
Analysis of Photosynthetic Differences of Rice Germplasm in Southeast Asia Based on Leaf-Tissue Structure, Physiology, and iTRAQ
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Investigations and Measurements
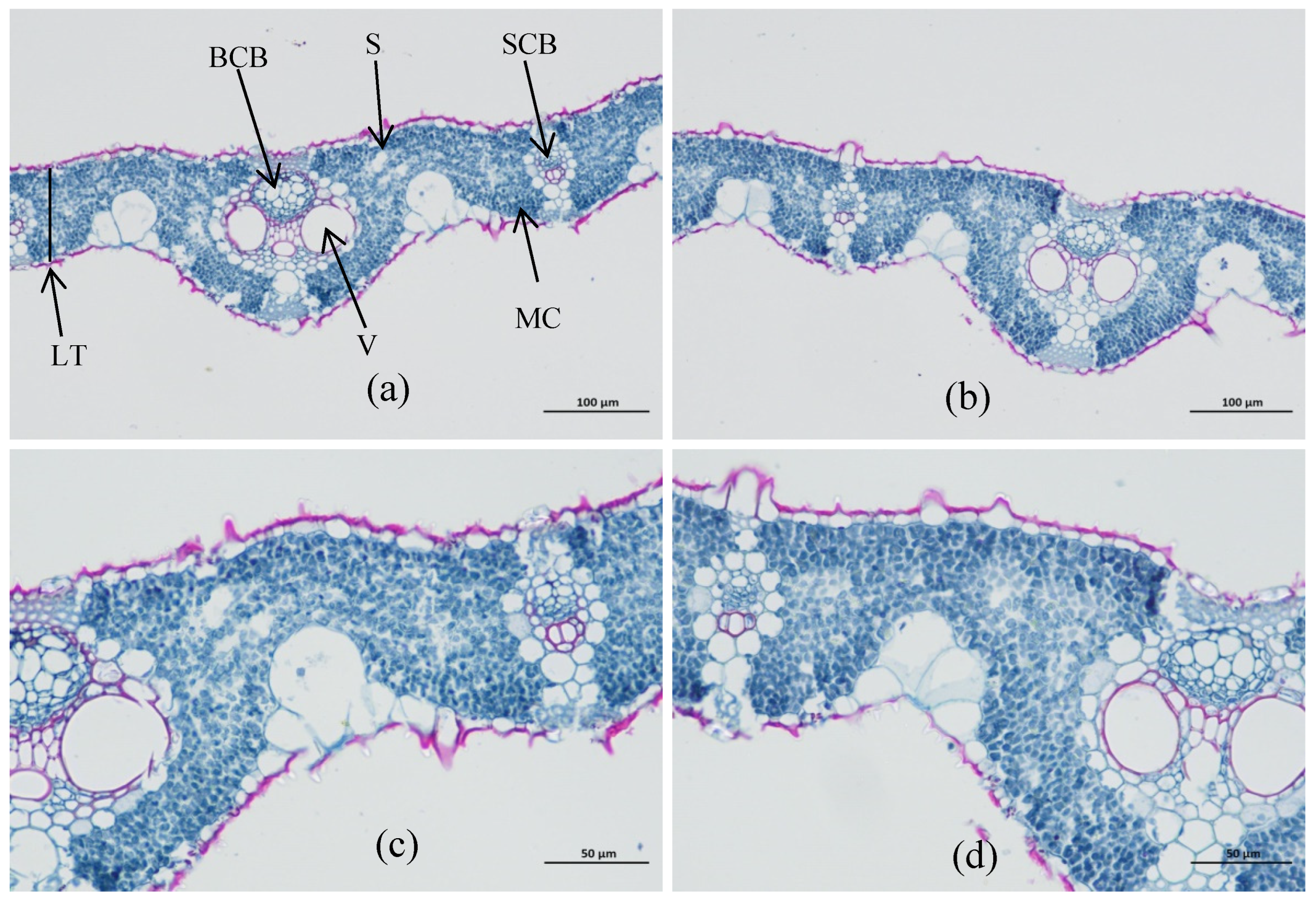
2.2.1. Leaf Tissue Structure
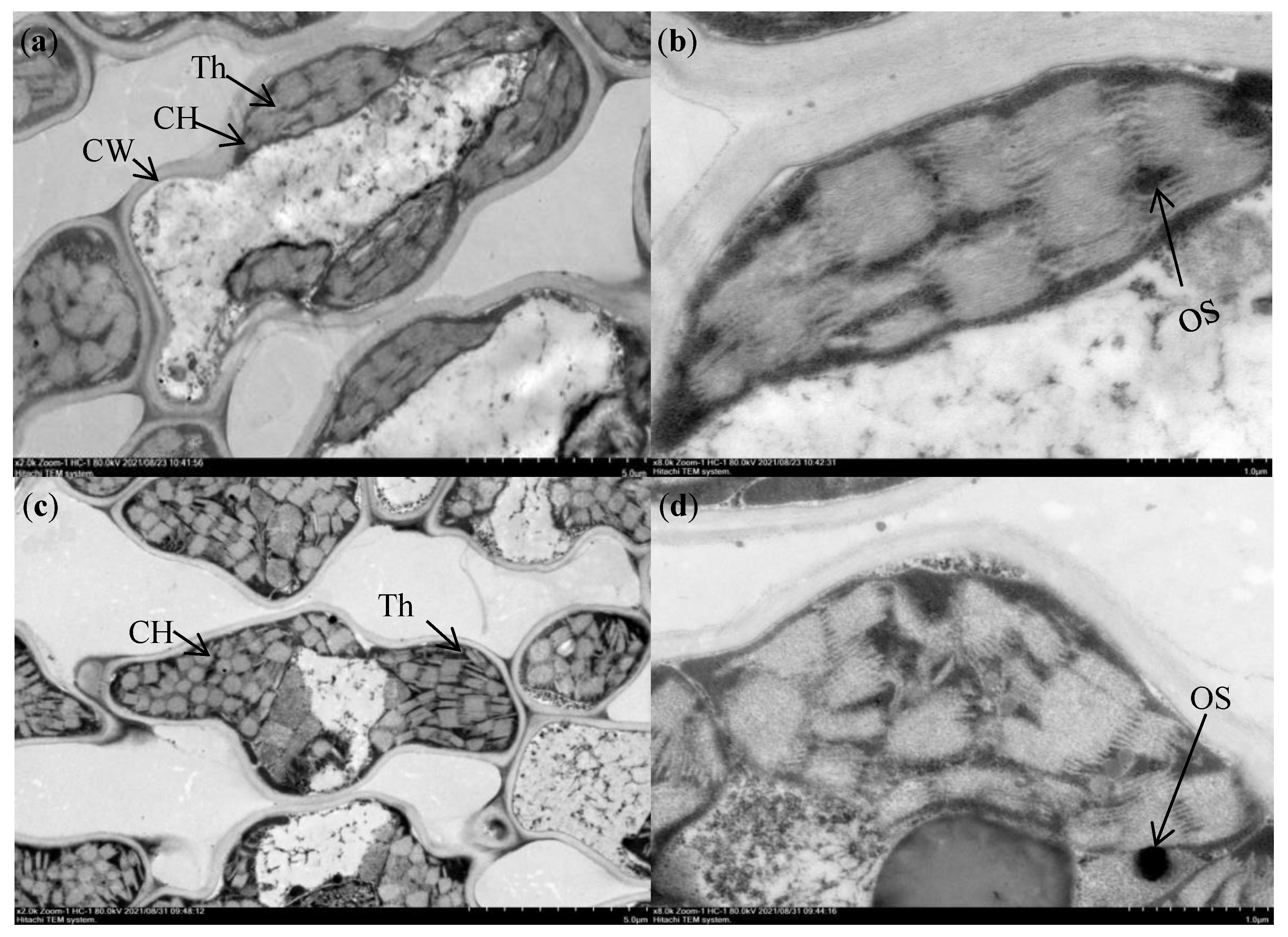
2.2.2. Chloroplast Ultrastructure
2.3. Physiological Indicators
2.3.1. Photosynthetic Characteristics
2.3.2. Chlorophyll Fluorescence Parameters
2.3.3. Chlorophyll Content SPAD and RSP
2.3.4. Rubisco Enzyme Activity and Content
2.4. iTRAQ Proteomics
2.4.1. Protein Extraction, Trypsin Digestion and iTRAQ Labeling
2.4.2. High-Performance Liquid Chromatography (HPLC) Fractionation
2.4.3. Liquid Chromatography (LC) Tandem Mass Spectrometry (MS/MS) Analysis
2.4.4. Database Search
2.5. Bioinformatics Analysis
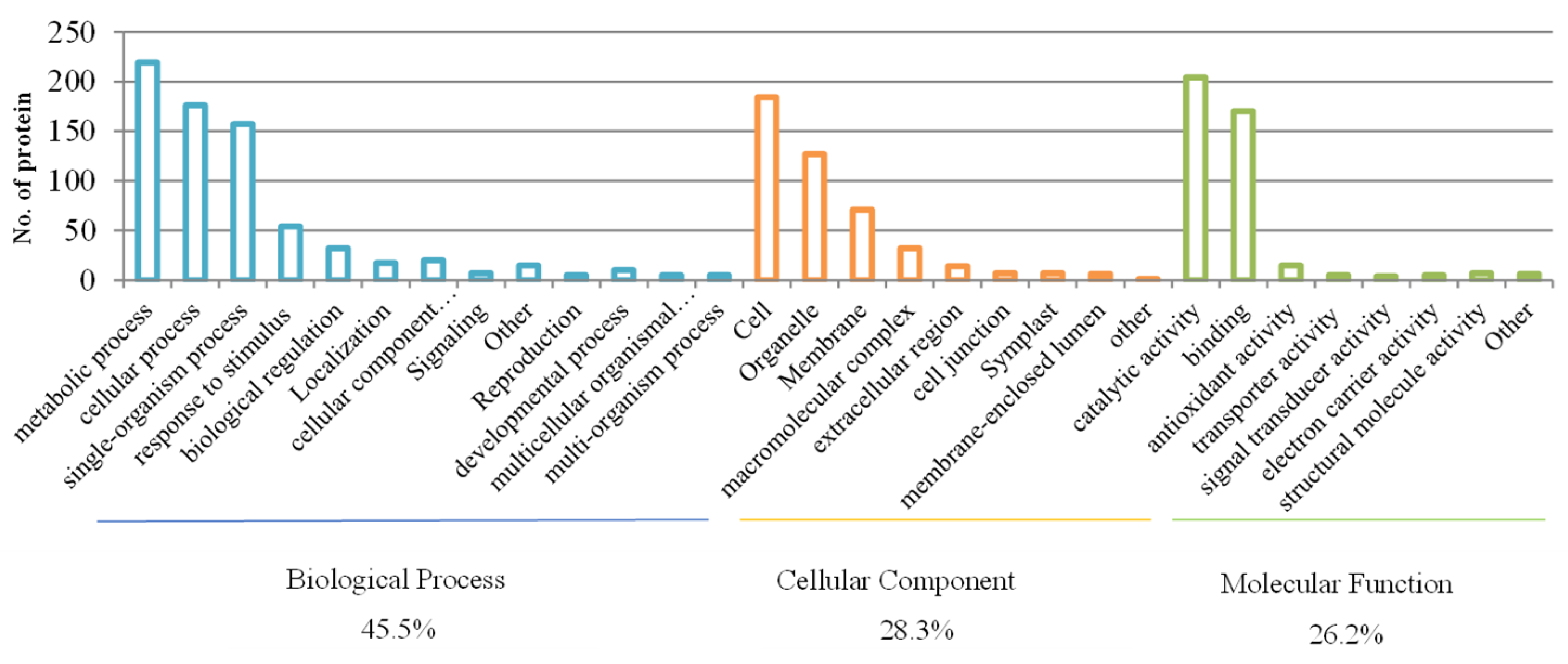
2.5.1. Gene Ontology (GO) Annotation
2.5.2. Domain Annotation
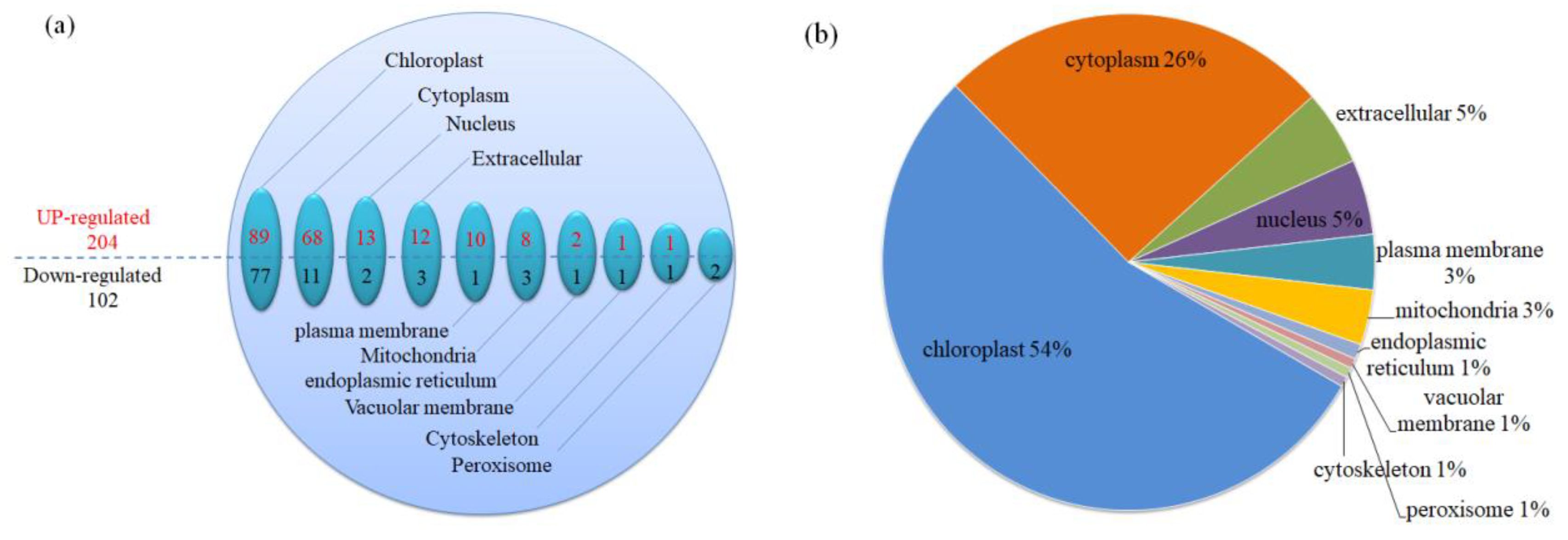
2.5.3. Sequence Subcellular Localization
2.5.4. Enrichment of GO Analysis
2.5.5. Enriched Pathway Analysis
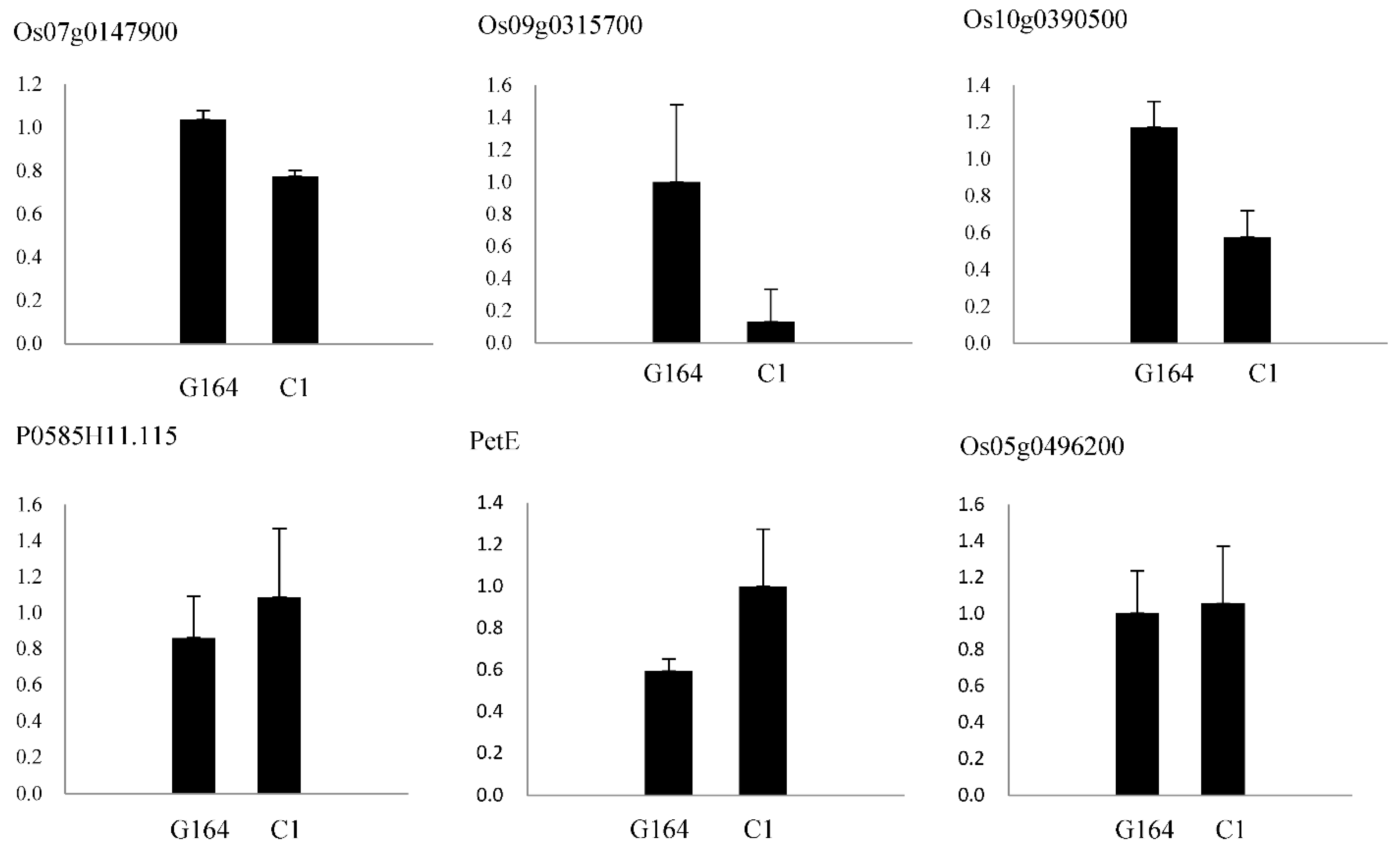
2.6. Real-Time Quantitative PCR Analysis
2.7. Statistics and Analysis
3. Results
3.1. Differences in Leaf Structure
3.2. Differences in Chloroplast Ultrastructure
3.3. Differences in Chlorophyll Value and Photosynthetic Rate
3.4. Differences in Chlorophyll Fluorescence Parameters and Rubisco
3.5. Proteomic Analysis of Different Germplasm Resources
3.5.1. Protein Identification and GO Analysis
3.5.2. Classification of Subcellular Structure Localization
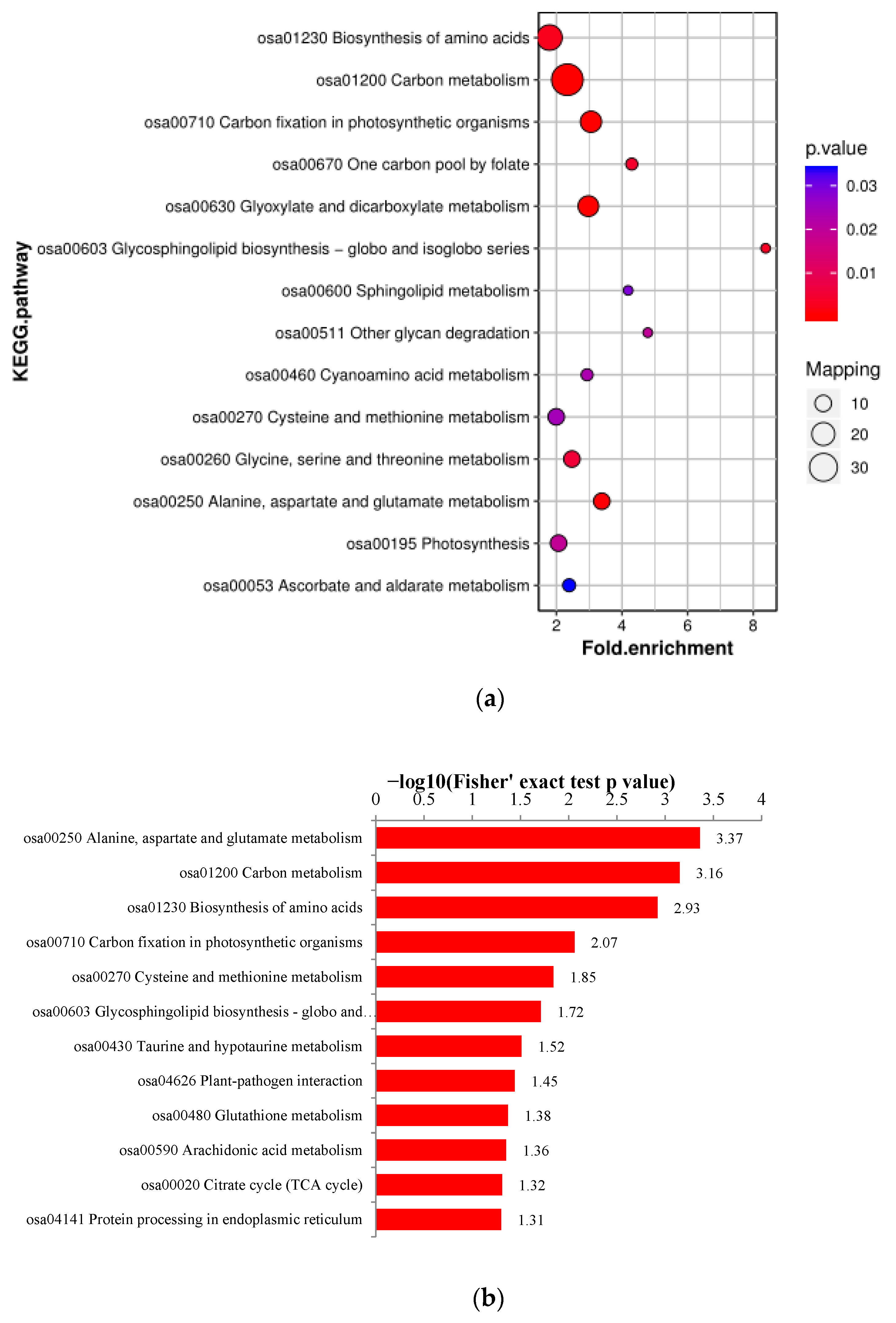
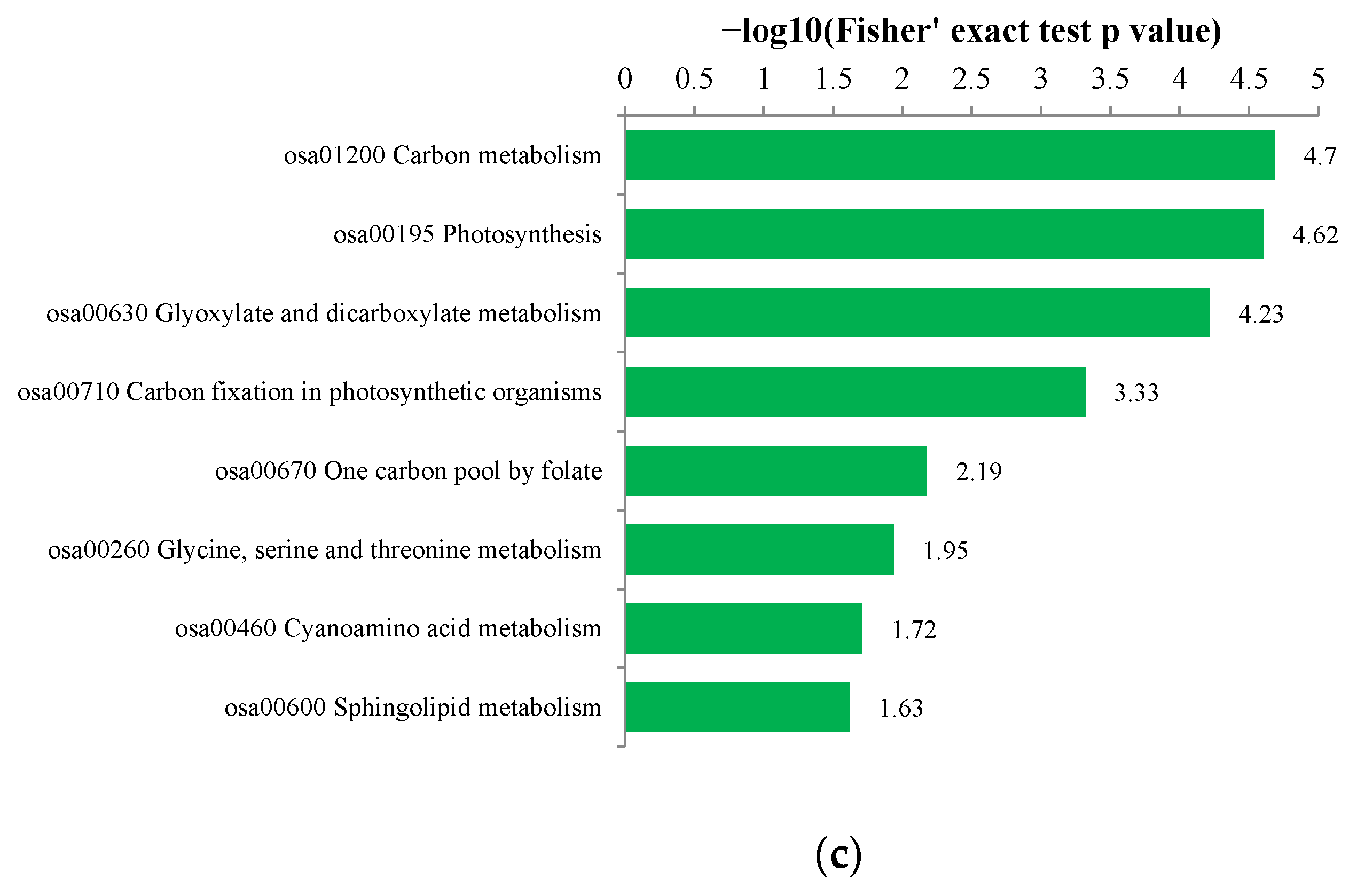
3.5.3. KEGG Analysis
4. Discussion
4.1. Effect of Tissue Structure on Pn
4.2. Effect of Photosynthetic Physiological Indicators on Pn
4.3. Differential Proteins among High- and Low-Light-Efficiency Germplasm Resources
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Murchie, E.H.; Pinto, M.; Horton, P. Agriculture and the new challenges for photosynthesis research. New Phytol. 2009, 181, 532–552. [Google Scholar] [CrossRef] [PubMed]
- Horton, P. Prospects for crop improvement through the genetic manipulation of photosynthesis: Morphological and biochemical aspects of light capture. J. Exp. Bot. 2000, 51, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Marshall-Colon, A.; Zhu, X.-G. Meeting the Global Food Demand of the Future by Engineering Crop Photosynthesis and Yield Potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, S.; Yamamoto, T.; Nakae, T.; Yamashita, M.; Uchida, M.; Karimata, R.; Ichihara, N.; Soda, K.; Ochiai, T.; Ao, R.; et al. Genetic architecture of leaf photosynthesis in rice revealed by different types of reciprocal mapping populations. J. Exp. Bot. 2019, 70, 5131–5144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Peng, S.; Li, Y. Intraspecific variation in photosynthetic nitrogen-use efficiency is positively related to photosynthetic rate in rice (Oryza sativa L.) plants. Photosynthetica 2019, 57, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Xiong, D.; Liu, X.; Liu, L.; Douthe, C.; Li, Y.; Peng, S.; Huang, J. Rapid responses of mesophyll conductance to changes of CO2 concentration, temperature and irradiance are affected by N supplements in rice. Plant Cell Environ. 2015, 38, 2541–2550. [Google Scholar] [CrossRef]
- Teng, S.; Qian, Q.; Zeng, D.; Kunihiro, Y.; Fujimoto, K.; Huang, D.; Zhu, L. QTL analysis of leaf photosynthetic rate and related physiological traits in rice (Oryza sativa L.). Euphytica 2004, 135, 1–7. [Google Scholar] [CrossRef]
- Degl’Innocenti, E.; Guidi, L.; Soldatini, G.F. Effects of elevated ozone on chlorophyll a fluorescence in symptomatic and asymptomatic leaves of two tomato genotypes. Biol. Plant. 2007, 51, 313–321. [Google Scholar] [CrossRef]
- Li, H.S.; Sun, Q.; Zhao, S.J. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; pp. 167–169. [Google Scholar]
- Griffiths, H.; Weller, G.; Toy, L.; Dennis, R. You’re so vein: Bundle sheath physiology, phylogeny and evolution in C3 and C4 plants. Plant Cell Environ. 2013, 36, 249–261. [Google Scholar] [CrossRef]
- Long, S.; Zhu, X.; Naidu, S.; Ort, D. Can improvement in photosynthesis increase crop yield? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Rosenthal, D.M.; Locke, A.M.; Khozaei, M.; Raines, C.A.; Long, S.P.; Ort, D.R. Over-expressing the C3 photosynthesis cycle enzyme Sedoheptulose-1-7 Bisphosphatase improves photosynthetic carbon gain and yield under fully open air CO2 fumigation (FACE). BMC Plant Biol. 2011, 11, 123. [Google Scholar] [CrossRef] [Green Version]
- Takahara, K.; Kasajima, I.; Takahashi, H.; Hashida, S.-N.; Itami, T.; Onodera, H.; Toki, S.; Yanagisawa, S.; Kawai-Yamada, M.; Uchimiya, H. Metabolome and Photochemical Analysis of Rice Plants Overexpressing Arabidopsis NAD Kinase Gene. Plant Physiol. 2010, 152, 1863–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tholen, D.; Boom, C.; Zhu, X. Opinion: Prospects for improving photosynthesis by altering leaf anatomy. Plant Sci. 2012, 197, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Coupe, S.; Palmer, B.; Lake, J.; Overy, S.; Oxborough, K.; Woodward, F.; Gray, J.; Quick, W. Systemic signalling of environmental cues in Arabidopsis leaves. J. Exp. Bot. 2006, 57, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, R.; Koteyeva, N.; Voznesenskaya, E.; Evans, M.A.; Cousins, A.B.; Edwards, G.E. Coordination of Leaf Photosynthesis, Transpiration, and Structural Traits in Rice and Wild Relatives (Genus Oryza). Plant Physiol. 2013, 162, 1632–1651. [Google Scholar] [CrossRef] [Green Version]
- Shunsuke, A.; Toru, N.; Masaki, U.; Kazuya, S.; Toshiyuki, T.; Takao, O.; Toshio, Y.; Taiichiro, O.; Hiroshi, M.; Masahiro, Y. The mesophyll anatomy enhancing CO2 diffusion is a key trait for improving rice photosynthesis. J. Exp. Bot. 2013, 64, 1061–1072. [Google Scholar]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [Green Version]
- Ocheltree, T.W.; Nippert, J.; Prasad, P.V. Changes in stomatal conductance along grass blades reflect changes in leaf structure. Plant Cell Environ. 2012, 35, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Shu, Y.; Peng, S.; Li, Y. Leaf photosynthesis is positively correlated with xylem and phloem areas in leaf veins in rice (Oryza sativa) plants. Ann. Bot. 2022, 129, 619–631. [Google Scholar] [CrossRef]
- Xiong, D.; Yu, T.; Zhang, T.; Li, Y.; Peng, S.; Huang, J. Leaf hydraulic conductance is coordinated with leaf morpho-anatomical traits and nitrogen status in the genus Oryza. J. Exp. Bot. 2015, 66, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Zhang, Y.; Lei, Z.; Zhang, W. The higher area-based photosynthesis in Gossypium hirsutum L. is mostly attributed to higher leaf thickness. Photosynthetica 2019, 57, 420–427. [Google Scholar] [CrossRef]
- Deng, Y.M.; Li, C.; Shao, Q.; Ye, X.; She, J. Differential responses of double petal and multi petal jasmine to shading: I. Photosynthetic characteristics and chloroplast ultrastructure. Plant Physiol. Biochem. 2012, 55, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Huang, J.; Peng, S.; Xiong, D. Diffusional conductance to CO2 is the key limitation to photosynthesis in salt-stressed leaves of rice (Oryza sativa). Physiol. Plant. 2017, 163, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Wu, L.; Chen, J.; Dong, L. Morphological plasticity, photosynthesis and chlorophyll fluorescence of Athyrium pachyphlebium at different shade levels. Photosynthetica 2011, 49, 611–618. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Lande, N.V.; Barua, P.; Gayen, D.; Kumar, S.; Chakraborty, S.; Chakraborty, N. Proteomic dissection of the chloroplast: Moving beyond photosynthesis. J. Proteom. 2019, 212, 103542. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, C.; Li, K.; Cai, X.; Wu, M.; Chen, G. Fe deficiency induced changes in rice (Oryza sativa L.) thylakoids. Environ. Sci. Pollut. Res. 2017, 24, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Wang, Y. Proteomic and Physiological Studies Provide Insight into Photosynthetic Response of Rice (Oryza sativa L.) Seedlings to Microgravity. Photochem. Photobiol. 2016, 92, 561–570. [Google Scholar] [CrossRef]
- Sakataa, S.; Mizusawaa, N.; Kawaib, H.; Sakuraia, I.; Wada, H. Psb28 is involved in recovery of photosystem II at high temperature in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta (BBA)-Bioenerg. 2013, 1827, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Zörb, C.; Herbst, R.; Forreiter, C.; Schubert, S. Short-term effects of salt exposure on the maize chloroplast protein pattern. Proteomics 2009, 9, 4209–4220. [Google Scholar] [CrossRef]
- Fan, Y.; Chen, J.; Wang, Z.; Tan, T.; Li, S.; Li, J.; Wang, B.; Zhang, J.; Cheng, Y.; Wu, X.; et al. Soybean (Glycine max L. Merr.) seedlings response to shading: Leaf structure, photosynthesis and proteomic analysis. BMC Plant Biol. 2019, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Li, M.; Qiu, Z.; Chen, D.; Zhang, G.; Wang, X.; Chen, G.; Hu, J.; Gao, Z.; Dong, G.; et al. Primary leaf-type ferredoxin 1 participates in photosynthetic electron transport and carbon assimilation in rice. Plant J. 2020, 104, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Feng, D. Photosynthetic Characteristics and Quantitative Proteomics Analysis of Maize and Rice Leaves. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2007. [Google Scholar]
- Zhang, Z.; Mao, X.; Ou, J.; Ye, N.; Zhang, J.; Peng, X. Distinct photo respiratory reactions are preferentially catalyzed by glutamate: Glyoxylate and serine: Glyoxylate aminotransferases in rice. J. Photochem. Photobiol. B Biol. 2015, 142, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, N.; Samejima, M.; Ishihara, K. Characteristics of photosynthetic carbon metabolism of spikelets in rice. Photosynth. Res. 1997, 52, 75–82. [Google Scholar] [CrossRef]
- Agarie, S.; Miura, A.; Sumikura, R.; Tsukamoto, S.; Nose, A.; Arima, S.; Matsuoka, M.; Miyao-Tokutomi, M. Overexpression of C4 PEPC caused O2-insensitive photosynthesis intransgenic rice plant. Plant Sci. 2002, 162, 257–265. [Google Scholar] [CrossRef]
- Fukayama, H.; Hatch, M.D.; Tamai, T.; Tsuchida, H.; Sudoh, S.; Furbank, R.T.; Miyao, M. Activity regulation and physiological impacts of maize C4-specific phosphoenolpyruvate carboxylase overproduced intransgenic rice plants. Photosyn Res. 2003, 77, 227–239. [Google Scholar] [CrossRef]
- Ku, M.S.; Agarie, S.; Nomura, M.; Fukayama, H.; Tsuchida, H.; Ono, K.; Hirose, S.; Toki, S.; Miyao, M.; Matsuoka, M. High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants. Nat. Biotechnol. 1999, 17, 76–80. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Stomatal Density (pcs) | Stomatal Size (mm) | LT (mm) | No. of BCB (pcs) | No. of SCB (pcs) | BCB Area (mm2) | SCB Area (mm2) | V Diameter (mm) | MC Size (mm2) |
|---|---|---|---|---|---|---|---|---|---|
| G164 | 3.667 a | 0.001 a | 0.100 a | 9 a | 46 a | 0.016 a | 0.0029 a | 0.0435 a | 2.4 × 10−5 b |
| C1 | 6.000 b | 0.004 b | 0.098 a | 20 b | 61 b | 0.015 a | 0.0028 a | 0.0345 b | 1.9 × 10−5 a |
| Varieties | Fv/Fm | qP | NPQ | ETR | ΦPSⅡ | Rubisco Activity (nmol/s/g) | Rubisco Content (ng/L) |
|---|---|---|---|---|---|---|---|
| C1 | 0.813 ± 0.006 a | 0.911 ± 0.011 a | 0.160 ± 0.055 a | 30.429 ± 0.432 a | 0.691 ± 0.009 a | 5.311 ± 9.74 b | 148.933 ± 2.95 a |
| G164 | 0.806 ± 0.002 a | 0.874 ± 0.023 a | 0.177 ± 0.042 a | 28.954 ± 0.887 a | 0.660 ± 0.019 a | 5.037 ± 2.43 a | 138.073 ± 4.28 a |
| KEGG Pathway | Protein Accession | Protein Description | Coverage [%] | Fold Change | Direction of Change | Subcellular Localization |
|---|---|---|---|---|---|---|
| Photosynthesis | O23877 | petH; ferredoxin—NADP+ reductase [EC:1.18.1.2] | 10.1 | 1.645 ** | up | chloroplast |
| P0C389 | petA; apocytochrome f | 46.2 | 0.764 ** | down | nucleus | |
| P12123 | petB; cytochrome b6 | 16.3 | 0.690 * | down | cytoplasm | |
| Q0DFC9 | petE; plastocyanin | 20.8 | 0.810 ** | down | chloroplast | |
| Q0J8M2 | petF; ferredoxin | 40.3 | 0.790 ** | down | chloroplast | |
| Q0JG75 | psb28; PSII 13kDa protein | 16.1 | 0.710 ** | down | chloroplast | |
| Q109L0 | psbP; photosystem II oxygen-evolving enhancer protein 2 | 17.7 | 0.645 ** | down | chloroplast | |
| Q10LV7 | psb27; photosystem II Psb27 protein | 34.5 | 0.745 ** | down | chloroplast | |
| Q69S39 | petC; cytochrome b6-f complex iron-sulfur subunit [EC:1.10.9.1] | 49.8 | 0.720 ** | down | chloroplast | |
| Q6ZFJ3 | petH; ferredoxin--NADP+ reductase [EC:1.18.1.2] | 56.3 | 0.770 ** | down | chloroplast | |
| Carbon fixation in photosynthetic organism | A0A0P0WP33 | PGK; phosphoglycerate kinase [EC:2.7.2.3] | 63.6 | 0.800 ** | down | chloroplast |
| A0A0P0XLG164 | PEPC; phosphoenolpyruvate carboxylase [EC:4.1.1.31] | 20.3 | 1.635 * | up | cytoplasm | |
| A0A0P0XUE4 | GPT; alanine transaminase [EC:2.6.1.2] | 22.4 | 1.425 * | up | cytoplasm | |
| Q0JJQ7 | E1.1.1.40; malate dehydrogenase (oxaloacetate-decarboxylating) (NADP+) [EC:1.1.1.40] | 29.1 | 1.500 ** | up | chloroplast | |
| Q2QTJ1 | rbcS; ribulose-bisphosphate carboxylase small chain [EC:4.1.1.39] | 53.1 | 0.790 ** | down | chloroplast | |
| Q42972 | MDH2; malate dehydrogenase [EC:1.1.1.37] | 15.7 | 1.465 ** | up | cytoplasm | |
| Q655Y9 | FBP; fructose-1,6-bisphosphatase I [EC:3.1.3.11] | 27.7 | 1.315 ** | up | chloroplast | |
| Q69K00 | TPI; triosephosphate isomerase (TIM) [EC:5.3.1.1] | 53 | 0.745 ** | down | chloroplast | |
| Q69UU3 | GGAT; glutamate--glyoxylate aminotransferase [EC:2.6.1.4 2.6.1.2 2.6.1.44] | 58.4 | 0.815 ** | down | peroxisome | |
| Q6H6C7 | PGK; phosphoglycerate kinase [EC:2.7.2.3] | 55.7 | 1.300 ** | up | chloroplast | |
| Q6YYW3 | E1.1.1.82; malate dehydrogenase (NADP+) [EC:1.1.1.82] | 47.8 | 0.795 ** | down | chloroplast | |
| Q7X8A1 | GAPA; glyceraldehyde-3-phosphate dehydrogenase (NADP+) (phosphorylating) [EC:1.2.1.13] | 51 | 0.775 ** | down | chloroplast | |
| Q7XDC8 | MDH1; malate dehydrogenase [EC:1.1.1.37] | 31.6 | 1.225 ** | up | cytoplasm | |
| Q7XZW5 | MDH2; malate dehydrogenase [EC:1.1.1.37] | 57.9 | 0.750 ** | down | chloroplast | |
| Q8H4V1 | pepc; phosphoenolpyruvate carboxylase [EC:4.1.1.31] | 35.7 | 1.310 ** | up | cytoplasm | |
| Q9SNK3 | GAPA; glyceraldehyde-3-phosphate dehydrogenase (NADP+) (phosphorylating) [EC:1.2.1.13] | 55 | 0.815 ** | down | chloroplast | |
| Q0J8A4 | GAPDH; glyceraldehyde 3-phosphate dehydrogenase [EC:1.2.1.12] | 64.1 | 1.390 ** | up | cytoplasm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Tang, M.; Wang, H.; Tao, W.; Wang, Q.; Chen, L.; Gao, G.; Lv, R.; Liang, T. Analysis of Photosynthetic Differences of Rice Germplasm in Southeast Asia Based on Leaf-Tissue Structure, Physiology, and iTRAQ. Agronomy 2022, 12, 3207. https://doi.org/10.3390/agronomy12123207
Zhang X, Tang M, Wang H, Tao W, Wang Q, Chen L, Gao G, Lv R, Liang T. Analysis of Photosynthetic Differences of Rice Germplasm in Southeast Asia Based on Leaf-Tissue Structure, Physiology, and iTRAQ. Agronomy. 2022; 12(12):3207. https://doi.org/10.3390/agronomy12123207
Chicago/Turabian StyleZhang, Xiaoli, Maoyan Tang, Hui Wang, Wei Tao, Qiang Wang, Lei Chen, Guoqing Gao, Ronghua Lv, and Tianfeng Liang. 2022. "Analysis of Photosynthetic Differences of Rice Germplasm in Southeast Asia Based on Leaf-Tissue Structure, Physiology, and iTRAQ" Agronomy 12, no. 12: 3207. https://doi.org/10.3390/agronomy12123207