458-460, 319–330. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.F.; Spaccini, R.; Mazzei, P.; de Rezende, C.E.; Canellas, L.P. Changes in water-extractable organic matter in tropical forest and agricultural soils as detected by the DRIFT spectroscopy technique. Land Degrad. Dev. 2021, 32, 4755–4767. [Google Scholar] [CrossRef]
- Habanyati, E.J.; Nyanga, P.H.; Umar, B.B. Factors contributing to disadoption of conservation agriculture among smallholder farmers in Petauke, Zambia. Kasetsart J. Soc. Sci. 2020, 41, 91–96. [Google Scholar] [CrossRef]
- Zhou, P.; Pan, G.X.; Spaccini, R.; Piccolo, A. Molecular changes in particulate organic matter (POM) in a typical Chinese paddy soil under different long-term fertilizer treatments. Eur. J. Soil Sci. 2010, 61, 231–242. [Google Scholar] [CrossRef]
- Canellas, L.P.; Spaccini, R.; Aguiar, N.O.; Garcia-Vega, M.R.; Olivares, F.L. Soil Organic Matter Quality From Soils Cropped by Traditional Peasants. Sustain. Agric. Res. 2014, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Miah, M.A.M.; Haque, M.E.; Bell, R.W.; Akhter, S.; Hossain, M.B. Availability and utilisation pattern of agricultural waste at household level in selected areas of Bangladesh. Waste Man Res. 2022, 40, 1277–1284. [Google Scholar] [CrossRef]
- Singh, S.; Sinha, R.K. Vermicomposting of organic wastes by earthworms: Making wealth from waste by converting ‘garbage into gold’ for farmers. In Advanced Organic Waste Management: Sustainable Practices and Approaches; Elsevier Radarweg 29: Amsterdam, Netherlands, 2022; pp. 93–120. [Google Scholar]
- Rosset, P.M.; Altieri, M.A. Agroecology: Science and Politics; Fernwood Publishing: Winnipeg, NS, Canada, 2017. [Google Scholar]
- Giller, K.E.; Hijbeek, R.; Andersson, J.A.; Sumberg, J. Regenerative Agriculture: An agronomic perspective. Outlook Agric. 2021, 50, 13–25. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 190–160 C=O | 160–145 O-aryl-C | 145–110 Aryl-C | 110–60 O-Alkyl-C | 60–45 CH3O/C-N | 45–0 Alkyl-C | HB | A/OA | Ar | LR | |
|---|---|---|---|---|---|---|---|---|---|---|
| HAs | 9.5 | 5.7 | 20.1 | 31.0 | 14.1 | 19.5 | 1.1 | 0.6 | 0.6 | 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canellas, L.P.; Canellas, N.O.A.; da Silva, R.M.; Spaccini, R.; Mota, G.P.; Olivares, F.L. Biostimulants Using Humic Substances and Plant-Growth-Promoting Bacteria: Effects on Cassava (Manihot esculentus) and Okra (Abelmoschus esculentus) Yield. Agronomy 2023, 13, 80. https://doi.org/10.3390/agronomy13010080
Canellas LP, Canellas NOA, da Silva RM, Spaccini R, Mota GP, Olivares FL. Biostimulants Using Humic Substances and Plant-Growth-Promoting Bacteria: Effects on Cassava (Manihot esculentus) and Okra (Abelmoschus esculentus) Yield. Agronomy. 2023; 13(1):80. https://doi.org/10.3390/agronomy13010080
Chicago/Turabian StyleCanellas, Luciano P., Natália O. A. Canellas, Rakiely M. da Silva, Riccardo Spaccini, Gabriela Petroceli Mota, and Fábio L. Olivares. 2023. "Biostimulants Using Humic Substances and Plant-Growth-Promoting Bacteria: Effects on Cassava (Manihot esculentus) and Okra (Abelmoschus esculentus) Yield" Agronomy 13, no. 1: 80. https://doi.org/10.3390/agronomy13010080