
The Streptomyces chromofuscus Strain RFS-23 Induces Systemic Resistance and Activates Plant Defense Responses against Tomato Yellow Leaf Curl Virus Infection

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Plants, Viruses and Bacterial Isolates
2.2. Bacterial Treatment of N. benthamiana Plants and Viral Inoculation
2.3. Monitoring of Disease Symptoms and Samples Collection
2.4. Quantification of Relative Chlorophyll Contents
2.5. Nucleic acid Extractions cDNA Synthesis and RT-qPCR
2.6. Relative Accumulation of TYLCV Coat Protein
2.7. Biochemical Analysis of Antioxidant Enzymes
2.8. Statistical Analysis
3. Results
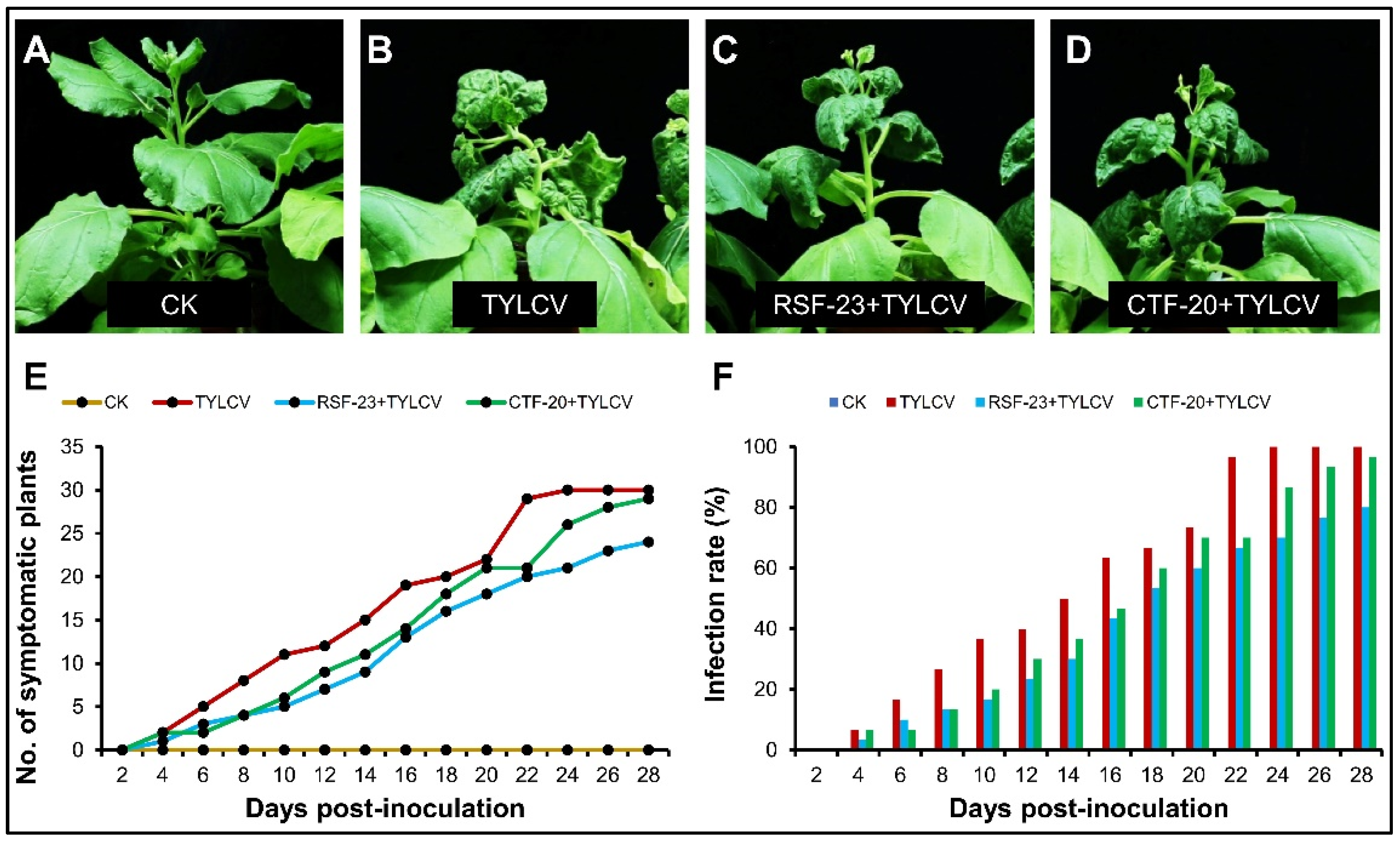
3.1. Comparative Effect of Streptomyces Strains on Development of Viral Disease
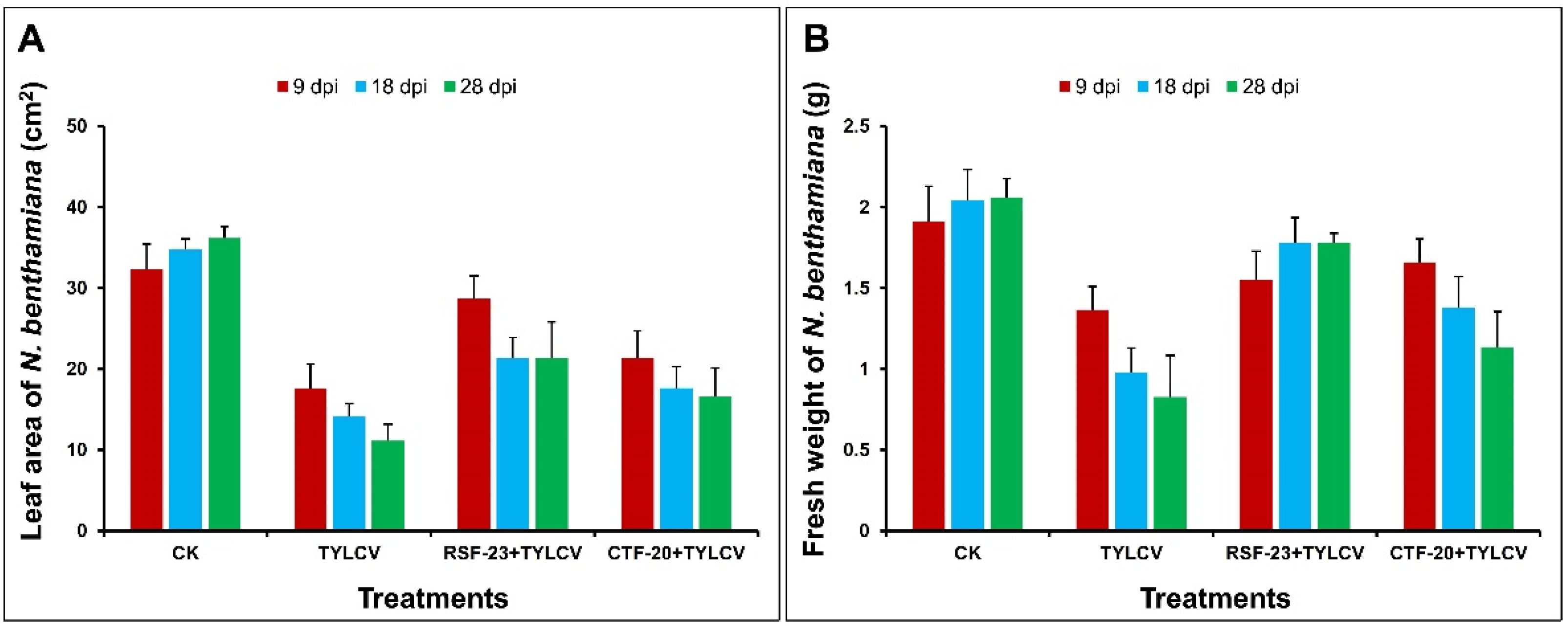
3.2. Estimation of Plant Growth Parameters
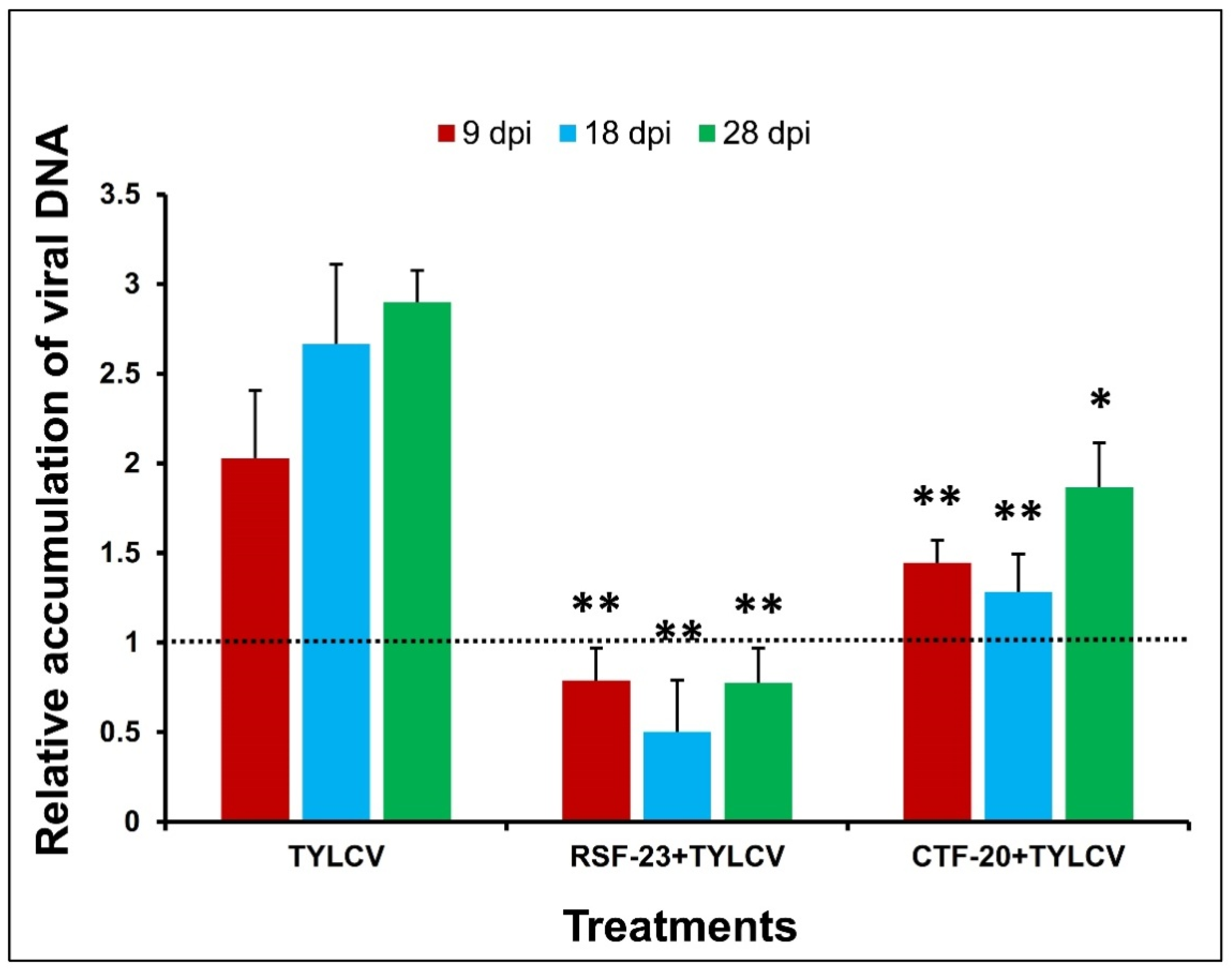
3.3. The Relative Accumulation of TYLCV Coat Protein
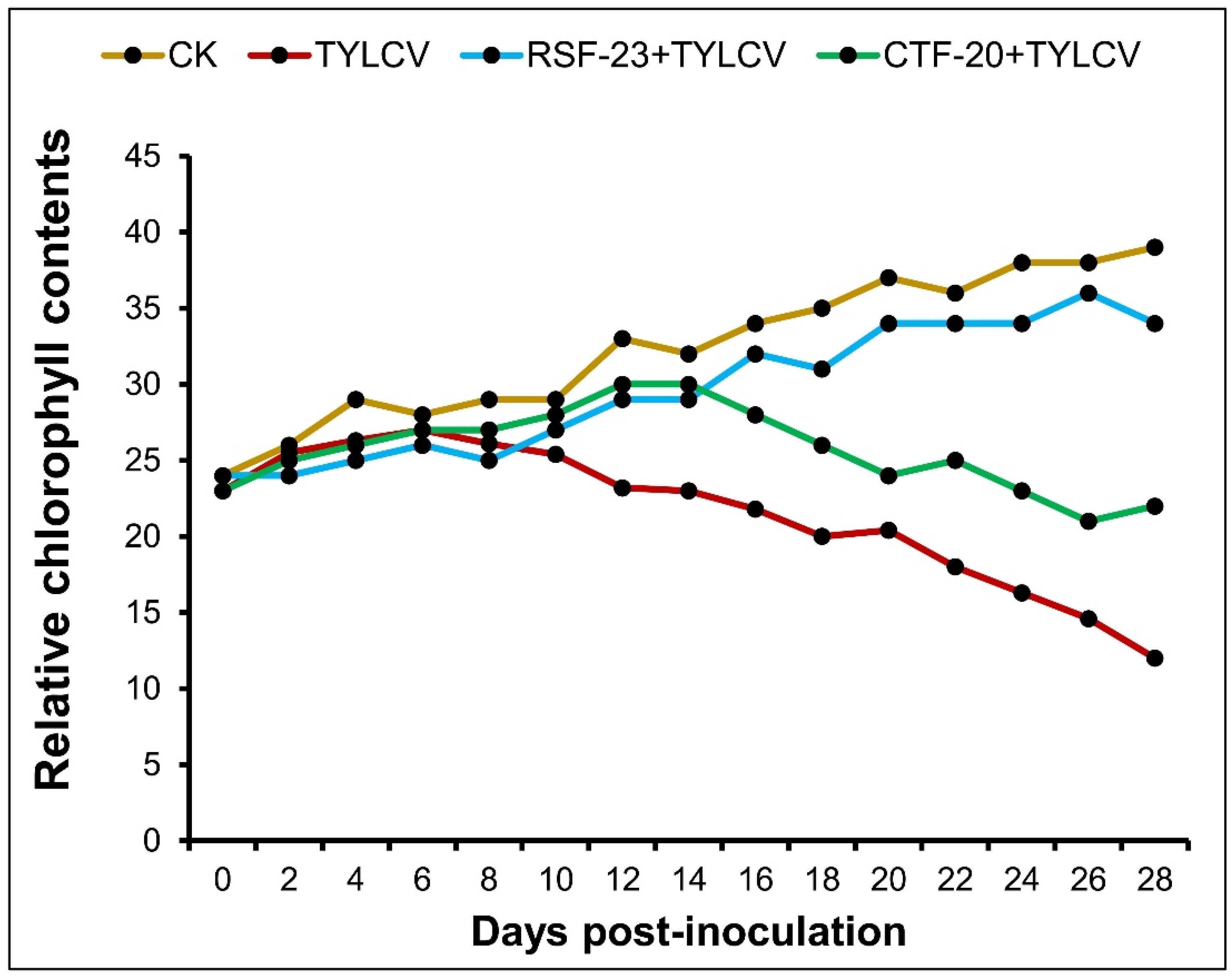
3.4. Determination of Relative Chlorophyll Contents
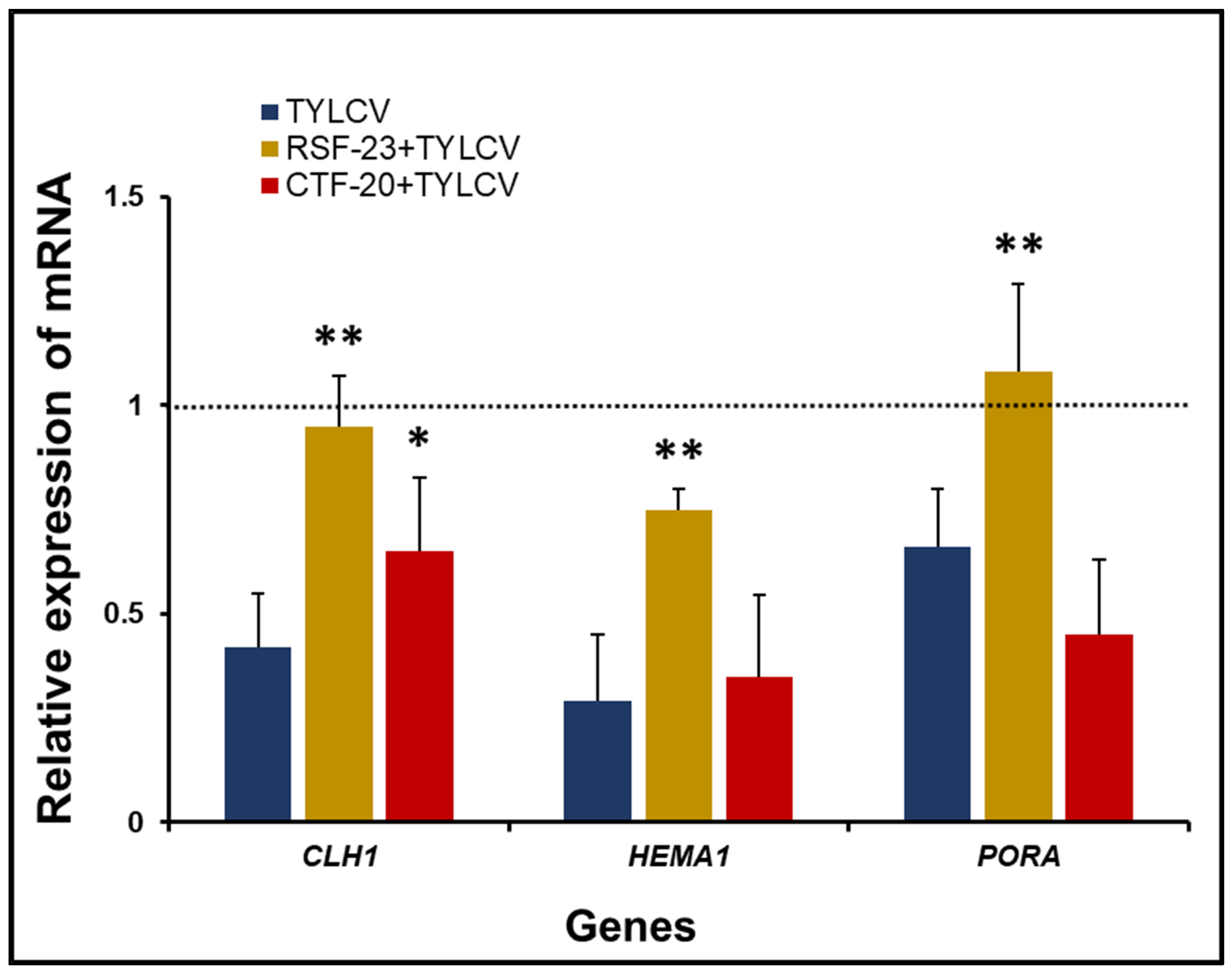
3.5. Comparative Effect of Bacterial (RSF-23 and CTF-20) Treatment on the Chlorophyl Biosynthesis Pathway
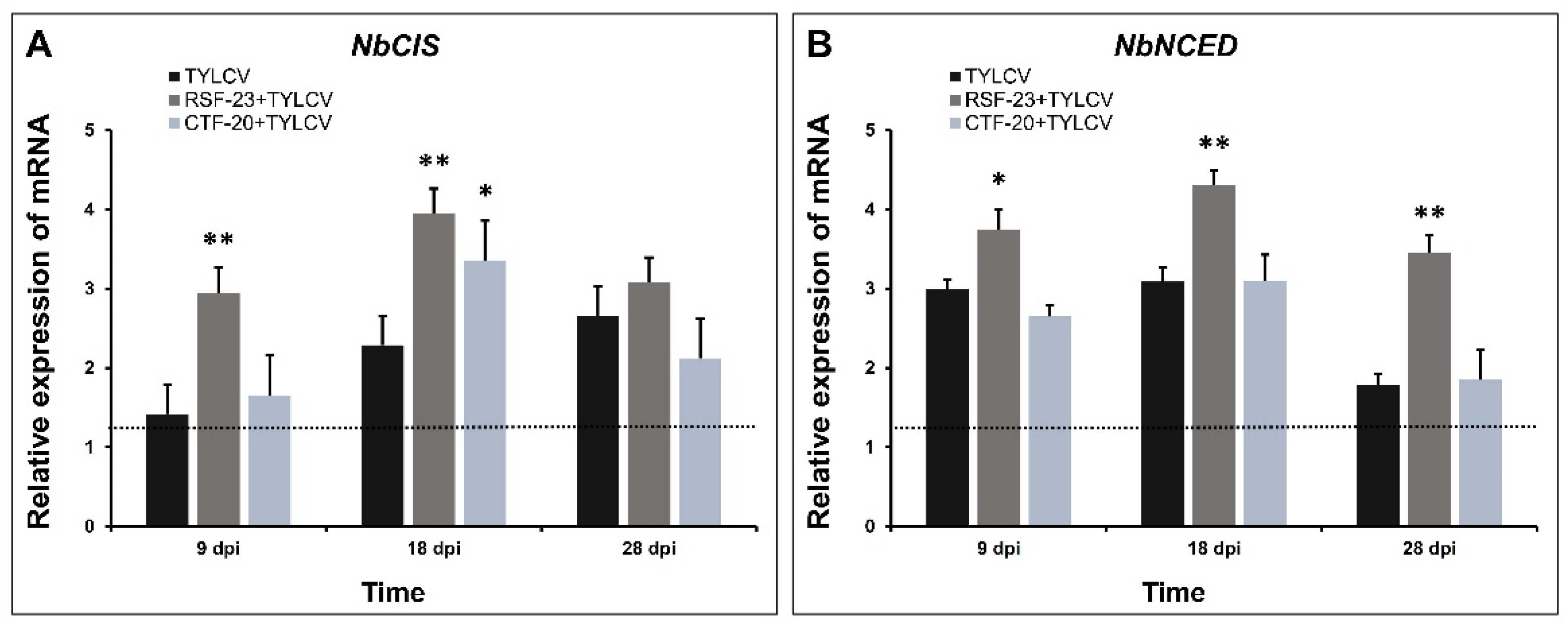
3.6. Activation of Defense-Related Genes in Response to Bacterial Treatment
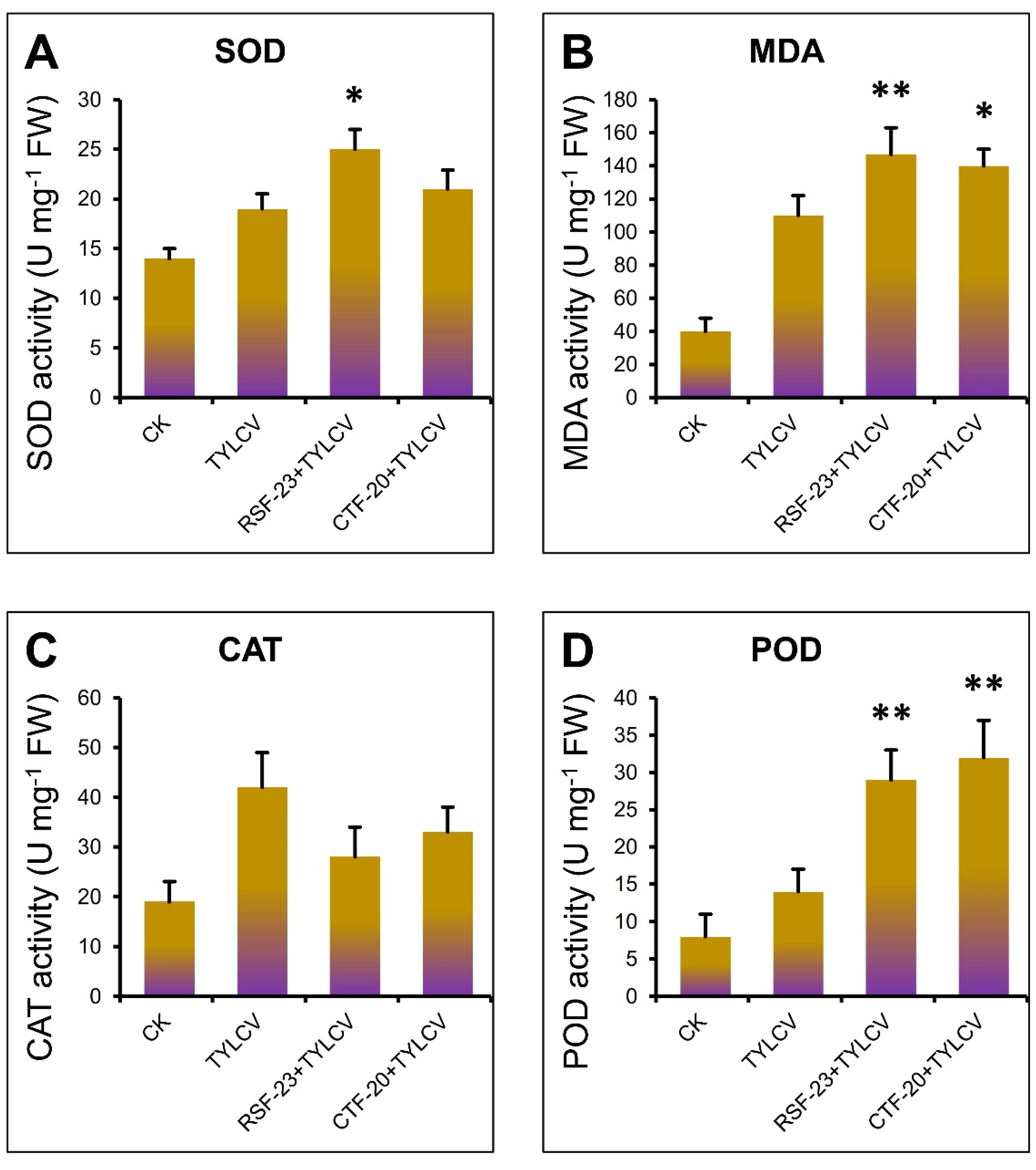
3.7. Activation of Antioxidant Enzymes by RSF-23 Strain in Response to TYLCV Infection
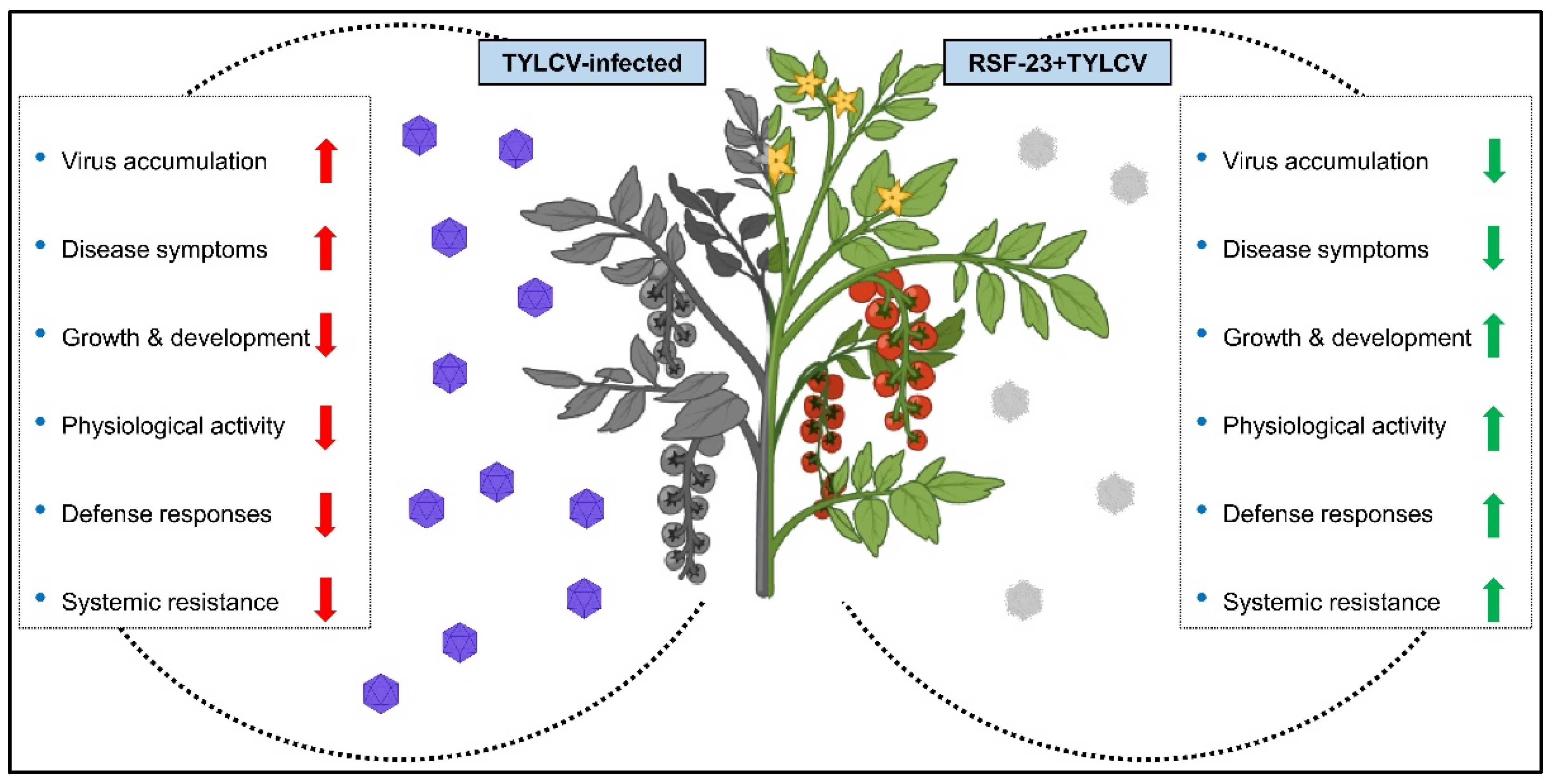
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdelkhalek, A.; Hafez, E. Plant viral diseases in Egypt and their control. In Cottage Industry of Biocontrol Agents and Their Applications; Springer: Cham, Switzerland, 2020; pp. 403–421. [Google Scholar]
- Hančinský, R.; Mihálik, D.; Mrkvová, M.; Candresse, T.; Glasa, M. Plant viruses infecting Solanaceae family members in the cultivated and wild environments: A Review. Plants 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant Sci. 2014, 5, 660. [Google Scholar] [CrossRef] [PubMed]
- Czosnek, H.; Laterrot, H. A worldwide survey of tomato yellow leaf curl viruses. Arch. Virol. 1997, 142, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, M.; Morin, S.; Zeidan, M.; Czosnek, H. Evidence for transovarial transmission of tomato yellow leaf curl virus by its vector, the whiteflyBemisia tabaci. Virology 1998, 240, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriones, E.; Navas-Castillo, J. Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide. Virus Res. 2000, 71, 123–134. [Google Scholar] [CrossRef]
- Papayiannis, L.; Katis, N.; Idris, A.; Brown, J. Identification of weed hosts of Tomato yellow leaf curl virus in Cyprus. Plant Dis. 2011, 95, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Anfoka, G.; Haj Ahmad, F.; Abhary, M.; Hussein, A. Detection and molecular characterization of viruses associated with tomato yellow leaf curl disease in cucurbit crops in Jordan. Plant Pathol. 2009, 58, 754–762. [Google Scholar] [CrossRef]
- Cohen, J.; Gera, A.; Ecker, R.; Ben-Joseph, R.; Perlsman, M.; Gokkes, M.; Lachman, O.; Antignus, Y. Lisianthus leaf curl-a new disease of lisianthus caused by tomato yellow leaf curl virus. Plant Dis. 1995, 79, 416–420. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Sánchez-Campos, S.; Díaz, J.A.; Sáez-Alonso, E.; Moriones, E. Tomato yellow leaf curl virus-Is causes a novel disease of common bean and severe epidemics in tomato in Spain. Plant Dis. 1999, 83, 29–32. [Google Scholar] [CrossRef] [Green Version]
- Reina, J.; Morilla, G.; Bejarano, E.; Rodríguez, M.; Janssen, D. First report of Capsicum annuum plants infected by tomato yellow leaf curl virus. Plant Dis. 1999, 83, 1176. [Google Scholar] [CrossRef]
- Albarracín Orio, A.G.; Brücher, E.; Ducasse, D.A. A strain of Bacillus subtilis subsp. subtilis shows a specific antagonistic activity against the soil-borne pathogen of onion Setophoma terrestris. Eur. J. Plant Pathol. 2016, 144, 217–223. [Google Scholar] [CrossRef]
- Fernando, W.G.D.; Ramarathnam, R.; Krishnamoorthy, A.S.; Savchuk, S.C. Identification and use of potential bacterial organic antifungal volatiles in biocontrol. Soil Biol. Biochem. 2005, 37, 955–964. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 inhibits Fusarium oxysporum growth and induces systemic resistance to cucumber mosaic virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Ozaki, K.; Hyakumachi, M. Induction of systemic resistance against Cucumber mosaic virus in Arabidopsis thaliana by Trichoderma asperellum SKT-1. Plant Pathol. J. 2013, 29, 193. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Chauhan, P.S.; Agrawal, L.; Raj, R.; Srivastava, A.; Gupta, S.; Mishra, S.K.; Yadav, S.; Singh, P.C.; Raj, S.K.; et al. Paenibacillus lentimorbus inoculation enhances tobacco growth and extenuates the virulence of Cucumber mosaic virus. PLoS ONE 2016, 11, e0149980. [Google Scholar] [CrossRef]
- Shen, L.; Wang, F.; Yang, J.; Qian, Y.; Dong, X.; Zhan, H. Control of tobacco mosaic virus by Pseudomonas fluorescens CZ powder in greenhouses and the field. Crop Prot. 2014, 56, 87–90. [Google Scholar] [CrossRef]
- Tan, L.T.-H.; Chan, K.-G.; Khan, T.M.; Bukhari, S.I.; Saokaew, S.; Duangjai, A.; Pusparajah, P.; Lee, L.-H.; Goh, B.-H. Streptomyces sp. MUM212 as a source of antioxidants with radical scavenging and metal chelating properties. Front. Pharmacol. 2017, 8, 276. [Google Scholar] [CrossRef]
- Hata, E.M.; Sijam, K.; Ahmad, Z.A.M.; Yusof, M.T.; Azman, N.A. In vitro Antimicrobial Assay of Actinomycetes in Rice Against Xanthomonas oryzae pv. oryzicola and as Potential Plant Growth Promoter. Braz. Arch. Biol. Technol. 2015, 58, 821–832. [Google Scholar] [CrossRef]
- Hasegawa, S.; Meguro, A.; Shimizu, M.; Nishimura, T.; Kunoh, H. Endophytic Actinomycetes and Their Interactions with Host Plants. Actinomycetologica 2006, 20, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Schrey, S.D.; Tarkka, M.T. Friends and foes: Streptomycetes as modulators of plant disease and symbiosis. Antonie Van Leeuwenhoek 2008, 94, 11–19. [Google Scholar] [CrossRef]
- Ara, I.; Bukhari, N.A.; Aref, N.; Shinwari, M.M.; Bakir, M. Antiviral activities of streptomycetes against tobacco mosaic virus (TMV) in Datura plant: Evaluation of different organic compounds in their metabolites. Afr. J. Biotechnol. 2012, 11, 2130–2138. [Google Scholar]
- Chen, J.; Liu, H.; Xia, Z.; Zhao, X.; Wu, Y.; An, M. Purification and structural analysis of the effective anti-TMV compound ε-poly-L-lysine produced by Streptomyces ahygroscopicus. Molecules 2019, 24, 1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Zaid, G.A.; Matar, S.M.; Abdelkhalek, A. Induction of Plant Resistance against Tobacco Mosaic Virus Using the Biocontrol Agent Streptomyces cellulosae Isolate Actino 48. Agronomy 2020, 10, 1620. [Google Scholar] [CrossRef]
- Alazem, M.; Lin, N.S. Roles of plant hormones in the regulation of host-virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting Ready for Battle. Mol. Plant-Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Norkunas, K.; Harding, R.; Dale, J.; Dugdale, B. Improving agroinfiltration-based transient gene expression in Nicotiana benthamiana. Plant Methods 2018, 14, 71. [Google Scholar] [CrossRef]
- Shirling, E.T.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef] [Green Version]
- Adeel, M.; Farooq, T.; White, J.C.; Hao, Y.; He, Z.; Rui, Y. Carbon-based nanomaterials suppress tobacco mosaic virus (TMV) infection and induce resistance in Nicotiana benthamiana. J. Hazard. Mater. 2021, 404, 124167. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Bardin, M.; Pugliese, M. Biocontrol Agents Against Diseases. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 385–407. [Google Scholar]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases–What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Farooq, T.; Adeel, M.; He, Z.; Umar, M.; Shakoor, N.; da Silva, W.; Elmer, W.; White, J.C.; Rui, Y. Nanotechnology and Plant Viruses: An Emerging Disease Management Approach for Resistant Pathogens. ACS Nano 2021, 15, 6030–6037. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- TariqJaveed, M.; Farooq, T.; Al-Hazmi, A.S.; Hussain, M.D.; Rehman, A.U. Role of Trichoderma as a biocontrol agent (BCA) of phytoparasitic nematodes and plant growth inducer. J. Invertebr. Pathol. 2021, 183, 107626. [Google Scholar] [CrossRef]
- Farooq, T.; Liu, D.; Zhou, X.; Yang, Q. Tomato Yellow Leaf Curl China Virus Impairs Photosynthesis in the Infected Nicotiana benthamiana with βC1 as an Aggravating Factor. Plant Pathol. J. 2019, 35, 521–529. [Google Scholar] [CrossRef]
- Marchant, W.G.; Gautam, S.; Hutton, S.F.; Srinivasan, R. Tomato Yellow Leaf Curl Virus-Resistant and -Susceptible Tomato Genotypes Similarly Impact the Virus Population Genetics. Front. Plant Sci. 2020, 11, 599697. [Google Scholar] [CrossRef]
- Prasad, A.; Sharma, N.; Hari-Gowthem, G.; Muthamilarasan, M.; Prasad, M. Tomato Yellow Leaf Curl Virus: Impact, Challenges, and Management. Trends Plant Sci. 2020, 25, 897–911. [Google Scholar] [CrossRef]
- Shteinberg, M.; Mishra, R.; Anfoka, G.; Altaleb, M.; Brotman, Y.; Moshelion, M.; Gorovits, R.; Czosnek, H. Tomato Yellow Leaf Curl Virus (TYLCV) Promotes Plant Tolerance to Drought. Cells 2021, 10, 2875. [Google Scholar] [CrossRef]
- Boukhatem, Z.F.; Merabet, C.; Tsaki, H. Plant Growth Promoting Actinobacteria, the Most Promising Candidates as Bioinoculants? Front. Agron. 2022, 4. [Google Scholar] [CrossRef]
- Farmer, P.B.; Uematsu, T.; Hogenkamp, H.P.; Suhadolnik, R.J. Nucleoside antibiotics. Epimerization of carbon 2′ of adenosine during the biosynthesis of 9- -D--arabinofuranosyladenine by Streptomyces antibioticus. J. Biol. Chem. 1973, 248, 1844–1847. [Google Scholar] [CrossRef]
- Igarashi, Y.; Trujillo, M.E.; Martínez-Molina, E.; Yanase, S.; Miyanaga, S.; Obata, T.; Sakurai, H.; Saiki, I.; Fujita, T.; Furumai, T. Antitumor anthraquinones from an endophytic actinomycete Micromonospora lupini sp. nov. Bioorg. Med. Chem. Lett. 2007, 17, 3702–3705. [Google Scholar] [CrossRef] [PubMed]
- de Reijke, T.M.; de Boer, E.C.; Schamhart, D.H.; Kurth, K.H. Immunostimulation in the urinary bladder by local application of Nocardia rubra cell wall skeleton preparation (Rubratin) for superficial bladder cancer immunotherapy—A phase I/II study. Urol. Res. 1997, 25, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Correction for Barka et al., Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. MMBR 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Berezin, V.; Abdukhakimova, D.; Trenozhnikova, L.; Bogoyavlenskiy, A.; Turmagambetova, A.; Issanov, A.; Azizan, A. Antiviral activities of extremophilic actinomycetes extracts from Kazakhstan’s unique ecosystems against influenza viruses and paramyxoviruses. Virol. J. 2019, 16, 150. [Google Scholar] [CrossRef]
- Djebaili, R.; Pellegrini, M.; Bernardi, M.; Smati, M.; Kitouni, M.; Del Gallo, M. Biocontrol Activity of Actinomycetes Strains against Fungal and Bacterial Pathogens of Solanum lycopersicum L. and Daucus carota L.: In Vitro and In Planta Antagonistic Activity. Biol. Life Sci. Forum 2021, 4, 27. [Google Scholar]
- Tran, T.M.; Ameye, M.; Devlieghere, F.; De Saeger, S.; Eeckhout, M.; Audenaert, K. Streptomyces Strains Promote Plant Growth and Induce Resistance Against Fusarium verticillioides via Transient Regulation of Auxin Signaling and Archetypal Defense Pathways in Maize Plants. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Alazem, M.; Kim, K.H.; Lin, N.S. Effects of Abscisic Acid and Salicylic Acid on Gene Expression in the Antiviral RNA Silencing Pathway in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 2538. [Google Scholar] [CrossRef] [Green Version]
- Alazem, M.; Lin, N.-S. Antiviral Roles of Abscisic Acid in Plants. Front. Plant Sci. 2017, 8, 1760. [Google Scholar] [CrossRef] [Green Version]
- Baebler, Š.; Witek, K.; Petek, M.; Stare, K.; Tušek-Žnidarič, M.; Pompe-Novak, M.; Renaut, J.; Szajko, K.; Strzelczyk-Żyta, D.; Marczewski, W.; et al. Salicylic acid is an indispensable component of the Ny-1 resistance-gene-mediated response against Potato virus Y infection in potato. J. Exp. Bot. 2014, 65, 1095–1109. [Google Scholar] [CrossRef]
- Bengtsson, T.; Weighill, D.; Proux-Wéra, E.; Levander, F.; Resjö, S.; Burra, D.D.; Moushib, L.I.; Hedley, P.E.; Liljeroth, E.; Jacobson, D.; et al. Proteomics and transcriptomics of the BABA-induced resistance response in potato using a novel functional annotation approach. BMC Genom. 2014, 15, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.; Khanna, K.; Manhas, R.K.; Bhardwaj, R.; Ohri, P.; Alkahtani, J.; Alwahibi, M.S.; Ahmad, P. Insights into the Role of Streptomyces hydrogenans as the Plant Growth Promoter, Photosynthetic Pigment Enhancer and Biocontrol Agent against Meloidogyne incognita in Solanum lycopersicum Seedlings. Plants 2020, 9, 1109. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, B.; Cai, S.; Zhang, Y.; Xu, M.; Zhang, C.; Yuan, B.; Xing, K.; Qin, S. Identification of Rhizospheric Actinomycete Streptomyces lavendulae SPS-33 and the Inhibitory Effect of its Volatile Organic Compounds against Ceratocystis fimbriata in Postharvest Sweet Potato (Ipomoea batatas (L.) Lam.). Microorganisms 2020, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akasaki, K.; Abe, H.; Seino, A.; Shirato, S. Yazumycin, a new antibiotic produced by Streptomyces lavendulae. J. Antibiot. 1968, 21, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groupé, V.; Frankel, J.W.; Lechevalier, M.P.; Waksman, S.A. Antiviral properties of ehrlichin, an antibiotic produced by Streptomyces lavendulae. J. Immunol. 1951, 67, 471–482. [Google Scholar]
- Hashimoto, K.; Nihira, T.; Sakuda, S.; Yamada, Y. IM-2, a butyrolactone autoregulator, induces production of several nucleoside antibiotics in Streptomyces sp. FRI-5. J. Ferment. Bioeng. 1992, 73, 449–455. [Google Scholar] [CrossRef]
- Saravana Kumar, P.; Al-Dhabi, N.A.; Duraipandiyan, V.; Balachandran, C.; Praveen Kumar, P.; Ignacimuthu, S. In vitro antimicrobial, antioxidant and cytotoxic properties of Streptomyces lavendulae strain SCA5. BMC Microbiol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Waksman, S.A.; Harris, D.; Lechevalier, M. Studies on Streptomyces lavendulae. J. Bacteriol. 1951, 62, 149–161. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Direction | Nucleotide Sequence (5′-3′) | Target Gene | Amplicon Size | Purpose | Reference |
|---|---|---|---|---|---|---|
| TYLCV-CP-F | Forward | ATCATGGACGTACAGGCC | CP | 200 bp | qPCR | this study |
| TYLCV-CP-R | Reverse | ACCTCTTACCAACTCTGTGA | qPCR | |||
| qGPDH-F | Forward | TCGACAGAGAAGGTGCCGGA | NbGAPDH | 190 bp | qPCR | this study |
| qGPDH-R | Reverse | TCAAGAACCTTGACAAAAGG | qPCR | |||
| qSA-F | Forward | GGCTCTGCTGTCTTCTTTACT | NbCIS | 200 bp | qPCR | [31] |
| qSA-R | Reverse | AGCTCATCGAACTCAACCTG | qPCR | |||
| qABA-F | Forward | CGTGGACTCTTTGGACTTGTT | NbNCED | 200 bp | qPCR | [31] |
| qABA-R | Reverse | GGGTGAGCTATCATTGTGGATT | qPCR | |||
| qChbio1-F | Forward | GTTCCAATTGGGGTTGGAA | CHL1 | 195 bp | qPCR | this study |
| qChbio1-R | Reverse | GAGATGTTGATTCTTATCT | qPCR | |||
| qChbio2-F | Forward | GGTGCGGTTTCGGTTAGCTCA | HEMA1 | 206 bp | qPCR | this study |
| qChbio2-R | Reverse | GGCATCTCCTCACGGATAGC | qPCR | |||
| qChbio3-F | Forward | GACTTGAAGAACTCCGAT | PORA | 200 bp | qPCR | this study |
| qChbio3-R | Reverse | TCTTTATACGCCTTTGCGCCA | qPCR |
| Analyzed Combination of Treatments | Time Post-Inoculation (days) | Morphological Parameter | |||
|---|---|---|---|---|---|
| Leaf Area | Fresh Weight | ||||
| Significant? | p-Value * | Significant? | p-Value * | ||
| CK vs. TYLCV | 9 | Yes | 0.009094 | Yes | 0.041102 |
| 18 | Yes | 0.000150 | Yes | 0.003412 | |
| 28 | Yes | 0.000141 | Yes | 0.003676 | |
| CK vs. RSF-23 + TYLCV | 9 | No | 0.297866 | No | 0.142864 |
| 18 | No | 0.039482 | No | 0.709585 | |
| 28 | Yes | 0.011114 | yes | 0.043202 | |
| CK vs. CTF-20 + TYLCV | 9 | Yes | 0.028906 | No | 0.241886 |
| 18 | Yes | 0.001175 | No | 0.025486 | |
| 28 | Yes | 0.001723 | Yes | 0.006770 | |
| TYLCV vs. RSF-23 + TYLCV | 9 | Yes | 0.020122 | No | 0.308886 |
| 18 | Yes | 0.002545 | Yes | 0.002980 | |
| 28 | Yes | 0.043614 | Yes | 0.007267 | |
| TYLCV vs. CTF-20 + TYLCV | 9 | No | 0.309201 | No | 0.110748 |
| 18 | No | 0.180613 | No | 0.083647 | |
| 28 | No | 0.121887 | No | 0.278941 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Ali, M.N.H.A.; Kamran, M.; Magsi, M.A.; Mora-Poblete, F.; Maldonado, C.; Waris, M.; Aljowaie, R.M.; Zehri, M.Y.; Elshikh, M.S. The Streptomyces chromofuscus Strain RFS-23 Induces Systemic Resistance and Activates Plant Defense Responses against Tomato Yellow Leaf Curl Virus Infection. Agronomy 2022, 12, 2419. https://doi.org/10.3390/agronomy12102419
Chen D, Ali MNHA, Kamran M, Magsi MA, Mora-Poblete F, Maldonado C, Waris M, Aljowaie RM, Zehri MY, Elshikh MS. The Streptomyces chromofuscus Strain RFS-23 Induces Systemic Resistance and Activates Plant Defense Responses against Tomato Yellow Leaf Curl Virus Infection. Agronomy. 2022; 12(10):2419. https://doi.org/10.3390/agronomy12102419
Chicago/Turabian StyleChen, Delai, Mian Noor Hussain Asghar Ali, Muhammad Kamran, Manzoor Ali Magsi, Freddy Mora-Poblete, Carlos Maldonado, Muhammad Waris, Reem M. Aljowaie, Mohammad Yakoob Zehri, and Mohamed S. Elshikh. 2022. "The Streptomyces chromofuscus Strain RFS-23 Induces Systemic Resistance and Activates Plant Defense Responses against Tomato Yellow Leaf Curl Virus Infection" Agronomy 12, no. 10: 2419. https://doi.org/10.3390/agronomy12102419