
Endophytic Candida membranifaciens from Euphorbia milii L. Alleviate Salt Stress Damages in Maize

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Fungal Endophyte
2.2. Fungal Isolates Screening for PGP Traits
2.3. Halo-Tolerance Screening of Fungal Isolates
2.4. Fungus Evaluation for the Production IAA, Siderophore and Phosphate Solubilization
2.5. Estimation of Phenol, Flavonoid in the CF of Endophytic Fungi
2.6. Molecular Identification of the Fungal Isolate
2.7. Evaluation of Fungi Effect on Maize Growth under Salt Stress
2.8. Estimation of Proline, Catalase and Peroxidase Activities
2.9. Total Phenolics, Flavonoids and Phytohormones Estimation in Maize
2.10. LCMS Data Analysis of Endophytic Fungal Culture Filtrate
2.11. Statistical Analysis
3. Results
3.1. Isolation of FH15 from Euphorbia Milii
3.2. Physiochemical Traits of FH15 and Salt Tolerance
3.3. Molecular Identification and Phylogenetic Analysis of FH15 Isolate
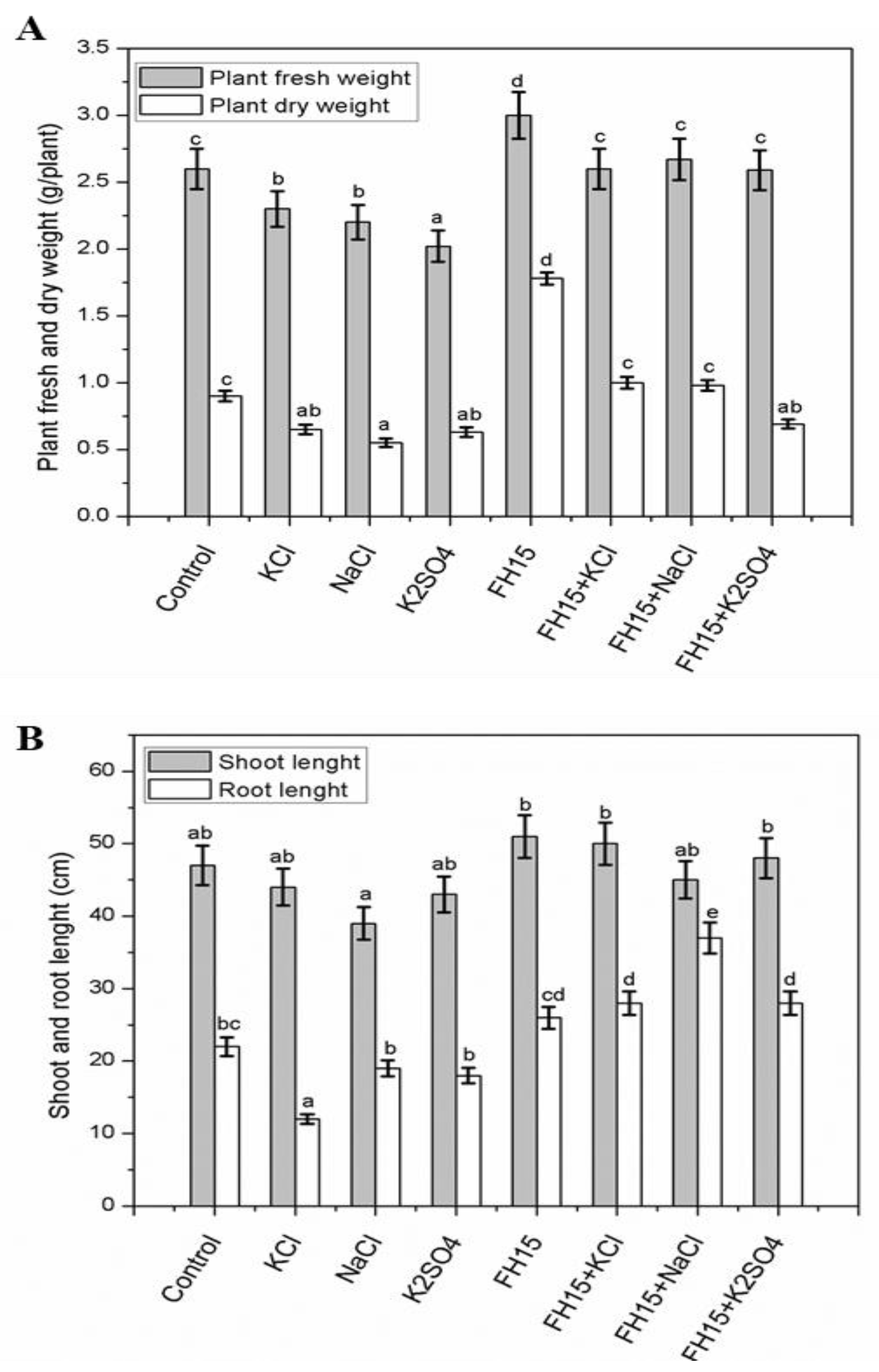
3.4. C. membranifaciens FH15 Augment Maize Plants under Salt Stress
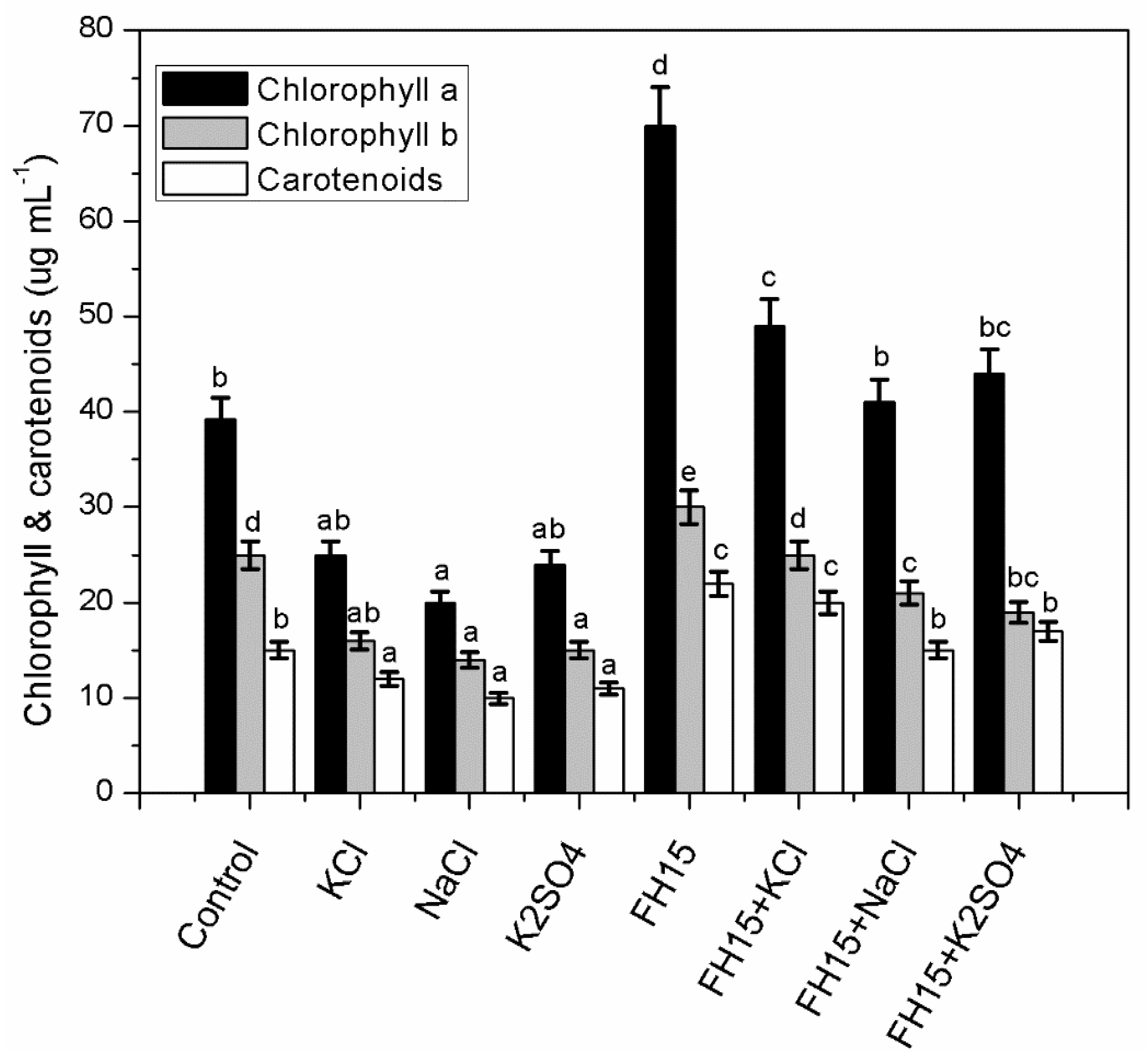
3.5. C. membranifaciens FH15 Improved Chlorophyll and Carotenoids Content in Maize Plants
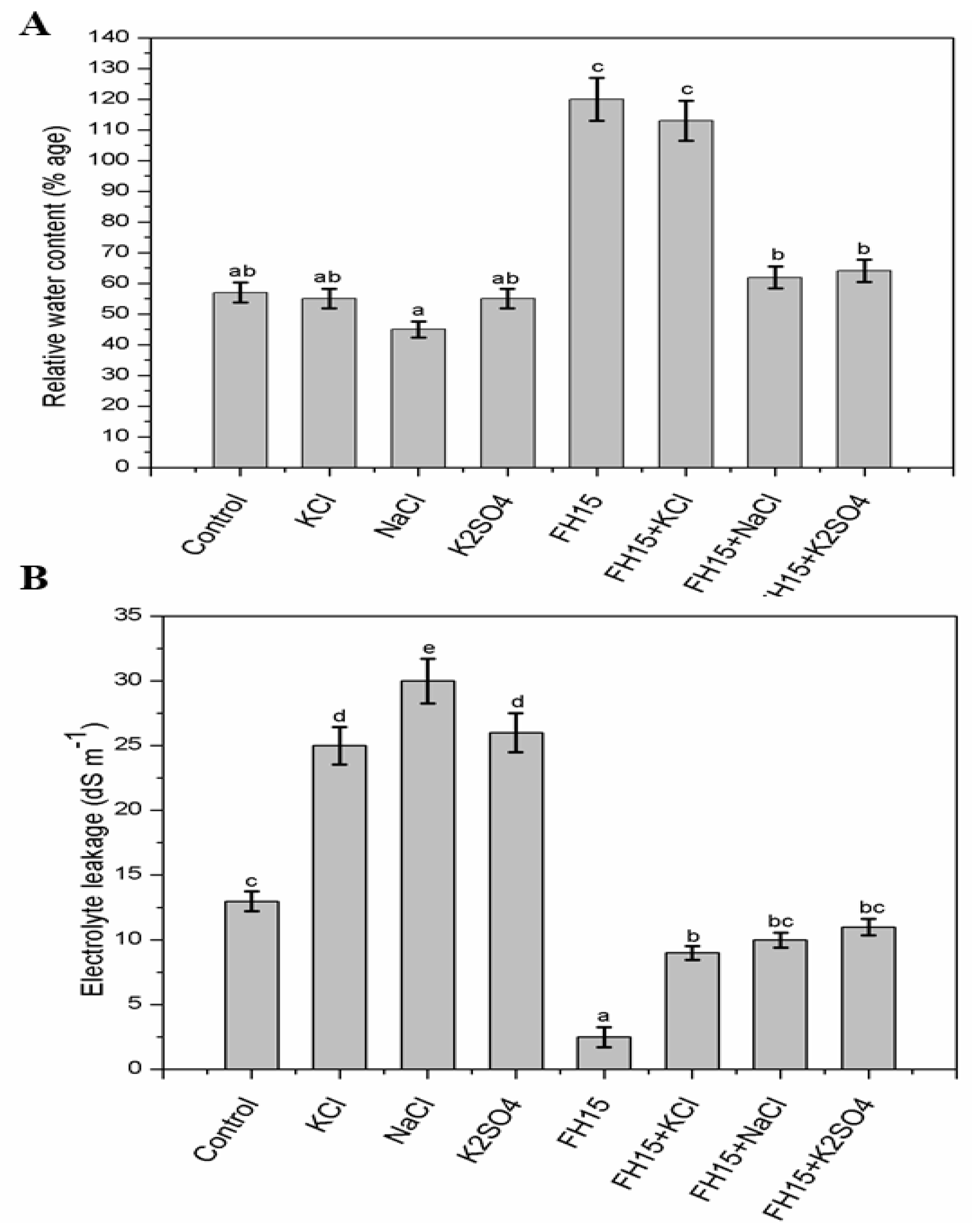
3.6. C. membranifaciens FH15 Improved RWC and Electrolyte Leakage in Maize Plants
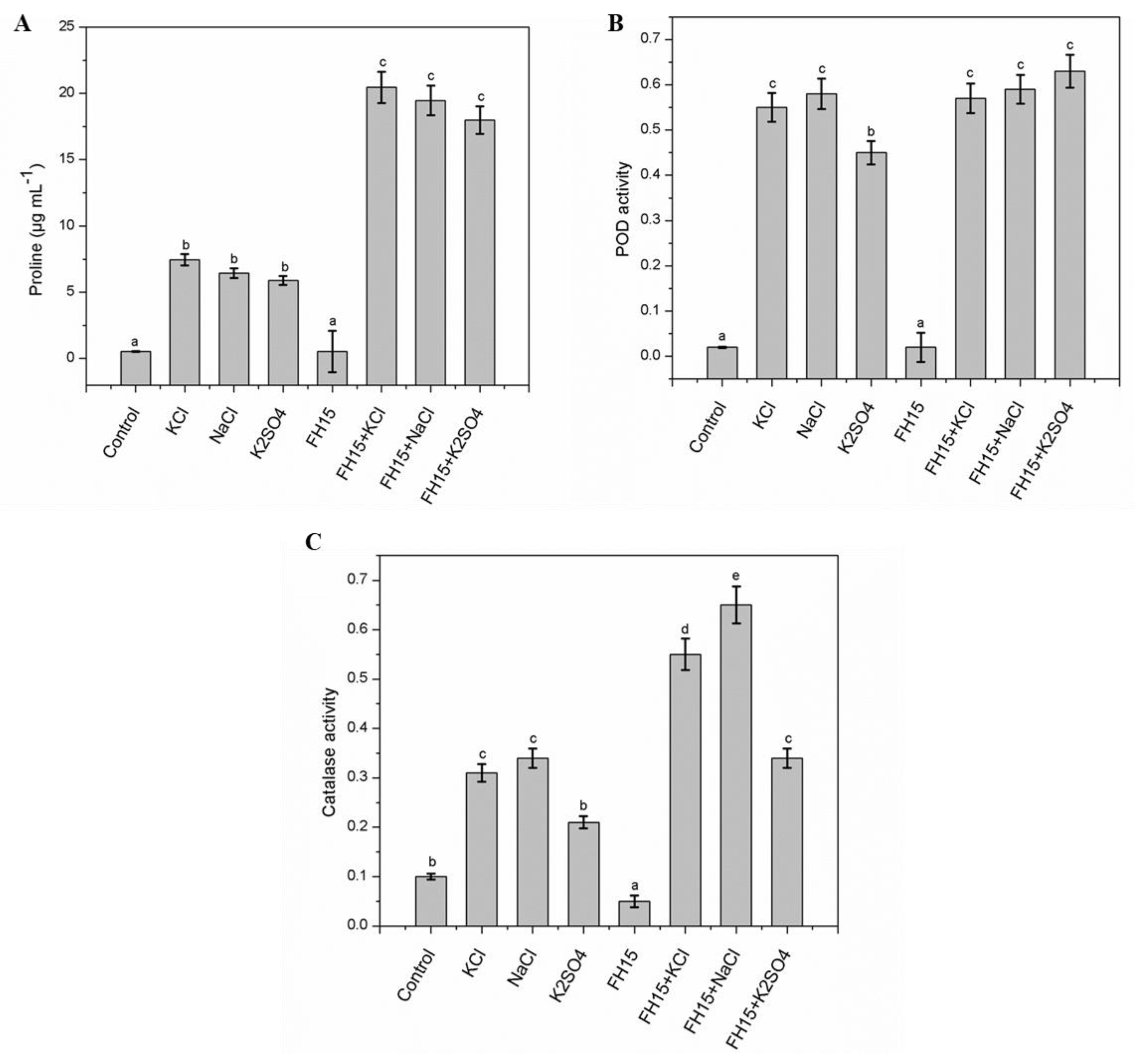
3.7. Effect of C. membranifaciens FH15 on Proline Content and Antioxidant Enzymes System
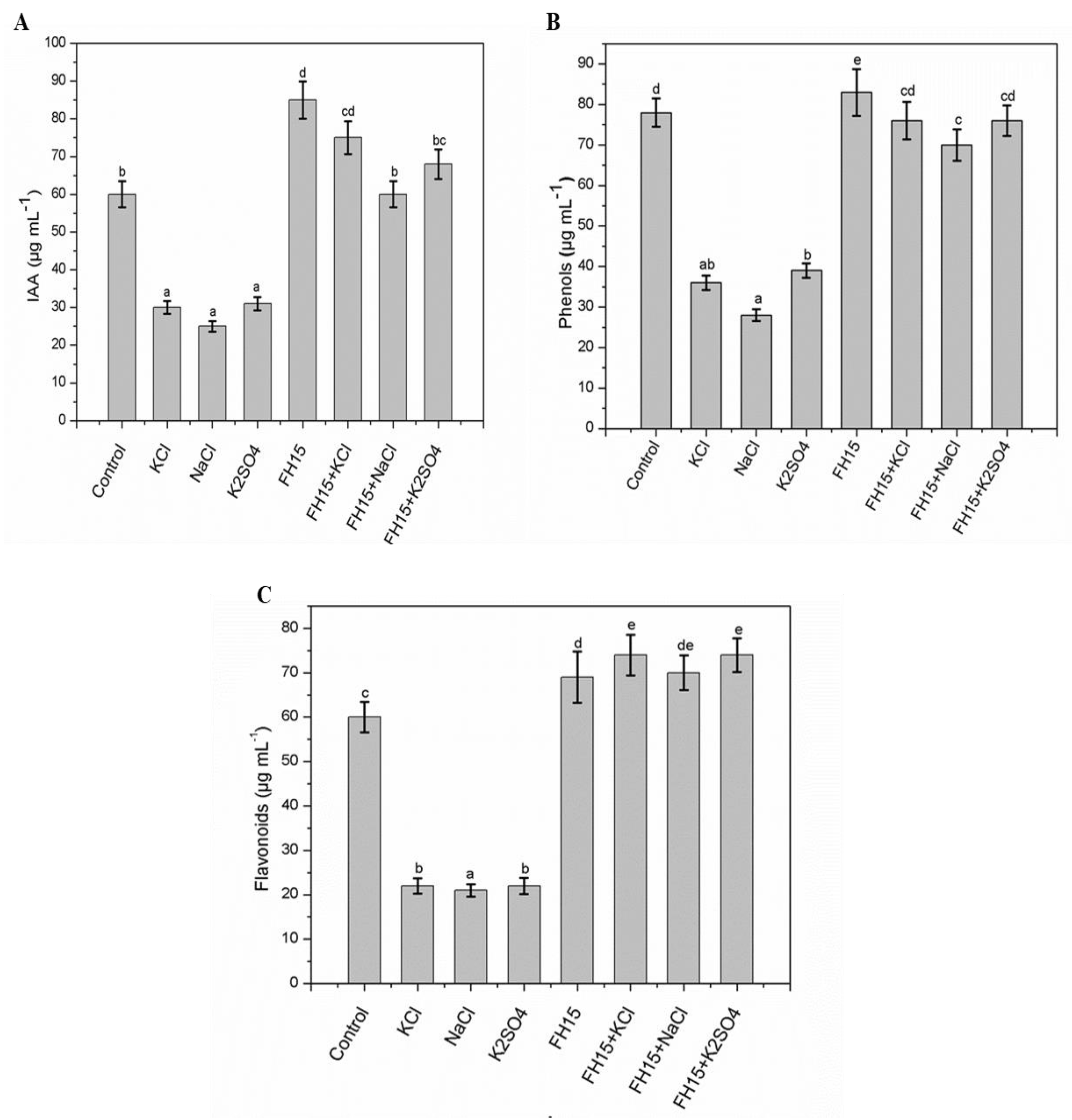
3.8. Effect of C. membranifaciens FH15 on IAA, Total Flavonoids and Phenolics
3.9. Identification of Compounds in C. membranifaciens FH15 in the CF by LC-ESI-MS/MS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ikram, M.; Ali, N.; Jan, G.; Iqbal, A.; Hamayun, M.; Jan, F.G.; Hussain, A.; Lee, I.-J. Trichoderma reesei improved the nutrition status of wheat crop under salt stress. J. Plant Interact. 2019, 14, 590–602. [Google Scholar] [CrossRef]
- Gul, S.L.; Moon, Y.-S.; Hamayun, M.; Khan, S.A.; Iqbal, A.; Khan, M.A.; Hussain, A.; Shafique, M.; Kim, Y.-H.; Ali, S.J.C.M. Porostereum spadiceum-AGH786 Regulates the Growth and Metabolites Production in Triticum aestivum L. Under Salt Stress. Curr. Microbiol. 2022, 79, 159. [Google Scholar] [CrossRef] [PubMed]
- Bouzouina, M.; Kouadria, R.; Lotmani, B. Fungal endophytes alleviate salt stress in wheat in terms of growth, ion homeostasis and osmoregulation. J. Appl. Microbiol. 2021, 130, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Khan, S.A.; Hamayun, M.; Iqbal, A.; Khan, A.L.; Hussain, A.; Shah, M. Endophytic fungi from Caralluma acutangula can secrete plant growth promoting enzymes. Fresenius Env. Bull 2019, 28, 2688–2696. [Google Scholar]
- Miransari, M. Hyperaccumulators, arbuscular mycorrhizal fungi and stress of heavy metals. Biotechnol. Adv. 2011, 29, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; Woodward, C.J.; Redman, R.S. Fungal influence on plant tolerance to stress. Biocomplexity Plant-Fungal Interact. 2012, 6, 155–163. [Google Scholar]
- Rai, M.; Rathod, D.; Agarkar, G.; Dar, M.; Brestic, M.; Pastore, G.M.; Junior, M.R.M. Fungal growth promotor endophytes: A pragmatic approach towards sustainable food and agriculture. Symbiosis 2014, 62, 63–79. [Google Scholar] [CrossRef]
- Zou, W.; Tan, R. Advances in Plant Science; China Higher Education Press: Beijing, China, 1999; Volume 2, pp. 183–190. [Google Scholar]
- Souchie, E.L.; Azcón, R.; Barea, J.M.; Saggin-Júnior, O.J.; Silva, E.M.R.D. Phosphate solubilization and synergism between P-solubilizing and arbuscular mycorrhizal fungi. Pesqui. Agropecuária Bras. 2006, 41, 1405–1411. [Google Scholar] [CrossRef]
- Haselwandter, K. Structure and function of siderophores produced by mycorrhizal fungi. Mineral. Mag. 2008, 72, 61–64. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Ali, S.; Moon, Y.-S.; Hamayun, M.; Khan, M.A.; Bibi, K.; Lee, I.-J. Pragmatic role of microbial plant biostimulants in abiotic stress relief in crop plants. J. Plant Interact. 2022, 17, 705–718. [Google Scholar] [CrossRef]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Lee, I.-J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, A.E. Endophytic fungi: Hidden Components of Tropical Community Ecology. In Tropical Forest Community Ecology; Carson, W.P., Schnitzer, S.A., Eds.; Blackwell Publishing Ltd: West Sussex, UK, 2008; pp. 178–188. [Google Scholar]
- Harper, J.K.; Arif, A.M.; Ford, E.J.; Strobel, G.A.; Porco, J.A., Jr.; Tomer, D.P.; Oneill, K.L.; Heider, E.M.; Grant, D.M. Pestacin: A 1, 3-dihydro isobenzofuran from Pestalotiopsis microspora possessing antioxidant and antimycotic activities. Tetrahedron 2003, 59, 2471–2476. [Google Scholar] [CrossRef]
- Sannigrahi, S.; Mazumder, U.K.; Pal, D.; Mishra, S.L. Hepatoprotective potential of methanol extract of Clerodendrum infortunatum Linn. against CCl4 induced hepatotoxicity in rats. Pharmacogn. Mag. 2009, 5, 394. [Google Scholar]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.-Y.; Suh, S.-J.; Hwang, S.-K.; Kim, J.-M.; Lee, I.-J.; Choo, Y.-S.; Yoon, U.-H. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hückelhoven, R.; Neumann, C.; Von Wettstein, D. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef]
- Yuan, Z.-l.; Zhang, C.-l.; Lin, F.-c. Role of diverse non-systemic fungal endophytes in plant performance and response to stress: Progress and approaches. J. Plant Growth Regul. 2010, 29, 116–126. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Radhakrishnan, R.; Waqas, M.; Kang, S.-M.; Kim, Y.-H.; Shin, J.-H.; Choo, Y.-S.; Kim, J.-G.; Lee, I.-J. Mutualistic association of Paecilomyces formosus LHL10 offers thermotolerance to Cucumis sativus. Antonie Van Leeuwenhoek 2012, 101, 267–279. [Google Scholar] [CrossRef]
- Strobel, G.A. Endophytes as sources of bioactive products. Microbes Infect. 2003, 5, 535–544. [Google Scholar] [CrossRef]
- Mehdi, S.; Ahsan, M. Genetic coefficient of variation, relative expected genetic advance and inter-relationships in maize (Zea mays L.) for green fodder purposes at seedling stage. Pak. J. Biol. Sci. 2000, 3, 1890–1891. [Google Scholar]
- Photita, W.; Lumyong, S.; Lumyong, P.; McKenzie, E.; Hyde, K. Are some endophytes of Musa acuminata latent pathogens? Fungal Divers. 2004, 16, 131–140. [Google Scholar]
- Jan, F.G.; Hamayun, M.; Moon, Y.-S.; Jan, G.; Shafique, M.; Ali, S.J.B. Endophytic aspergillus oryzae reprograms Abelmoschus esculentus L. to higher growth under salt stress via regulation of physiochemical attributes and antioxidant system. Biologia 2022, 1–14. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Khan, S.A.; Kim, H.-Y.; Khan, A.L.; Waqas, M.; Irshad, M.; Iqbal, A.; Rehman, G.; Jan, S. Gibberellins producing endophytic fungus Porostereum spadiceum AGH786 rescues growth of salt affected soybean. Front. Microbiol. 2017, 8, 686. [Google Scholar] [CrossRef] [PubMed]
- Chadha, N.; Prasad, R.; Varma, A. Plant Promoting Activities of Fungal Endophytes Associated with Tomato Roots from Central Himalaya, India and Their Interaction with Piriformospora Indica. IJPBS 2015, 6, 333–343. [Google Scholar]
- Bhalodia, N.R.; Acharya, R.; Shukla, V. Evaluation of in vitro Antioxidant Activity of hydroalcoholic seed extratcs of Cassia fistula linn. Free Radic. Antioxid. 2011, 1, 68–76. [Google Scholar] [CrossRef]
- Akbay, P.; Basaran, A.A.; Undeger, U.; Basaran, N. In vitro immunomodulatory activity of flavonoid glycosides from Urtica dioica L. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2003, 17, 34–37. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Hamayun, M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Co-synergism of endophyte Penicillium resedanum LK6 with salicylic acid helped Capsicum annuum in biomass recovery and osmotic stress mitigation. BMC Microbiol. 2013, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, A.A.; Saadatmand, S.; Niknam, V.; Nejadsatari, T.; Babaeizad, V. Effect of endophytic fungus, Piriformospora indica, on growth and activity of antioxidant enzymes of rice (Oryza sativa L.) under salinity stress. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 1337–1350. [Google Scholar]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmüller, R. The endophytic fungus Piriformospora indica stimulates the expression of nitrate reductase and the starch-degrading enzyme glucan-water dikinase in tobacco and Arabidopsis roots through a homeodomain transcription factor that binds to a conserved motif in their promoters. J. Biol. Chem. 2005, 280, 26241–26247. [Google Scholar]
- Gurupavithra, S.; Jayachitra, A. Isolation and identification of endophytic fungi from Ocimum sanctum and analyse its antioxidant properties. Int. J. Resear. Pharmaceut. Biomed. Sci 2013, 4, 1120–1125. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Khan, I.; Rahman, H.; El-Salam, N.M.A.; Tawab, A.; Hussain, A.; Khan, T.A.; Khan, U.A.; Qasim, M.; Adnan, M.; Azizullah, A. Punica granatum peel extracts: HPLC fractionation and LC MS analysis to quest compounds having activity against multidrug resistant bacteria. BMC Complementary Altern. Med. 2017, 17, 247. [Google Scholar] [CrossRef] [PubMed]
- Jebamony, J.R.; Thomas Muthiah, P. 8-Hydroxyquinolinium–Salicylate–Salicylic Acid (1/1/1) Complex, C9H8NO+. C7H5O3−. C7H6O3. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1998, 54, 539–540. [Google Scholar] [CrossRef]
- Jiang, C.; Liang, G.; Ren, Y.; Xu, T.; Song, Y.; Jin, W. An UPLC-MS/MS method for simultaneous quantification of the components of Shenyanyihao oral solution in rat plasma. BioMed Res. Int. 2020, 2020, 4769267. [Google Scholar] [CrossRef]
- Said, A.; Aboutabl, E.A.; Melek, F.R.; Abdel Jaleel Raheem Abdel Jaleel, G.; Raslan, M. Phytoconstituents profiling of Cissus rotundifolia (Forssk.) Vahl. by HPLC-MS/MS, and evaluation of its free radical scavenging activity (DPPH) and cytotoxicity. Trends Phytochem. Res. 2018, 2, 65–74. [Google Scholar]
- Ghareeb, M.A.; Sobeh, M.; Rezq, S.; El-Shazly, A.M.; Mahmoud, M.F.; Wink, M. HPLC-ESI-MS/MS profiling of polyphenolics of a leaf extract from Alpinia zerumbet (Zingiberaceae) and its anti-inflammatory, anti-nociceptive, and antipyretic activities in vivo. Molecules 2018, 23, 3238. [Google Scholar] [CrossRef] [PubMed]
- Sisó-Terraza, P.; Luis-Villarroya, A.; Fourcroy, P.; Briat, J.-F.; Abadía, A.; Gaymard, F.; Abadía, J.; Álvarez-Fernández, A. Accumulation and secretion of coumarinolignans and other coumarins in Arabidopsis thaliana roots in response to iron deficiency at high pH. Front. Plant Sci. 2016, 7, 1711. [Google Scholar] [CrossRef]
- Bilia, A.R.; Rubio, M.d.M.E.; Alvarez, M.L.; Morelli, I.; Gonzalez, J.M. New benzyl alcohol glycosides from Pyrus bourgaeana. Planta Med. 1994, 60, 569–571. [Google Scholar] [CrossRef]
- Beelders, T.; De Beer, D.; Stander, M.A.; Joubert, E. Comprehensive phenolic profiling of Cyclopia genistoides (L.) Vent. by LC-DAD-MS and-MS/MS reveals novel xanthone and benzophenone constituents. Molecules 2014, 19, 11760–11790. [Google Scholar] [CrossRef] [PubMed]
- Koolen, H.H.F.; da Silva, F.M.A.; Gozzo, F.C.; de Souza, A.Q.L.; de Souza, A.D.L. Antioxidant, antimicrobial activities and characterization of phenolic compounds from buriti (Mauritia flexuosa L. f.) by UPLC–ESI-MS/MS. Food Res. Int. 2013, 51, 467–473. [Google Scholar] [CrossRef]
- Bonta, R.K. Dietary phenolic acids and flavonoids as potential anti-cancer agents: Current state of the art and future perspectives. Anti Cancer Agents Med. Chem. 2020, 20, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Kaleta, E.; Wang, P. Simultaneous quantitation of 78 drugs and metabolites in urine with a dilute-and-shoot LC–MS-MS assay. J. Anal. Toxicol. 2015, 39, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K. The future products: Endophytic fungal metabolites. J. Biodivers. Bio Prospect. Dev. 2015, 2, 2–7. [Google Scholar]
- Tenguria, R.K.; Khan, F.N.; Quereshi, S. Endophytes-mines of pharmacological therapeutics. World J. Sci. Technol. 2011, 1, 127–149. [Google Scholar]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, I.-J. Ameliorative symbiosis of endophyte (Penicillium funiculosum LHL06) under salt stress elevated plant growth of Glycine max L. Plant Physiol. Biochem. 2011, 49, 852–861. [Google Scholar] [CrossRef]
- Mastouri, F.; Björkman, T.; Harman, G.E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit. Mol. Plant-Microbe Interact. 2012, 25, 1264–1271. [Google Scholar] [CrossRef]
- Singh, P.K.; Gautam, S. Role of salicylic acid on physiological and biochemical mechanism of salinity stress tolerance in plants. Acta Physiol. Plant. 2013, 35, 2345–2353. [Google Scholar] [CrossRef]
- Qi, W.-Z.; Liu, H.-H.; Liu, P.; Dong, S.-T.; Zhao, B.-Q.; So, H.B.; Li, G.; Liu, H.-D.; Zhang, J.-W.; Zhao, B. Morphological and physiological characteristics of corn (Zea mays L.) roots from cultivars with different yield potentials. Eur. J. Agron. 2012, 38, 54–63. [Google Scholar] [CrossRef]
- Rawat, L.; Singh, Y.; Shukla, N.; Kumar, J. Alleviation of the adverse effects of salinity stress in wheat (Triticum aestivum L.) by seed biopriming with salinity tolerant isolates of Trichoderma harzianum. Plant Soil 2011, 347, 387–400. [Google Scholar] [CrossRef]
- Zhang, F.; Yuan, J.; Yang, X.; Cui, Y.; Chen, L.; Ran, W.; Shen, Q. Putative Trichoderma harzianum mutant promotes cucumber growth by enhanced production of indole acetic acid and plant colonization. Plant Soil 2013, 368, 433–444. [Google Scholar] [CrossRef]
- Paixão Resende, M.; Jakoby, I.C.M.C.; dos Santos, L.C.R.; Soares, M.A.; Pereira, F.D.; Souchie, E.L.; Silva, F.G. Phosphate solubilization and phytohormone production by endophytic and rhizosphere Trichoderma isolates of guanandi (Calophyllum brasiliense Cambess). Afr. J. Microbiol. Res. 2014, 8, 2616–2623. [Google Scholar]
- Iqbal, M.; Ashraf, M. Alleviation of salinity-induced perturbations in ionic and hormonal concentrations in spring wheat through seed preconditioning in synthetic auxins. Acta Physiol. Plant. 2013, 35, 1093–1112. [Google Scholar] [CrossRef]
- Behera, R.K.; Mishra, P.C.; Choudhury, N.K. High irradiance and water stress induce alterations in pigment composition and chloroplast activities of primary wheat leaves. J. Plant Physiol. 2002, 159, 967–973. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.; Sharma, S. Antioxidant defense system, lipid peroxidation, proline-metabolizing enzymes, and biochemical activities in two Morus alba genotypes subjected to NaCl stress. Russ. J. Plant Physiol. 2010, 57, 509–517. [Google Scholar] [CrossRef]
- Sarwat, M.; Hashem, A.; Ahanger, M.A.; Abd_Allah, E.F.; Alqarawi, A.; Alyemeni, M.N.; Ahmad, P.; Gucel, S. Mitigation of NaCl stress by arbuscular mycorrhizal fungi through the modulation of osmolytes, antioxidants and secondary metabolites in mustard (Brassica juncea L.) plants. Front. Plant Sci. 2016, 7, 869. [Google Scholar] [CrossRef]
- Singh, A. The physiology of salt tolerance in four genotypes of chickpea during germination. J. Agric. Sci. 2018, 6, 87–93. [Google Scholar]
- Amanullah, M.M.; Sekar, S.; Vincent, S. Plant growth substances in crop production: A review. Asian J. Plant Sci. 2010, 9, 215–222. [Google Scholar] [CrossRef]
- Chen, F.; Long, X.; Liu, Z.; Shao, H.; Liu, L. Analysis of phenolic acids of Jerusalem artichoke (Helianthus tuberosus L.) responding to salt-stress by liquid chromatography/tandem mass spectrometry. Sci. World J. 2014, 2014, 568043. [Google Scholar] [CrossRef]
- Tsuji, Y.; Chen, F.; Yasuda, S.; Fukushima, K. Unexpected behavior of coniferin in lignin biosynthesis of Ginkgo biloba L. Planta 2005, 222, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Wianowska, D.; Gil, M. Recent advances in extraction and analysis procedures of natural chlorogenic acids. Phytochem. Rev. 2019, 18, 273–302. [Google Scholar] [CrossRef]
- Grace, S.C.; Logan, B.A. Energy dissipation and radical scavenging by the plant phenylpropanoid pathway. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2000, 355, 1499–1510. [Google Scholar] [CrossRef]
- Moghaddam, E.; Teoh, B.-T.; Sam, S.-S.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a metabolite of baicalein with antiviral activity against dengue virus. Sci. Rep. 2014, 4, 5452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alenazi, M.; Egamberdieva, D.; Ahmad, P. Arbuscular mycorrhizal fungi mitigates NaCl induced adverse effects on Solanum lycopersicum L. Pak. J. Bot 2015, 47, 327–340. [Google Scholar]
- Woodward, A.J.; Bennett, I.J. The effect of salt stress and abscisic acid on proline production, chlorophyll content and growth of in vitro propagated shoots of Eucalyptus camaldulensis. Plant Cell Tissue Organ Cult. 2005, 82, 189–200. [Google Scholar] [CrossRef]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M.; Khan, M.M.A. Calcium chloride and gibberellic acid protect linseed (Linum usitatissimum L.) from NaCl stress by inducing antioxidative defence system and osmoprotectant accumulation. Acta Physiol. Plant. 2010, 32, 121–132. [Google Scholar] [CrossRef]
- Herrera-Medina, M.J.; Steinkellner, S.; Vierheilig, H.; Ocampo Bote, J.A.; García Garrido, J.M. Abscisic acid determines arbuscule development and functionality in the tomato arbuscular mycorrhiza. New Phytol. 2007, 175, 554–564. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Jung, H.-Y.; Lee, J.-H.; Lee, I.-J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef]
- Younis, M.E.; Hasaneen, M.N.; Kazamel, A.M. Exogenously applied ascorbic acid ameliorates detrimental effects of NaCl and mannitol stress in Vicia faba seedlings. Protoplasma 2010, 239, 39–48. [Google Scholar] [CrossRef]
- Özdemir, F.; Bor, M.; Demiral, T.; Türkan, İ. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul. 2004, 42, 203–211. [Google Scholar] [CrossRef]
- Li, J.-T.; Qiu, Z.-B.; Zhang, X.-W.; Wang, L.-S. Exogenous hydrogen peroxide can enhance tolerance of wheat seedlings to salt stress. Acta Physiol. Plant. 2011, 33, 835–842. [Google Scholar] [CrossRef]
- Ma, L.; Li, Y.; Yu, C.; Wang, Y.; Li, X.; Li, N.; Chen, Q.; Bu, N. Alleviation of exogenous oligochitosan on wheat seedlings growth under salt stress. Protoplasma 2012, 249, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E. Multifunctional fungal plant symbionts: New tools to enhance plant growth and productivity. New Phytol. 2011, 189, 647–649. [Google Scholar] [CrossRef]
- Gusain, Y.S.; Singh, U.; Sharma, A. Enhance activity of stress related enzymes in rice (Oryza sativa L.) induced by plant growth promoting fungi under drought stress. Afr. J. Agric. Res. 2014, 9, 1430–1434. [Google Scholar]
- Ali, S.; Khan, A.L.; Ali, L.; Rizvi, T.S.; Khan, S.A.; Hussain, J.; Al-Harrasi, A. Enzyme inhibitory metabolites from endophytic Penicillium citrinum isolated from Boswellia sacra. Arch. Microbiol. 2017, 199, 691–700. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Fresh Biomass | Fungal Dry Biomass | IAA (µg/mL) | Phenols (µg/mL) | Flavonoids (µg/mL) | |
|---|---|---|---|---|---|
| Control | 3.97 ± 0.2 b | 1.9 ± 0.2 c | 21 ± 1.2 c | 50 ± 2.8 c | 40 ± 2.3 b |
| 100 mM NaCl | 3.7 ± 0.2 b | 1.8 ± 0.1 c | 35.8 ± 1.3 d | 70 ± 3.2 d | 50 ± 2.5 c |
| 150 mM NaCl | 1.1 ± 0.1 a | 0.6 ± 0.1 b | 8 ± 0.5 b | 20 ± 1.1 b | 11 ± 0.6 a |
| 200 mM NaCl | 1 ± 0.1 a | 0.1 ± 0.1 a | 2 ± 0.1 a | 9 ± 0.5 a | 7 ± 0.4 a |
| NO | t R (min) | Proposed Formula | Mode | Precursor Ion, m/z | LC-ES1-MS/MS Ions | Identification | References |
|---|---|---|---|---|---|---|---|
| 1 | 1.66 | C7H6O3 | _ | [M-H] − 137 | 106.92, 92.92 | Salicylic acid | Bduhafsdun et al., 2018 [36] |
| 2 | 3.39 | C15H10O5 | _ | [M-H] − 269 | 267, 251, 225, 223, 209, 197, 195, 181, 167, 154 | Baicalein | Soraia et al., 2009 [37] |
| 3 | 2.58 | C6H6O6 | _ | [M-H] − 173 | 115.92, 128.83 | Aconitic acid | Soraia et al., 2009 [38] |
| 4 | 7.83 | C17H20O9 | _ | [M-H] − 367 | 177.08 | Feruloylquinic acid | Ghareeb et al., 2018 [39] |
| 5 | 7.79 | C16H20O8 | _ | [M-H] − 339 | 163.00, 132.92 | Coniferyl aldehyde hexoside | Terraza et al., 2016 [40] |
| 6 | 5.53 | C18H26O10 | _ | [M-H] − 401 | 383, 365, 357, 344, 321, 284, 260, 241, 213, 197, 176, 144 | Benzyl alcohol hexose pentose | Bystroma et al., 2008 [41] |
| 7 | 8.26 | C26H29O15 | _ | [M-H] − 581 | 501.42 | Pentose | Beelders et al., 2014 [42] |
| 8 | 7.33 | C16H18O9 | _ | [M-H] − 353 | 177.00, 163.00 | Chlorogenic acid | Koolen et al., 2013 [43] |
| 9 | 4.55 | C15H10O8 | + | [M+H] + 318 | 300.33, 256.25 | Myricetin | Bonta, 2017 [44] |
| 10 | 4.71 | C22H29NO2 | + | [M+H] + 340 | 322, 296, 215, 284, 312 | Propoxyphene | Cao et al., 2015 [45] |
| 11 | 3.85 | C16H14FN3O | + | [M+H] + 284 | 228.17, 198.08, 184.00, 170.00, 157.00, 143.92, 129.92 | Aminoflunitrazepam | Cao et al., 2015 [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jan, F.G.; Hamayun, M.; Hussain, A.; Jan, G.; Ali, S.; Khan, S.A.; Lee, I.-J. Endophytic Candida membranifaciens from Euphorbia milii L. Alleviate Salt Stress Damages in Maize. Agronomy 2022, 12, 2263. https://doi.org/10.3390/agronomy12102263
Jan FG, Hamayun M, Hussain A, Jan G, Ali S, Khan SA, Lee I-J. Endophytic Candida membranifaciens from Euphorbia milii L. Alleviate Salt Stress Damages in Maize. Agronomy. 2022; 12(10):2263. https://doi.org/10.3390/agronomy12102263
Chicago/Turabian StyleJan, Farzana Gul, Muhammad Hamayun, Anwar Hussain, Gul Jan, Sajid Ali, Sumera Afzal Khan, and In-Jung Lee. 2022. "Endophytic Candida membranifaciens from Euphorbia milii L. Alleviate Salt Stress Damages in Maize" Agronomy 12, no. 10: 2263. https://doi.org/10.3390/agronomy12102263