1. Introduction
With the greenhouse effect, more frequent and intense high temperatures have a very adverse impact on the survival of plants. Plants with different morphological structures and physiological characteristics have different tolerance ranges to heat stress (HS). When the ambient temperature exceeds its tolerance range, the extreme temperature will adversely affect the cells and inhibit their normal physiological functions [
1]. When the temperature exceeds the tolerance range, plants will inhibit reproductive development, invest more energy in resisting external pressure, seeds will stop germination, flowers and fruits will delay development, and even their vegetative growth will be inhibited [
1]. In order to cope with high temperatures, plants will change their own morphological structure and internal physiological and biochemical processes: reduce the light-receiving area by curling leaves, close stomata to reduce water loss, increase intracellular reactive oxygen species (ROS) to activate downstream regulatory pathways, imbalance ion channels on the membrane, and increase various heat shock proteins to perform functions.
When plants are exposed to high temperatures for a long time, they will change their metabolism to adapt to the environment and have stronger resistance and adaptation to high temperature. This adaptation is called heat tolerance. Heat tolerance involves a variety of regulatory pathways in organisms: a series of complex physiological and biochemical processes, such as the reduction in membrane lipid molecular fluidity, the production and removal of reactive oxygen species, and the regulation of heat shock protein production [
2,
3,
4].
In order to reduce the damage of increasingly severe high-temperature environments to plants, it is crucial to further explore the transmission of heat stress signals in plants.
2. Adaptation of Plants to High Temperatures in Different Climate Types
There are many plants living on the earth. Because of different climatic conditions and growth environments, plants have formed different living habits, and it can be said that the diversity of plants is caused by climate. Therefore, this paper will classify the heat tolerance of plants by climate environment.
2.1. Desert Plants
Ecosystems in arid areas suffer from intense evaporation, strong winds and sand, strong sunlight and high temperatures, and depleted and salinized soils, creating an extremely harsh environment. It is difficult to grow plants in this area, or the growing conditions are extremely harsh. In the process of plant evolution, plant leaves are more sensitive to environmental changes and have stronger plasticity. Their structural and functional characteristics show the corresponding countermeasures of plants to the environment and the ability of plants to use resources [
5]. In order to adapt to the environment of extreme drought and high salinity, some leaves of desert plants shrink and even evolve into thin scales or hard spines. Some leaves are fleshy and the shape is mostly an irregular cylinder or cone. Some leaves have strong splitting, white fuzz, deep stomata, and so on to reduce the loss of water [
6]. Some of the leaves have been degraded, using the shoots of the previous year for photosynthesis.
As a nutrient transport bridge between plant roots and leaves, stems have developed unique structural characteristics in order to adapt to extreme environments. Encapsulated by the pericarp to inhibit transpiration in plants, they have developed transport tissue to transport water. It is composed of a large number of parenchyma cells, which improves the water retention of cells [
7]. In arid regions, most plants have developed roots in order to absorb water from deeper soil. However, in some desert areas with more precipitation, the root system will have great changes: the taproot will be inhibited and the lateral root will develop. This is because shallow soil can carry more moisture [
8]. Arid regions have less water and higher temperatures, which are accompanied by more saline soils. Plants that live in the region will have degraded leaves that cone, reducing water loss from transpiration, and having more developed roots for water uptake [
9]. Its molecular regulation is also more inclined to the mechanism of salt and heat tolerance; for example, the cell contains more sodium ions and potassium ions.
In addition to the evolution of special forms, there are also very complex regulatory mechanisms at the molecular level for abiotic stress in plants. These responses are controlled by the expression of only a few genes, and the products of some genes are not only stress tolerance effector proteins but also signal transduction factors and transcription factors in stress responses: AREB, sHSP, NAC, WRKY, MYB, ERF [
10]. These gene families have been proven to regulate drought, heat, and salt stress in different ways. They form vast and complex networks meticulously calculating the best way to survive [
11].
2.2. Tropical Plants
The temperature of the appropriate interval affects the growth and development of plants, but extreme temperatures may endanger plant life. However, some plants can raise the tolerance range of temperatures. In the tropics, even during the rainy season, temperatures exceed 40 °C. Tropical plants have also evolved more comprehensively to withstand heat.
To adapt to high temperatures, plants have evolved a number of different ways and channels. Changes in plant tissue morphology are used to better adapt to high temperatures. The flat shape of the leaf is designed to absorb light energy efficiently, but enough light is often accompanied by deadly heat, so leaves have evolved fur and scales to filter sunlight. Other plants also reflect sunlight through their leathery leaves, which can reduce the excess energy that comes with it and protect the plant body from heat damage. Some plant leaves are arranged vertically so that the leaf edge is towards light or the leaves are folded under high-temperature conditions, reducing the area of light absorption [
2]. The physiological adaptation of plants to high temperatures is mainly to reduce the water content of cells and increase the concentration of soluble sugar or salt, which is conducive to slowing down the metabolic rate and increasing the anticoagulation ability of protoplasm. The second is to rely on vigorous transpiration to avoid plant body damage due to overheating [
3].
Different plants have different defense mechanisms against high-temperature stress, which vary with stress time and intensity [
4]. Some specific pathways are widely present in plants, such as the ROS regulatory system. Trace amounts of ROS act as signal molecules to promote downstream defense responses, whereas excessive ROS can aggravate membrane lipid oxidation, cause fatty acid decomposition, increase intracellular malondialdehyde (MDA) levels, and destroy normal physiological functions of cells [
12,
13]. Hormones also widely regulate plant stress defense responses and ethylene signal transduction pathways cross with heat stress responses through APETAL2/ethylene-responsive element binding factors (AP2/ERF) transcription factors to generate heat shock proteins (HSPs). Melatonin, as a signal molecule, can scavenge intracellular free radicals (ROS/reactive nitrogen species (RNS)) [
14] and heat shock protein-dependent pathways altered in plants, thereby enhancing plant abiotic stress resistance [
15]. In addition, under high-temperature environments, the content of various compounds such as proline and brassinolide in plants increases, which regulates osmotic stress and improves plant heat tolerance [
16]. For example, brassinolide regulates the activity of transcription factor BES1, promotes the production of heat shock proteins, and helps plants to rebuild homeostasis [
17,
18].
3. Changes in Cell Membrane Permeability
The plant heat stress signaling pathway is a complex collection of cellular signaling pathways and molecular mechanisms, and the mutual regulation of these signaling pathways can help plants adapt to and resist high-temperature environments to ensure their normal physiological functions. High-temperature environments affect the stability of proteins, membranes, and cytoskeletons in plant cells to varying degrees. They also affect the activity of enzymes to change the efficiency of enzyme catalysis and further cause metabolic disorder in plants.
The integrity and function of biomembranes are very sensitive to high temperatures and can feel a slight change in temperature. Prolonged exposure to high temperatures will change the tertiary and quaternary structures of protein, accelerate the movement of phospholipid molecules on the cell membrane, increase the concentration of unsaturated fat acid, and enhance fluidity.
4. Conduction of Calcium Signal
When plants are subjected to high temperatures, sensitive calcium channels on the cell membrane are activated and there is an extracellular Ca
2+ influx. Calcium ion influx can activate various downstream signaling pathways, completing the first step of plant primary heat tolerance response. However, in reality, calcium ion channels cannot directly sense temperature changes, but rather the changes in plasma membrane fluidity and permeability caused by high temperatures induce channel opening. In higher plants, experiments using Ca
2+-chelating agent-lanthanum chloride anhydrous (LaCl
3) have shown that calcium channels have also been validated as one of the main thermal sensors [
1,
2,
4].
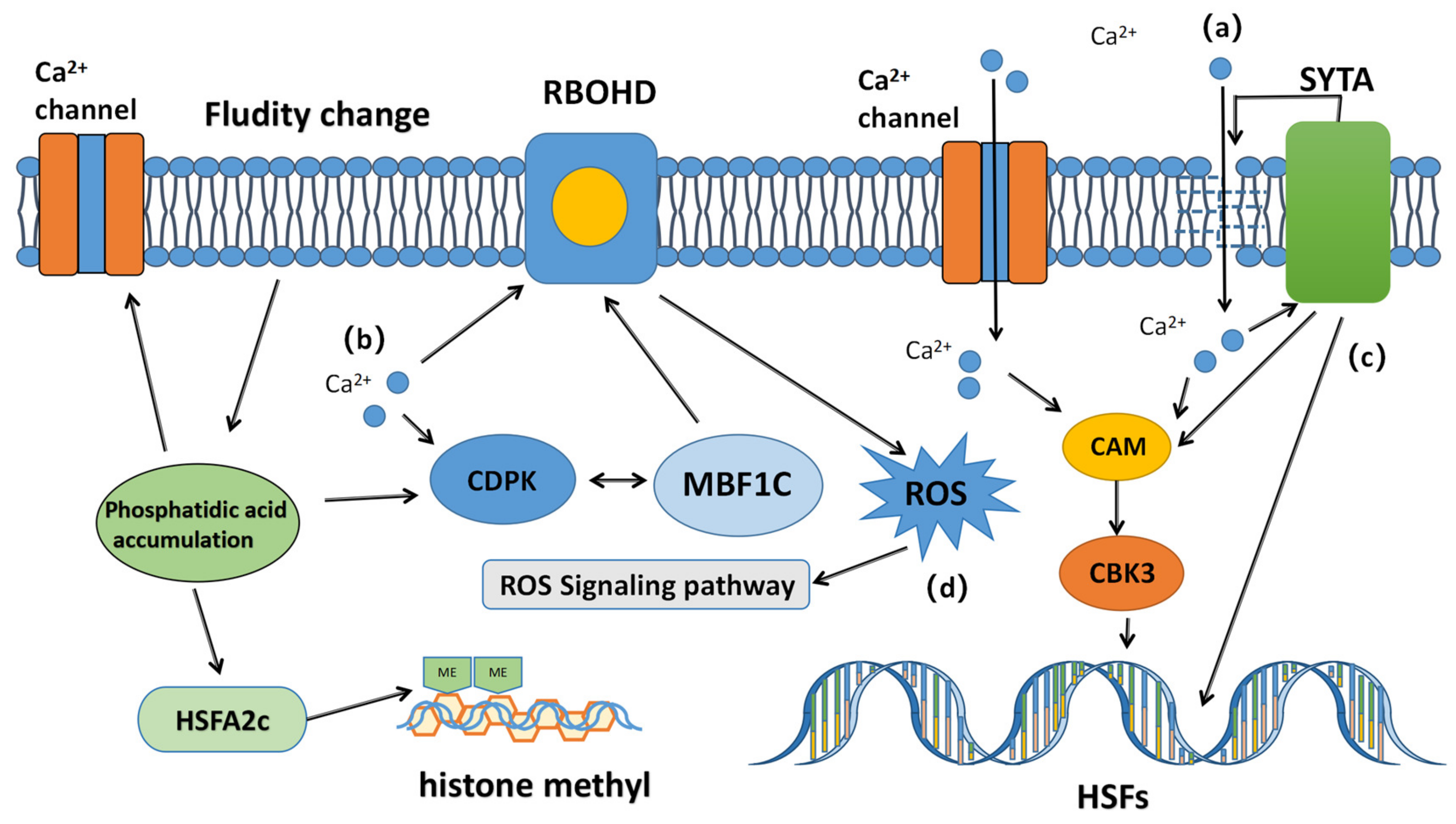
Free Ca
2+ is a second messenger commonly present in animals and plants, participating in various physiological reactions. When the cell sensor captures changes in the external environment, opens the calcium channel protein, and the intracellular Ca concentration increases, the calcium sensor transducer signals are transmitted to different substrates through the calcium sensor to different receivers, thereby activating different regulatory pathways (
Figure 1) [
19].
First, intracellular calcium activates calmodulin 3 (CaM3) and calcium-dependent protein kinases (CDPKs). After the combination of calcium ions and calmodulin, the released signal molecules start calmodulin-binding kinase (CBK3). Ca
2+ binds to CaM3 and activates and phosphorylates heat shock factors (HSFs). Combined with the heat shock element in the promoter region of the
HSP gene, the transcriptional regulation of HSP production was activated and involved in the regulation of HSP production. Multiprotein-bridging factor 1c (MBF1c), as a co-activator of CDPK, stimulates respiratory burst oxidase homologous protein D (RBOHD) to produce (ROS) and regulates the expression of downstream antioxidant enzymes [
20]. Next, the activated CDPK activates mitogen-activated protein kinases (MAPKs) and RBOHD [
21], phosphorylates HSFA2, and other transcription factors, regulates the expression of HSPs, and then antagonizes HS [
22].
Calcium ions also activate many other pathways to enhance high-temperature resistance. Synaptotagmin A (SYTA) is a transmembrane protein that is driven by Ca
2+ to repair damaged cell membranes and regulate CaM3 expression. Intracellular calcium concentration also affects the production of specific heat shock molecules (HSFA2C) (
Figure 1) [
23]. At high temperatures, phospholipids are hydrolyzed into phosphatidic acid, which acts as an important second messenger in signal transduction, enhances the expression of HSFA2C, and then regulates the expression of HSP genes. In wheat, calmodulin is activated by calcium ions, which are involved in regulating the expression of heat shock proteins and establishing stress defense [
23]. At the same time, phospholipid-signaling molecules can also further regulate the flow of calcium ions.
5. Production of Reactive Oxygen Species (ROS) under Heat Stress
In the early stage of HS, the content of reactive oxygen species in the cells gradually increased, and the expression of HSFs and the accumulation of HSPs in this stage played a decisive role. Low levels of ROS act as signaling molecules to promote downstream HS reactions, but excessive ROS accumulation produces toxicity, and the detoxification mechanism of ROS protects plants from HT stress [
24].
The slight change in temperature will significantly affect the photosynthesis of plants [
1,
25]. The high temperature of the environment will inhibit the activity of photosystem II, photosystem I, and rubisco enzymes on the thylakoid membrane of plants, thus affecting their mediated light and electron transport chains, leading to the disorder of redox and metabolism, and a large amount of light energy is left to produce more reactive oxygen species (ROS) [
26]. Under a stress environment, ROS often play a role in the establishment of plant defense. Low levels of ROS are involved in signal transduction to promote downstream responses and high levels of ROS cause membrane peroxidation [
23], causing oxidative damage to cells, resulting in metabolic imbalance, protein denaturation, and deformation, which results in protein toxicity stress, destruction of membrane stability and cytoskeleton integrity, and leads to cell structure collapse and even programmed death [
27].
Therefore, the signal network of ROS needs to be highly conserved in plants to control a series of physiological metabolic reactions. In previous studies, the focus was on the toxicity of ROS to cells. However, recent studies have focused on its direction as a signal molecule. This requires the plant to maintain a dynamic balance of ROS so that it exercises the role of signal molecules without poisoning the plant.
Under abiotic stress, plants produce ROS signals and induce downstream reactions. One of them is the activation of defense genes, resulting in specific stress responses. For example, Arabidopsis upregulates the expression of antioxidant genes and improves the antioxidant capacity of plants in response to ROS signal transduction. However, the mechanisms involved in ROS induction remain unclear.
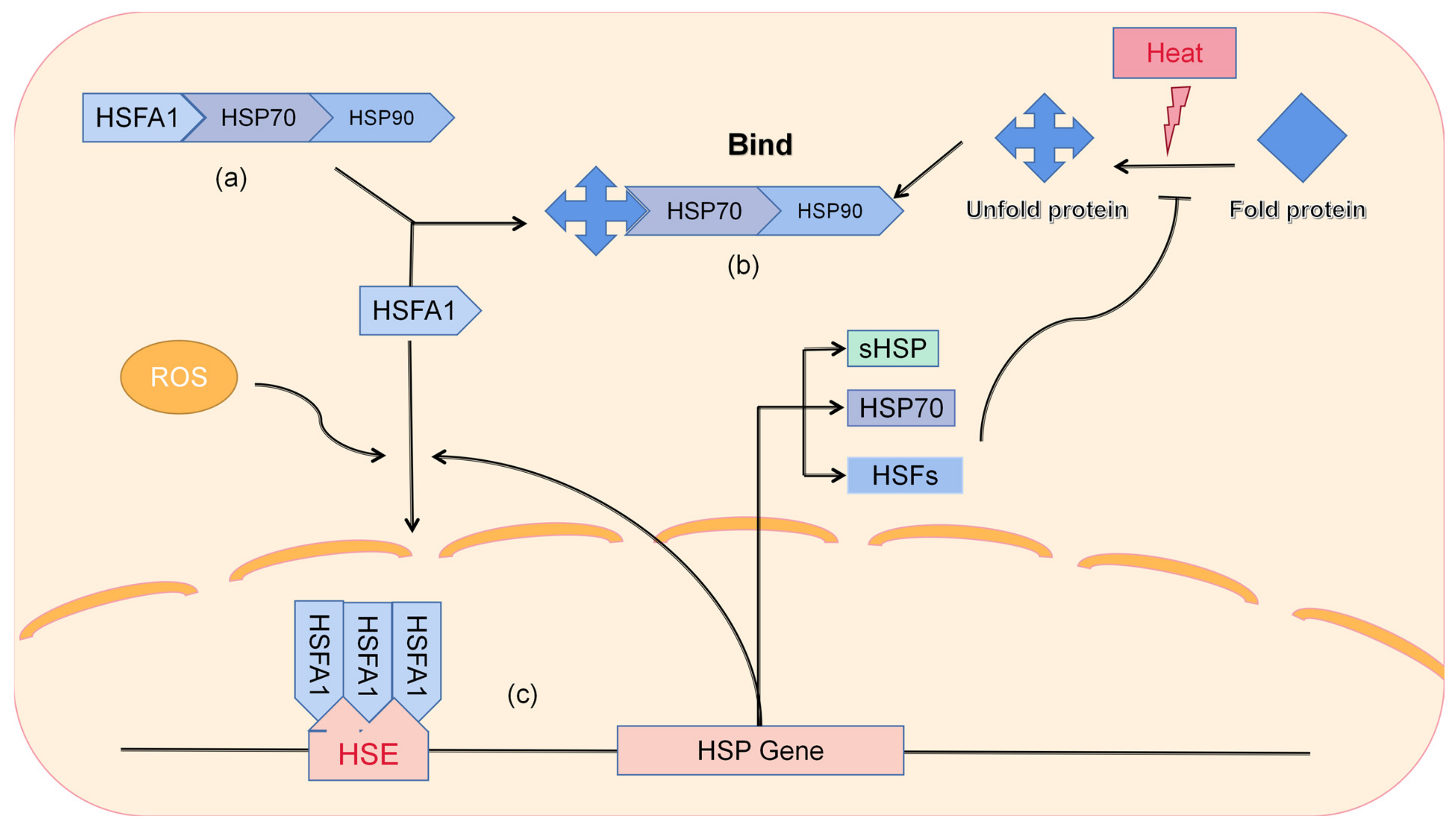
6. Production and Function of Heat Stress Protein
Heat shock proteins are the main participants in plant heat stress. Usually, heat stress genes (HSGs) are upregulated by environmental high-temperature induction. Further encoding HSPs, protecting intracellular proteins from denaturation, and maintaining their stability and function through protein folding thereby helps plants rebuild homeostasis and resist high temperatures [
28] (
Figure 2).
In plants, heat stress proteins are generally divided into five families based on their molecular weight: HSP100, HSP90, HSP70, HSP60, and small molecular weight HSP20 (sHSPs) [
29]. HSP can not only solve the problem of protein misfolding to some extent in cells but also protect the stability of other proteins as molecular chaperones.
The function of the protein is related to its conception, and the function of the protein with the same three-dimensional structure is also consistent [
24]. When the three-dimensional structure is destroyed, the protein will lose its original function. Timperio pointed out that HSP, as a molecular-binding protein, can bind to the substrate protein to maintain its stability and molecular function; on the other hand, it can reduce the expression of other proteins to a certain extent [
30]. QU (2013) also shows how to perform the functions of heat shock proteins. As the dominant factor, HSFA1 triggers the heat stress effect by producing co-activators HSFA1b and HSFA2, forming a homotrimer that binds to the heat shock promoter element and induces the expression of various HSPs, HSF70, HSF101, and sHSP [
25].
The regulatory expression of HSP is largely controlled by HSFs [
31]. An HSF does not normally participate in heat tolerance (
Figure 2). After changes in environmental temperature, HSFs are activated by various signaling molecules through various pathways. In the whole HSF gene family,
HSFA1 is responsible for the main regulatory function. Studies have shown that if this gene is inhibited in tomatoes, the plants will not express HSPs or the expression level will be very small, affecting the normal growth and development of the plants and showing more serious symptoms of heat damage [
32]. The HSF family, with HSFA1 as the main regulator, presents a complex network structure and co-regulates the expression of HSP [
33,
34]. Despite extensive research on HSFs, no direct upstream transcription factors with a role in thermal signal transduction have been found. Further research is needed on both HSFs and other novel transcription factors.
7. Heat-Resistant Mechanism of Unfolded Protein
Unfolded protein response (UPR) is a signal pathway activated in cells, which responds to the stress of damaged protein stability in the endoplasmic reticulum (ER) [
33,
35]. Secreted proteins and membrane proteins are folded and modified in the endoplasmic reticulum. Under high-temperature conditions, a large number of secreted proteins gather in the endoplasmic reticulum waiting for modification. When the folding capacity of the system is exceeded, a large number of unfolded proteins will accumulate, and the normal biological function of the endoplasmic reticulum will be blocked [
36]. In plants, two UPR pathways have been reported, one in the ER and the other in the cytoplasm [
37,
38]. When the organism is subjected to stress disasters such as high temperature, high salt, and drought, the protein that needs to be folded exceeds the capacity of the system to fold, and the UPR response is activated [
39]. In the process of endoplasmic reticulum folding, the regulation of cofactors is required, and the basic leucine zipper (bZIP) and transcription factors can help other proteins prevent misfolding. Under external stress, bZIP and transcription factors in the endoplasmic reticulum are released [
40]. The released transcription factors enter the nucleus, activating the transcription of specific genes within the nucleus, leading to the accumulation of ER partner transcripts and the activation of brassinosteroid signaling [
40]. In
Arabidopsis thaliana, bZIP28 and bZIP60 are upregulated by high-temperature stress and play a role in the regulatory pathway of UPR, whereas the decrease in their expression levels will show serious high-temperature damage and reduce plant resistance [
40].
Different from the UPR in the ER, the UPR in the cytoplasm is triggered by the unfolded protein in the cytoplasm, which is mainly regulated by HSF and HSFA2. HSFA2 binds to the HSF binding element in the heat shock response (HSR) gene promoter [
40]. However, in the early stage of heat stress temperature rise, secreted proteins do not accumulate, and UPR response is slow, unlike calcium ion channels. However, heat stress proteins can accumulate in plant cells at warmer temperatures [
41], and the activation of UPR seems to require plasmalemma to emit specific calcium signals [
42], indicating that UPR acts as a downstream heat stress signal transduction rather than a heat receptor.
8. Carbohydrates Can Be Used as Heat Conduction Resistance Signals
In plants, as an organism’s source of carbon and energy, carbohydrates carry out important biological functions. According to research, glucose metabolism can also act as signaling molecules to regulate other physiological processes [
42]. Intracellular accumulation of soluble sugars can regulate osmotic pressure, reduce abnormal ion loss, and maintain membrane stability to ensure physiological function [
43,
44]. Therefore, changes in glucose metabolism caused by HS can have adverse effects on sugar-sensitive systems.
In the whole development process of plants, the reproductive stage is extremely sensitive to high temperature, and even a small temperature rise will affect the development of pollen tubes [
45].
At high temperatures, the resistance mechanism of plants is activated, the rate of sugar metabolism is accelerated to provide energy for the body, and the expression of glycolysis- and citric acid cycle-related proteins are increased [
46]. Glucose is the most common sugar with a stable molecular structure. When the ambient temperature rises, glucose will reduce the peroxidation concentration of membrane lipids and then protect the process of photosynthesis to cope with heat stress [
47]. At the same time, it was found that MAPKs in the heat-resistant signal transduction pathway were activated after being treated with Aspergillus pinosus, whereas those treated with sucrose did not respond, indicating that plants may have different signal transduction pathways under heat stress [
48].
9. Relationship between Proline and Heat Stress
Proline is involved in the regulation of heat stress under heat stress. Many studies have shown that proline performs a variety of functions, helping to regulate osmotic pressure, remove reactive oxygen species, and stabilize cell structure [
49]. Early studies have shown that the accumulation ability of proline is related to stress tolerance [
50,
51]. Proline is a kind of amino acid that plays an important role under stress conditions, serving as an osmotic pressure solute and signal molecule in plants [
52]. In broad beans, the biosynthesis of proline and the involvement of nitric oxide and proline in the heat tolerance mechanism have been reported [
52]. Proline can also effectively remove hydroxyl free radicals in cells and reduce or eliminate oxidative stress caused by reactive oxygen species under heat stress [
53]. Many studies have shown that proline accumulation is positively related to the stress degree of plants, and the higher the plant stress resistance, the more proline accumulation in cells; however, some people believe that the increase in proline concentration under stress is a result of coercion and is not a response to stress [
51,
52,
54,
55]. Proline, as a molecular chaperone, regulates the integrity of proteins and enhances the activities of different enzymes. Its functions include preventing protein aggregation, clearing hydroxyl-reactive oxygen species, and regulating intracellular osmotic potential [
56]. Many studies attribute antioxidant properties to proline, certificating its ability to scavenge reactive oxygen species and its role as a singlet oxygen quencher [
57].
10. miRNAs Involved in the Construction of Plant Heat Tolerance Networks
miRNA is a small endogenous non-coding single-stranded RNA molecule that is involved in the heat-resistant defense construction of plants [
1]. miRNA can be used as a signal molecule to regulate the corresponding genes of plant heat tolerance and affect plant heat tolerance. In response to heat stress, upregulated miRNAs may downregulate their specific genes, where miRNAs act as negative regulators of heat tolerance. Conversely, downregulated miRNAs may upregulate their target genes and affect heat tolerance [
58].
In addition, miRNAs also play a major role in plant growth and development [
59], hormone response [
13], and stress defense [
60]. A series of miRNAs that regulate heat stress responses have been discovered in different plants such as Arabidopsis [
61]. In poplar, 12 miRNA families were identified in response to high-temperature stress. Most of their target genes were drought stress response factors or signal transduction proteins and different families had different induction patterns [
62]. Under heat stress conditions, miRNA regulates a large number of stress-responsive genes, which are involved in various biological pathways for plants to acquire heat tolerance defense mechanisms [
63].
11. Conclusions
Plant high-temperature stress is a very complex process, from morphological construction to the regulation of plant biochemical molecules, which are involved in the resistance to adverse environments. High temperatures will first accelerate the evaporation of water so that plants are in a state of drought and water shortage, reduce the generation and transfer of active ingredients in the body, and at the same time, the activity of enzymes in cells is reduced under stress, disrupting normal metabolism. In previous studies, high temperature affects the activity of all enzymes and disrupts the normal metabolism of plants. There has been a more reasonable explanation of the adaptive mechanism of HS, but there are still some gaps. The plasma membrane can effectively capture changes in temperature, but its mechanism is not fully interpretable, and there are still many questions to be answered. For example, how do cell membranes conduct cascade conduction after sensing heat, and how does the mechanism of action of phosphate groups and calcium channels, MAPKs, act on HSF?
The heat stress signaling cascade is interconnected at different levels, activating all signaling pathways. The complex regulatory network can sensitively perceive the current stress state and provide the most appropriate coping strategies. Different related metabolites are also appropriately accumulated in order to better maintain the normal biological function of the plant. Under the current conditions of climate change and population explosion, it is necessary to develop heat-resistant varieties to meet the world’s food demand. Therefore, it is very necessary to explore and develop the heat-resistant mechanism of plants.
Author Contributions
Conceptualization, Z.C. and E.W.; methodology, software, and analysis by X.X. (Xinyi Xu), C.T. and X.Z.; formal analysis validation and investigation by Z.C. and E.W.; writing by E.W., Z.C. and Y.Z., J.W., X.S., X.X. (Xinli Xia), H.-l.W. analyzed the data.; Z.C. and E.W. visualized the data and drafted the manuscript that was critically revised by X.X. (Xinli Xia) and Y.Z.; Y.Z. had the overall responsibility of experimental design and manuscript preparation. All authors have read and agreed to the published version of the manuscript.
Funding
This study was jointly supported by Hainan Provincial Natural Science Foundation (320RC471). National Natural Science Foundation of China (32360415, 32160400) and Hainan Provincial Key Research and Development Program of China (ZDYF2022XDNY179).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kaushal, N.; Bhandari, K.; Siddique, K.H.M.; Nayyar, H. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric. 2016, 2, 1134380. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, J.K. Abiotic stress signal transduction in plants: Molecular and genetic perspectives. Physiol. Plant. 2001, 112, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Antunes, S.C.; Freitas, R.; Figueira, E.; Gonçalves, F.; Nunes, B. Biochemical effects of acetaminophen in aquatic species: Edible clams Venerupis decussata and Venerupis philippinarum. Environ. Sci. Pollut. Res. Int. 2013, 20, 6658–6666. [Google Scholar] [CrossRef] [PubMed]
- Driedonks, N.; Rieu, I.; Vriezen, W.H. Breeding for plant heat tolerance at vegetative and reproductive stages. Plant Reprod. 2016, 29, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xu, H.; Wang, Y.; Wang, X.; Qiu, Y.; Xu, B. The effect of salt stress on the chlorophyll level of the main sand-binding plants in the shelterbelt along the Tarim Desert Highway. Chin. Sci. Bull. 2008, 53 (Suppl. 2), 109–111. [Google Scholar] [CrossRef]
- Went, F.W. Ecology of Desert Plants. II. The Effect of Rain and Temperature on Germination and Growth. Ecology 1949, 30, 1–13. [Google Scholar] [CrossRef]
- Kirschner, G.K.; Xiao, T.T.; Blilou, I. Rooting in the Desert: A Developmental Overview on Desert Plants. Genes 2021, 12, 709. [Google Scholar] [CrossRef]
- Li, C.; Han, H.; Ablimiti, M.; Liu, R.; Zhang, H.; Fan, J. Morphological and physiological responses of desert plants to drought stress in a man-made landscape of the Taklimakan desert shelter belt. Ecol. Indic. 2022, 140, 109037. [Google Scholar] [CrossRef]
- Downton, W.J.; Berry, J.A.; Seemann, J.R. Tolerance of Photosynthesis to High Temperature in Desert Plants. Plant Physiol. 1984, 74, 786–790. [Google Scholar] [CrossRef]
- Ma, J.; Lu, J.; Xu, J.; Duan, B.; He, X.; Liu, J. Genome-wide Identification of WRKY Genes in the Desert Poplar Populus euphratica and Adaptive Evolution of the Genes in Response to Salt Stress. Evol. Bioinform. 2015, 11 (Suppl. 1), 47–55. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Ahmad, M.; Waraich, E.A.; Skalicky, M.; Hussain, S.; Zulfiqar, U.; Anjum, M.Z.; Habib ur Rahman, M.; Brestic, M.; Ratnasekera, D.; Lamilla-Tamayo, L.; et al. Adaptation strategies to improve the resistance of oil seed crops to heat stress under a changing climate: An overview. Front. Plant Sci. 2021, 12, 767150. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Bhattacharjee, S. Differential competence of redox-regulatory mechanism under extremes of temperature determines growth performances and cross tolerance in two indica rice cultivars. J. Plant Physiol. 2015, 176, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Bahuguna, R.N.; Jha, J.; Pal, M.; Shah, D.; Lawas, L.M.; Khetarpal, S.; Jagadish, K.S. Physiological and biochemical characterization of NERICA-L-44: A novel source of heat tolerance at the vegetative and reproductive stages in rice. Physiol. Plant. 2015, 154, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Khan, M.A.; Shahzad, R.; Bilal, S.; Khan, M.; Yun, B.-W.; Khan, A.L.; Lee, I.-J. Melatonin ameliorates thermotolerance in soybean seedling through balancing redox homeostasis and modulating antioxidant defense, phytohormones and polyamines biosynthesis. Molecules 2021, 26, 5116. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Katano, K. Coordination between ROS regulatory systems and other pathways under heat stress and pathogen attack. Front. Plant Sci. 2018, 9, 490. [Google Scholar] [CrossRef]
- Braam, J. Regulated expression of the calmodulin-related TCH genes in cultured Arabidopsis cells: Induction by calcium and heat shock. Proc. Natl. Acad. Sci. USA 1992, 89, 3213–3216. [Google Scholar] [CrossRef]
- Larkindale, J.; Knight, M.R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiol. 2002, 128, 682–695. [Google Scholar] [CrossRef]
- Suri, S.S.; Dhindsa, R.S. A heat-activated MAP kinase (HAMK) as a mediator of heat shock response in tobacco cells. Plant Cell Environ. 2008, 31, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.-P. Calcium Signaling in Plant Biotic Interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, B. Lipid-and calcium-signaling regulation of HsfA2c-mediated heat tolerance in tall fescue. Environ. Exp. Bot. 2017, 136, 59–67. [Google Scholar] [CrossRef]
- Pucciariello, C.; Banti, V.; Perata, P. ROS signaling as common element in low oxygen and heat stresses. Plant Physiol. Biochem. 2012, 59, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Qu, A.L.; Ding, Y.F.; Jiang, Q.; Zhu, C. Molecular mechanisms of the plant heat stress response. Biochem. Biophys. Res. Commun. 2013, 432, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Portis, A.R. Temperature dependence of photosynthesis in Arabidopsis plants with modifications in Rubisco activase and membrane fluidity. Plant Cell Physiol. 2005, 46, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- Li, G.; Meng, X.Q.; Cai, J.; Dong, T.T.; Zhu, M.K. Advances in the function of reactive oxygen species in plant abiotic stress response. J. Plant Physiol. 2018, 54, 951–959. [Google Scholar]
- Baniwal, S.K.; Bharti, K.; Chan, K.Y.; Fauth, M.; Ganguli, A.; Kotak, S.; Mishra, S.K.; Nover, L.; Port, M.; Scharf, K.D.; et al. Heat stress response in plants: A complex game with chaperones and more than twenty heat stress transcription factors. J. Biosci. Bioeng. 2004, 29, 471–487. [Google Scholar] [CrossRef]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Med. Genom. 2007, 8, 125. [Google Scholar] [CrossRef]
- Levitt, M.; Gerstein, M.; Huang, E.; Subbiah, S.; Tsai, J. Protein folding: The endgame. Annu. Rev. Biochem. 1997, 66, 549–579. [Google Scholar] [CrossRef]
- Timperio, A.M.; Egid, M.G.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (HSP). J. Proteom. 2008, 71, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Morrow, G.; Tanguay, R.M. Small heat shock protein expression and functions during development. Int. J. Biochem. Cell Biol. 2012, 44, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.D. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Dev. 2002, 16, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Hu, G.; Han, B. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice. Plant Sci. 2009, 176, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Pincus, D.; Chevalier, M.W.; Aragón, T.; van Anken, E.; Vidal, S.E.; El-Samad, H.; Walter, P. BiP binding to the ER-stress sensor Ire1 tunes the homeostatic behavior of the unfolded protein response. PLoS Biol. 2010, 8, e1000415. [Google Scholar] [CrossRef]
- Moreno, A.A.; Orellana, A. The physiological role of the unfolded protein response in plants. Biol. Res. 2011, 44, 75–80. [Google Scholar] [CrossRef]
- Howell, S.H. Endoplasmic Reticulum Stress Responses in Plants Stephen. Rev. Plan Biol. 2013, 64, 477–499. [Google Scholar] [CrossRef]
- Sugio, A.; Dreos, R.; Aparicio, F.; Maule, A.J. The cytosolic protein response as a subcomponent of the wider heat shock response in Arabidopsis. Plant Cell 2009, 21, 642–654. [Google Scholar] [CrossRef]
- Che, P.; Bussell, J.D.; Zhou, W.; Estavillo, G.M.; Pogson, B.J.; Smith, S.M. Signaling from the endoplasmic reticulum activates brassinosteroid signaling and promotes acclimation to stress in Arabidopsis. Sci. Signal. 2010, 3, 69. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Humbert, S.; Liu, J.-X.; Srivastava, R.; Rothstein, S.J.; Howell, S.H. Heat induces the splicing by IRE1 of a mRNA encoding a transcription factor involved in the unfolded protein response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 7247–7252. [Google Scholar] [CrossRef] [PubMed]
- Saidi, Y.; Domini, M.; Choy, F.; Zryd, J.P.; Schwitzguebel, J.P.; Goloubinoff, P. Activation of the heat shock response in plants by chlorophenols: Transgenic Physcomitrella patens as a sensitive biosensor for organic pollutants. Plant Cell Environ. 2007, 30, 753–763. [Google Scholar] [CrossRef]
- Saidi, Y.; Finka, A.; Muriset, M.; Bromberg, Z.; Weiss, Y.G.; Maathuis, F.J.; Goloubinoff, P. The heat shock response in moss plants is regulated by specific calcium-permeable channels in the plasma membrane. Plant Cell 2009, 21, 2829–2843. [Google Scholar] [CrossRef] [PubMed]
- Granot, D.; David-Schwartz, R.; Kelly, G. Hexose kinases and their role in sugar sensing and plant development. Front. Plant Sci. 2013, 4, 44. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lawas, L.M.F.; Malo, R.; Glaubitz, U.; Erban, A.; Mauleon, R.; Heuer, S.; Zuther, E.; Kopka, J.; Hincha, D.K.; et al. Metabolic and transcriptomic signatures of rice floral organs reveal sugar starvation as a factor in reproductive failure under heat and drought stress. Plant Cell Environ. 2015, 38, 2171–2192. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kamiyama, M.; Iwata, T.; Makita, N.; Furukawa, H.; Ikeda, H. Moderate increase ofmean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot. 2006, 97, 731–738. [Google Scholar] [CrossRef]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef]
- Huang, Y.-W.; Nie, Y.-X.; Wan, Y.-Y.; Chen, S.-Y.; Sun, Y.; Wang, X.-J.; Bai, J.-G. Exogenous glucose regulates activities of antioxidant enzyme, soluble acid invertase and neutral invertase and alleviates dehydration stress of cucumber seedlings. Sci. Hortic. 2013, 162, 20–30. [Google Scholar] [CrossRef]
- Chavez-Arias, C.C.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Evaluation of heat stress period duration and the interaction of daytime temperature and cultivar on common bean. Environ. Exp. Bot. 2018, 155, 600–608. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Surabhi, G.K.; Reddy, A.M.; Sudhakar, C. NaCl effects on proline metabolism in two high yielding genotypes of mulberry (Morus alba L.) with contrasting salt tolerance. Plant Sci. 2003, 165, 1245–1251. [Google Scholar]
- Farag, M.A.; Meyer, A.; Ali, S.E.; Salem, M.A.; Giavalisco, P.; Westphal, H.; Wessjohann, L.A. A comparative metabolomics approach detects stress specific responses during coral bleaching in soft corals. J. Proteome Res. 2018, 17, 2060–2071. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.A.; Siddiqui, M.H.; Al-Khaishany, M.Y.; Khan, M.N.; Ali, H.M.; Alakeel, K.A. Nitric oxide-mediated cross-talk of proline and heat shock proteins induce thermo tolerance in Vicia faba L. Environ. Exp. Bot. 2018, 161, 290–302. [Google Scholar] [CrossRef]
- Anaytullah, S.A.; Bose, B. Impact of seed hardening treatment with nitrate salts on nitrogen and anti oxidant defense metabolisms in Triticum aestivum L. under different sowing conditions. Vegetos 2012, 25, 292–299. [Google Scholar]
- Chiang, H.H.; Dandekar, A.M. Regulation of proline accumulation in Arabidopsis thaliana (L.) Heynh during development and in response to desiccation. Plant Cell Environ. 1995, 18, 1280–1290. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Murmu, K.; Murmu, S.; Kundu, C.K.; Bera, P.S. Exogenous proline and glycine betaine in plants under stress tolerance. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 901–913. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Wang, M.; Shah, L.; Lu, S.; Wang, X.; Ma, C. Unraveling field crops sensitivity to heat stress: Mechanisms, approaches, and future prospects. Agronomy 2018, 8, 128. [Google Scholar] [CrossRef]
- Liu, H.; Yu, H.; Tang, G.; Huang, T. Small but powerful: Function of microRNAs in plant development. Plant Cell Rep. 2018, 37, 515–528. [Google Scholar] [CrossRef]
- Curaba, J.; Singh, M.B.; Bhalla, P.L. miRNAs in the crosstalk between phytohormone signaling pathways. J. Exp. Bot. 2014, 65, 1425–1438. [Google Scholar] [CrossRef]
- Wu, J.; Wang, L.; Wang, S. microRNAs associated with drought response in the pulse crop common bean (Phaseolus vulgaris L.). Gene 2017, 10, 78–86. [Google Scholar] [CrossRef]
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).



 ,
, {kind=link}
{kind=link}

