
The Effect of Hot Wind on Needle and Stem Water Status: Response Strategies in Resprouting and Non-Resprouting Pine Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Measurements
2.3. Needle Water Potential and Needle Osmotic Potential
2.4. Needle Water Content
2.5. Stem Water Status
2.6. Sprouting
2.7. Statistical Analyses
3. Results
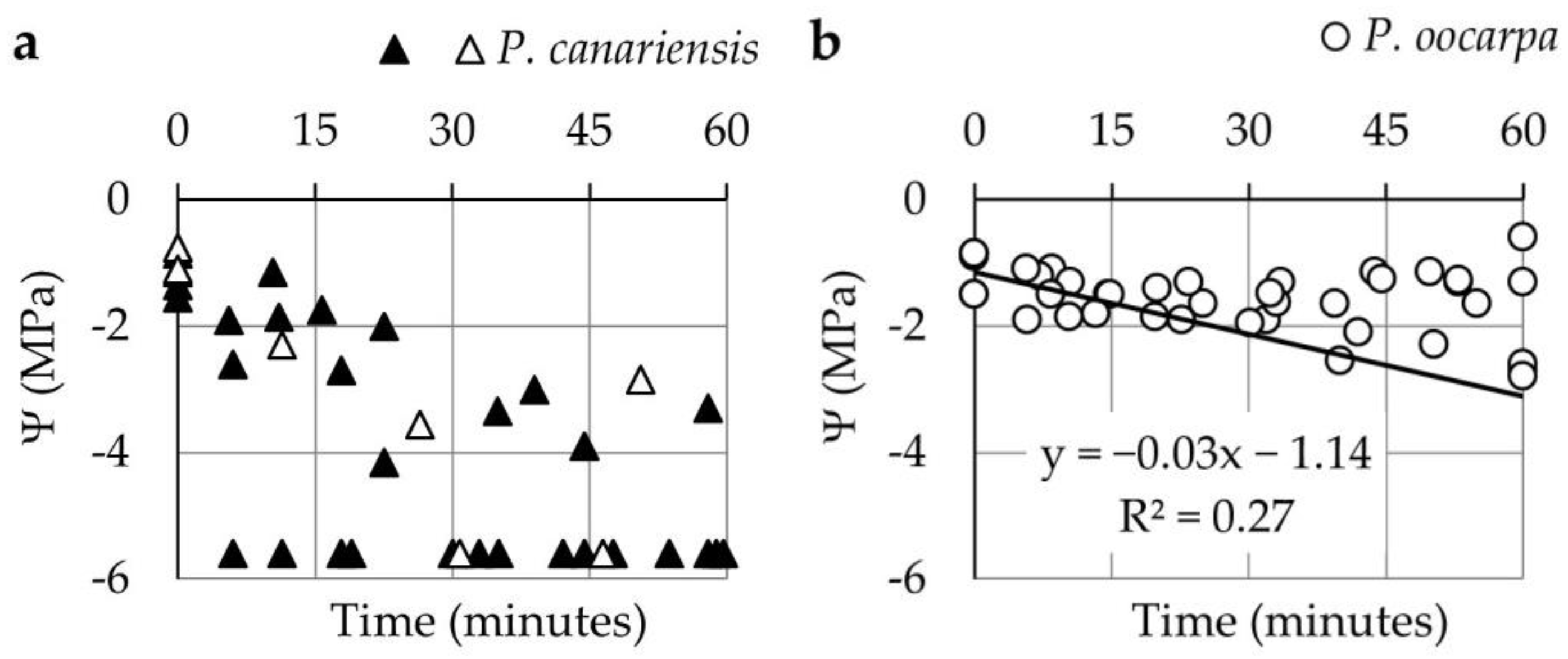
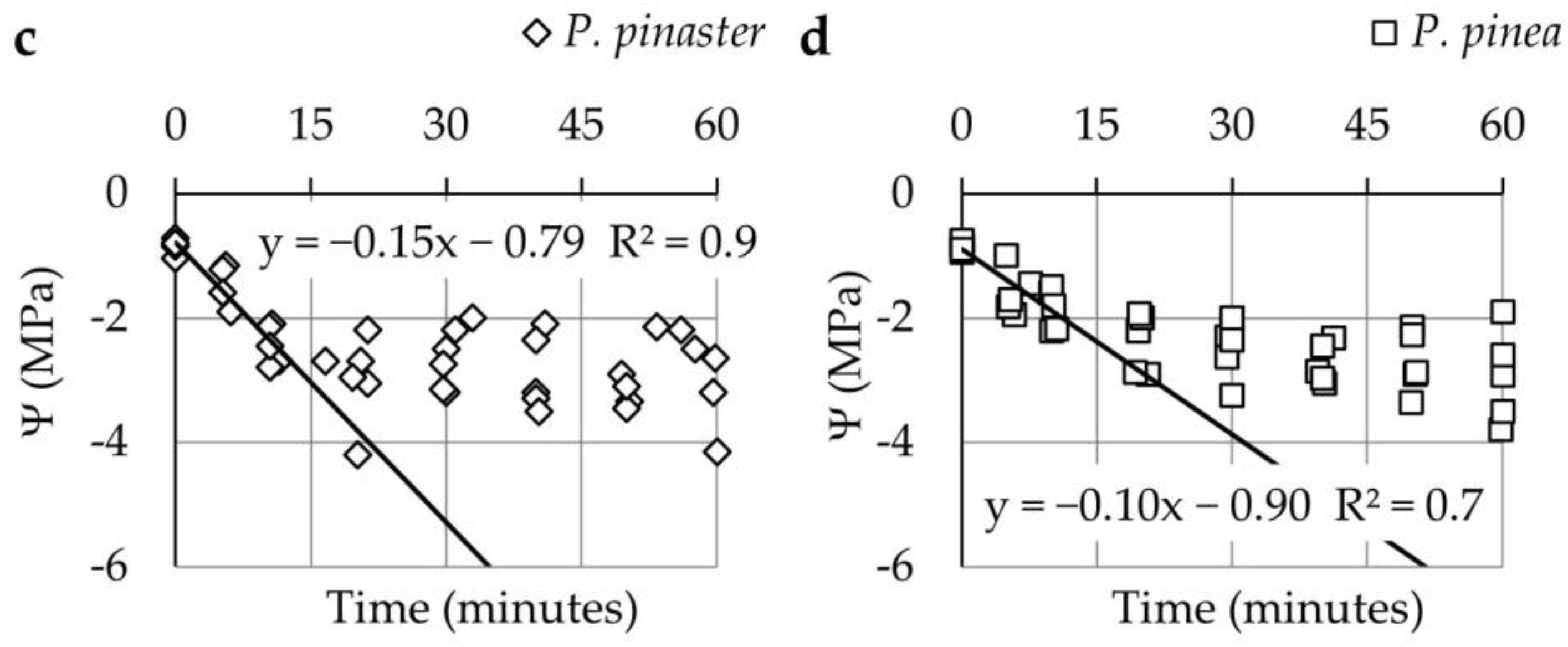
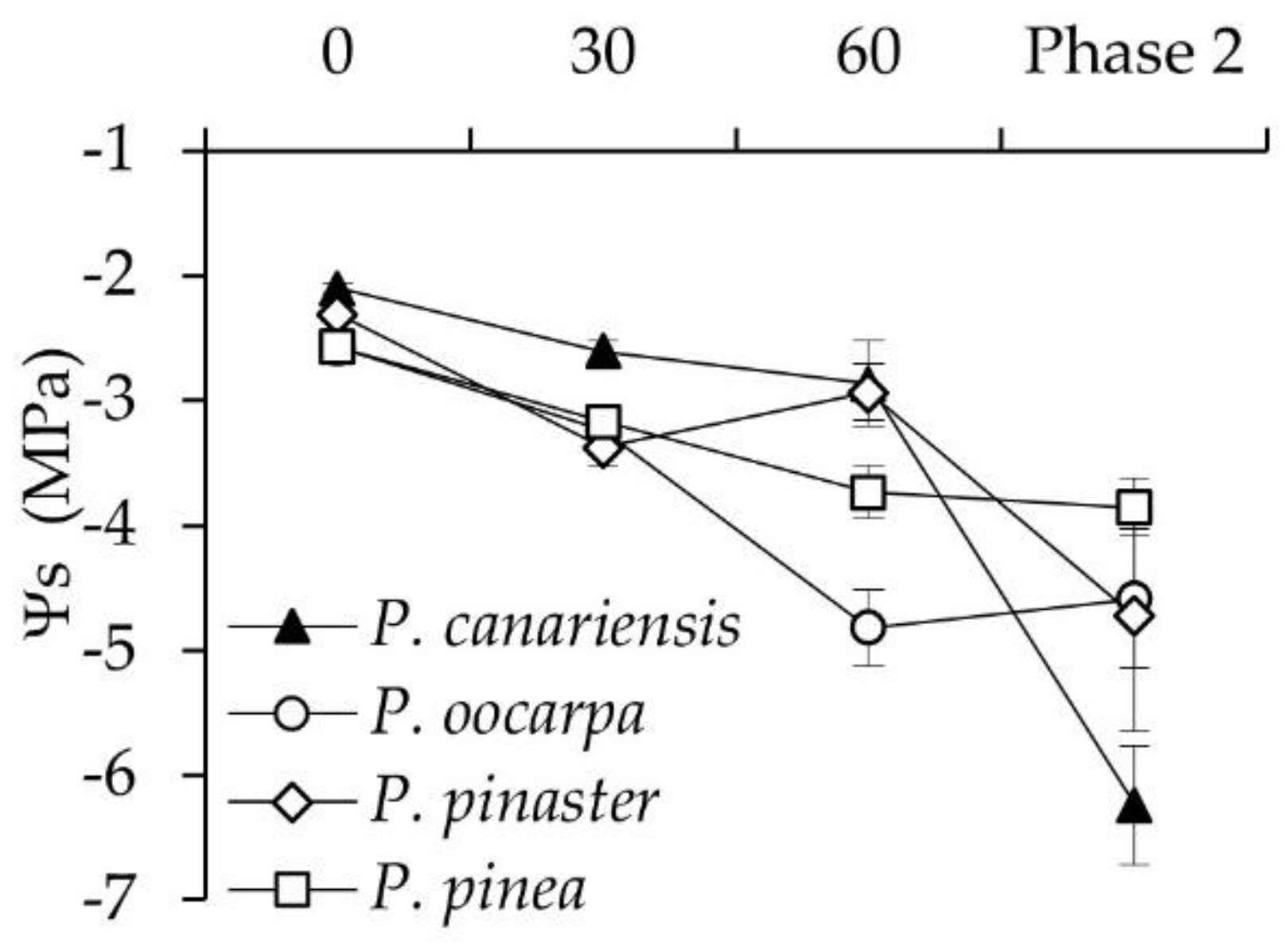
3.1. Changes in Ψ and Ψs during the First and Second Phase of Hot Wind
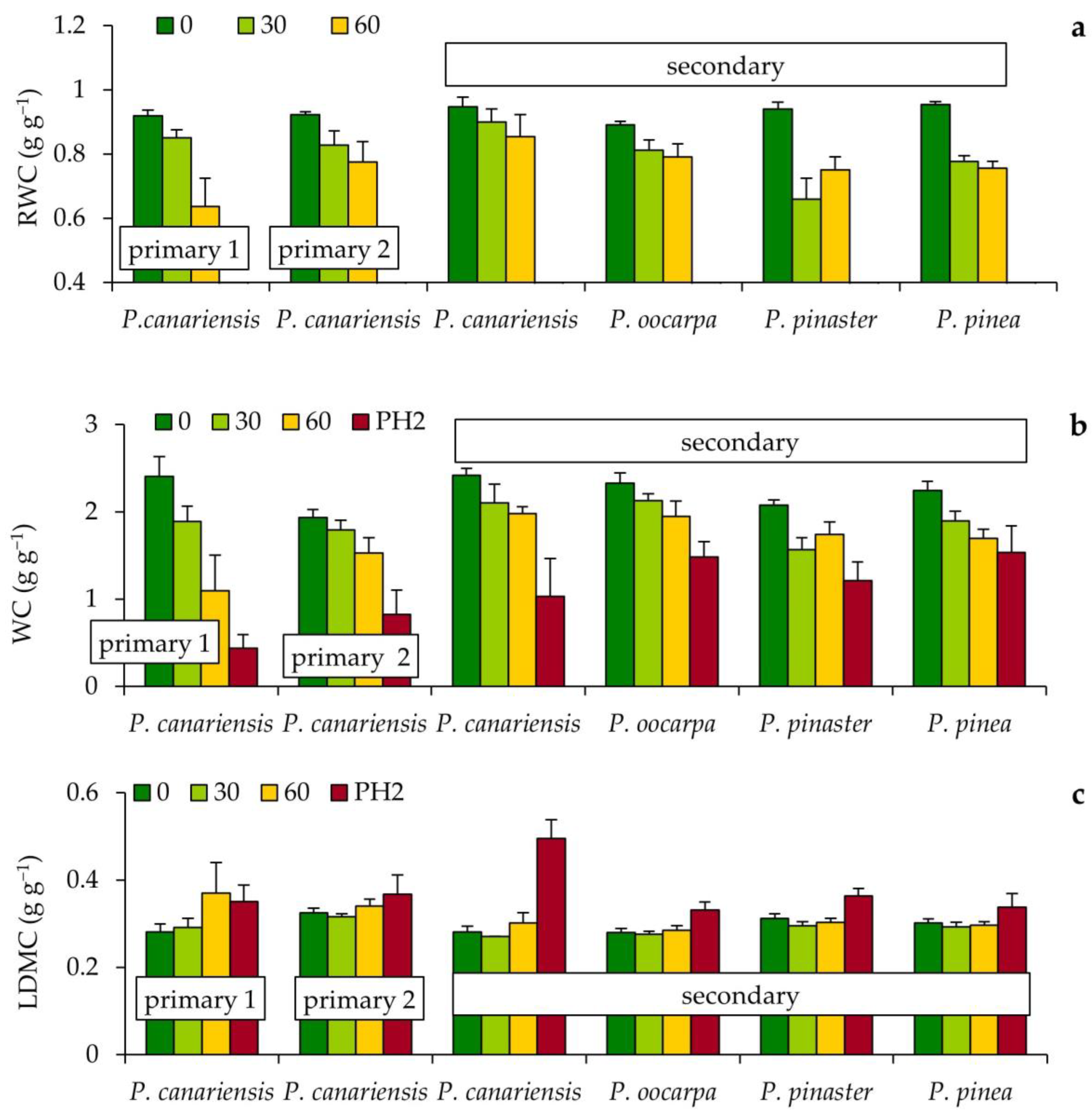
3.2. Changes in Needle Water Content during the First and Second Phases of Hot Wind
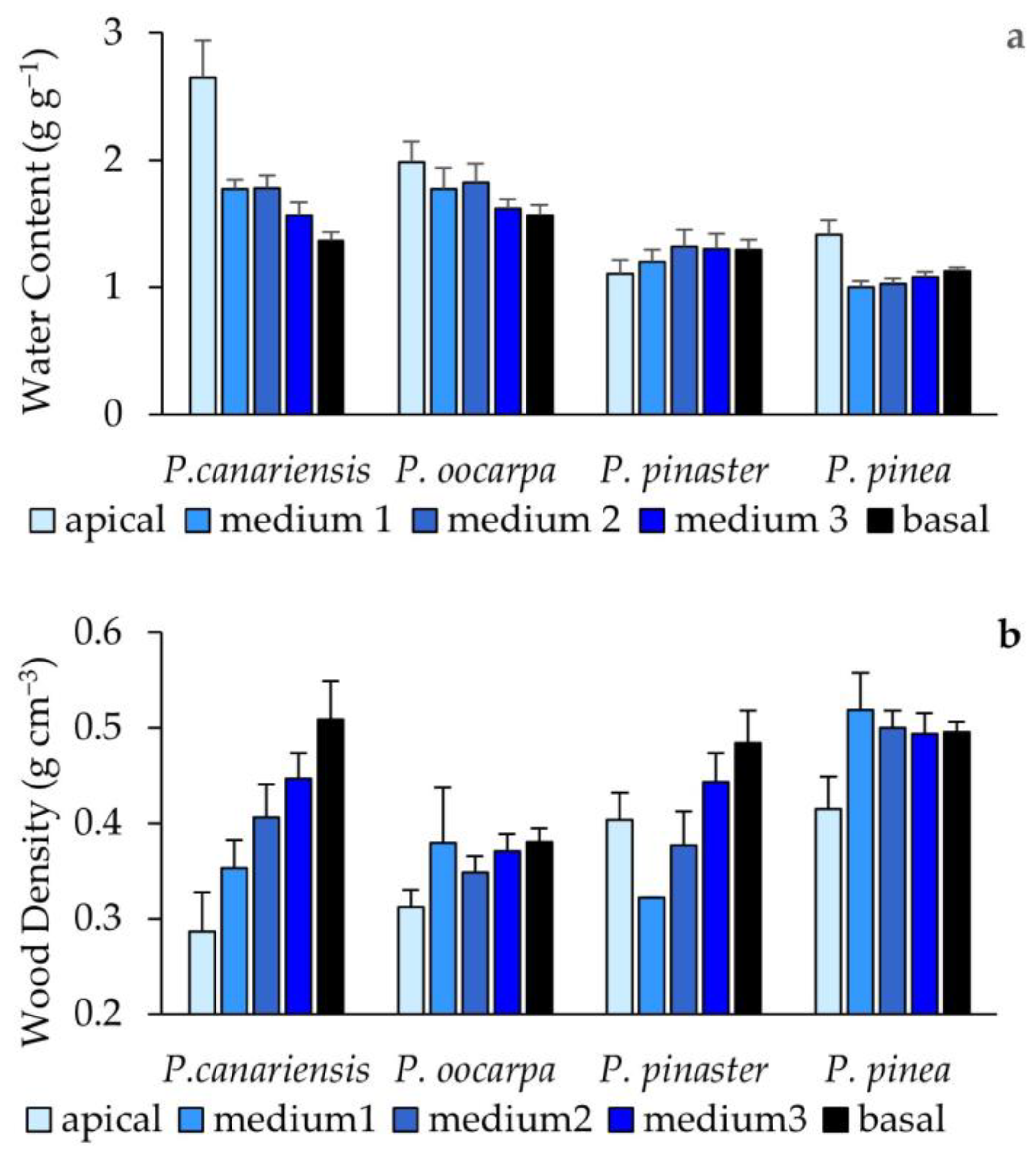
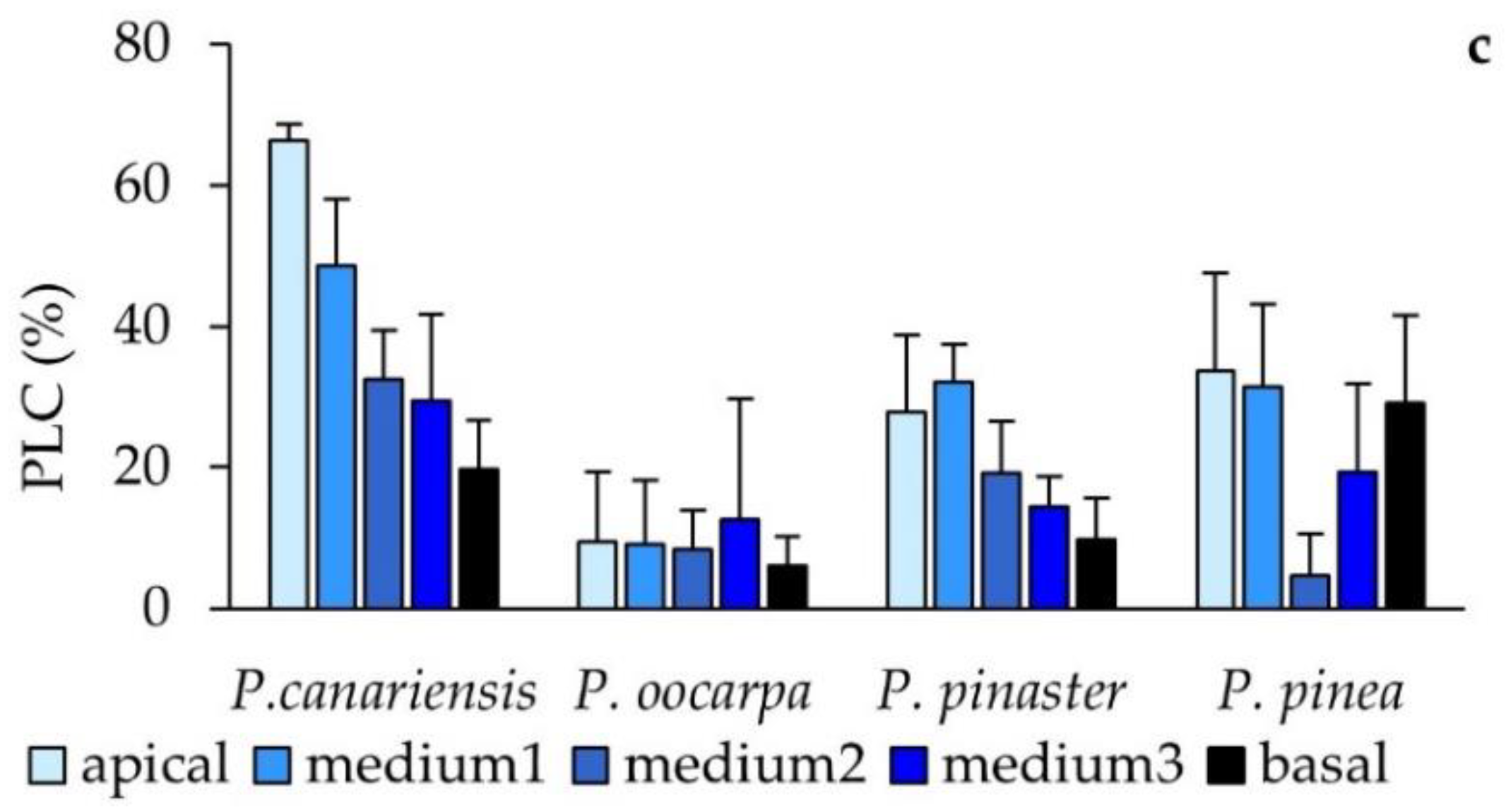
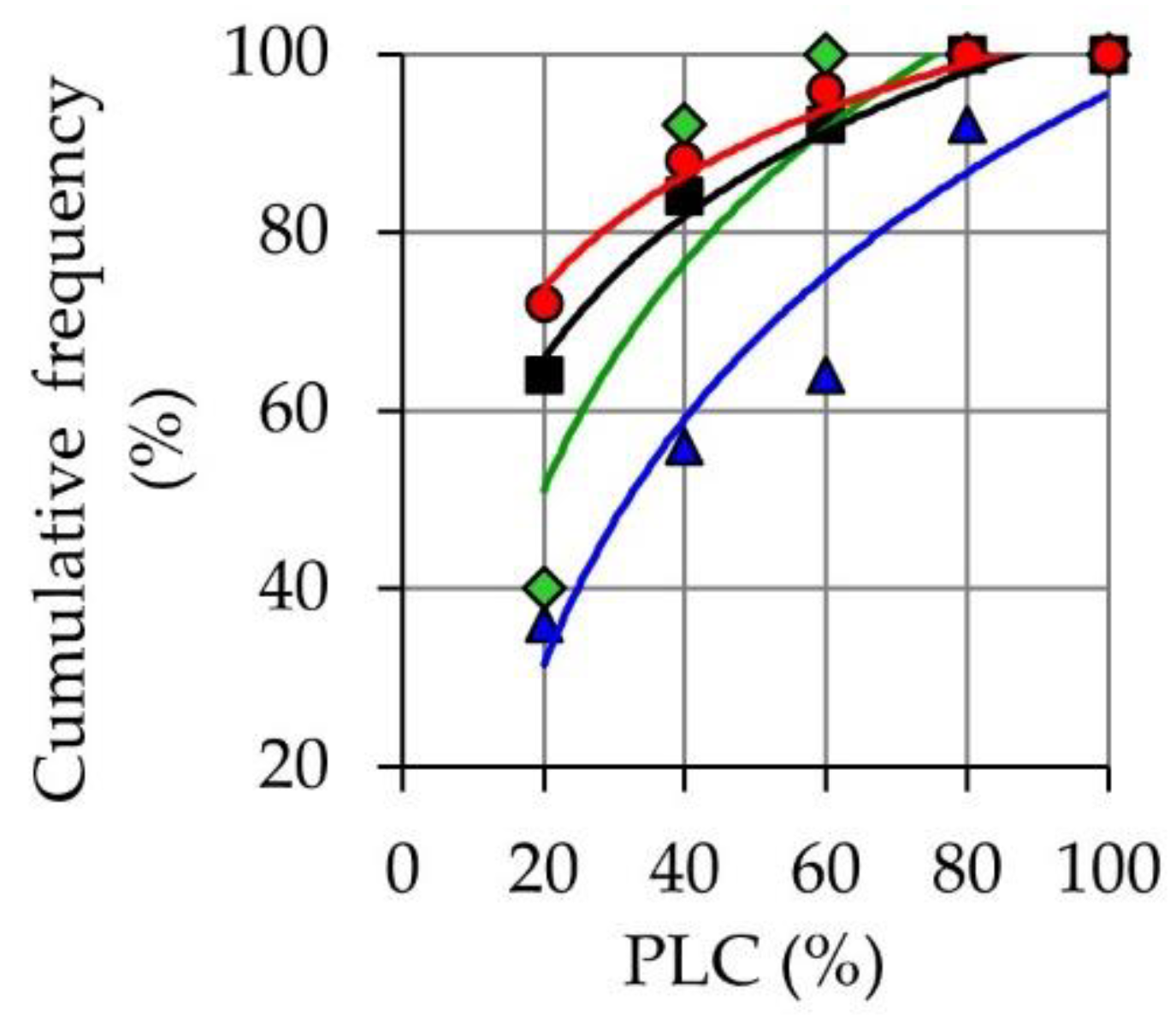
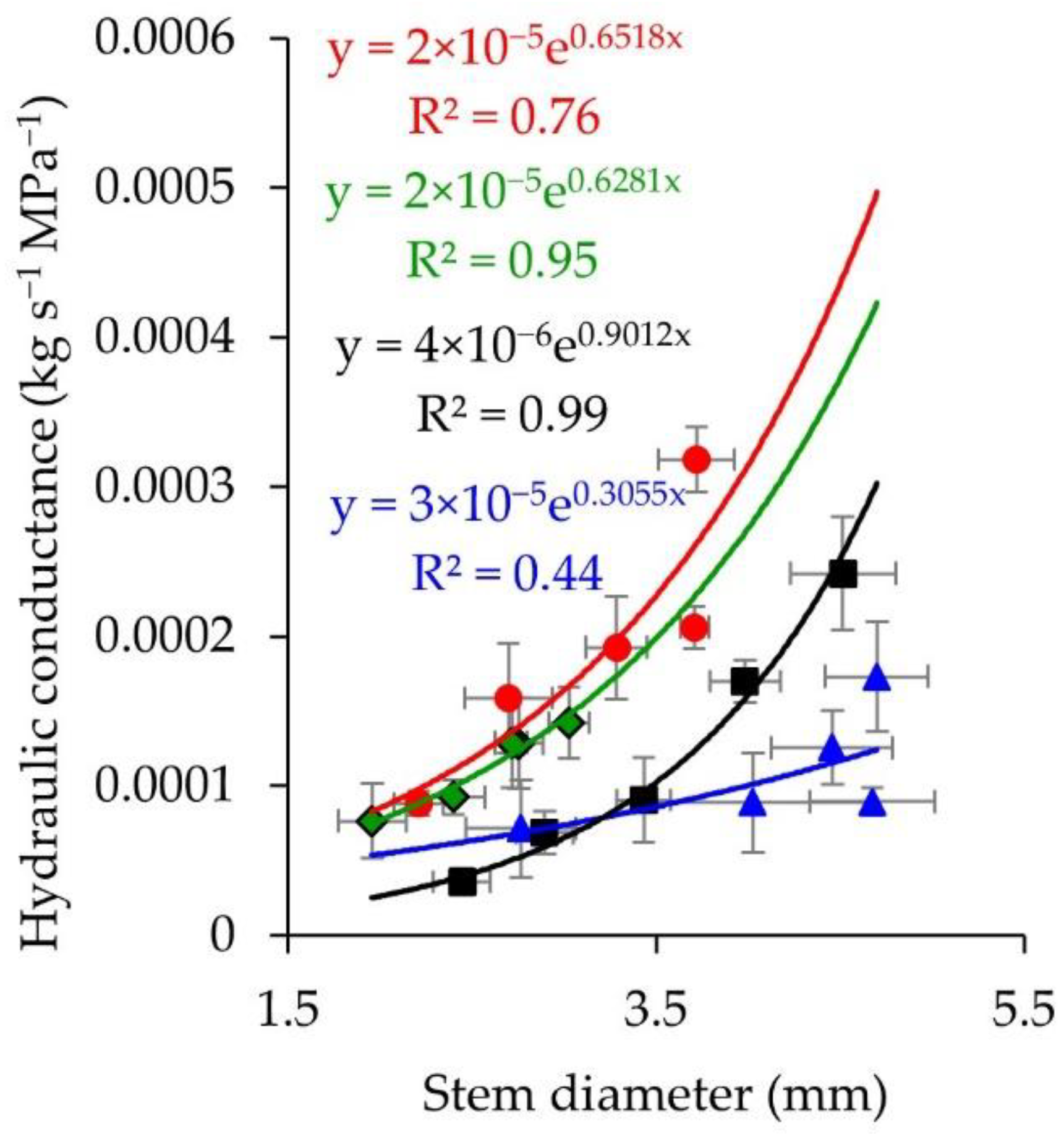
3.3. Stem Water Content, Hydraulic Conductance, and Stem Density Measured after the Second Phase of Hot Wind
3.4. Resprouting
4. Discussion
4.1. Changes in Needle Water Potential under Hot Wind: Resprouting vs. Non-Resprouting Species
4.2. Changes in Needle Osmotic Potential under Hot Wind: P. canariensis vs. P. oocarpa
4.3. Differences between Species in Stem Density and Stem Hydraulic Conductance
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, D.M.; Bradford, J.B.; Lauenroth, W.K. Early indicators of change: Divergent climate envelopes between tree life stages imply range shifts in the western United States. Glob. Ecol. Biogeogr. 2014, 23, 168–180. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Bazzaz, F.A. Changes in drought response strategies with ontogeny in Quercus rubra: Implications for scaling from seedlings to mature trees. Oecologia 2000, 124, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Rank, R.; Maneta, M.; Higuera, P.; Holden, Z.; Dobrowski, S. Conifer Seedling Survival in Response to High Surface Temperature Events of Varying Intensity and Duration. Front. For. Glob. Chang. 2022, 4, 731267. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, M.; Mildrexler, D.J.; Motesharrei, S.; Mu, Q.; Kalnay, E.; Zhao, F.; Li, S.; Wang, K. Potential and Actual impacts of deforestation and afforestation on land surface temperature. J. Geophys. Res. Atmos. 2016, 121, 14–372. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, M.; Motesharrei, S.; Mu, Q.; Kalnay, E.; Li, S. Local cooling and warming effects of forests based on satellite observations. Nat. Commun. 2015, 6, 6603. [Google Scholar] [CrossRef] [PubMed]
- Levitt, J. Responses of Plants to Environmental Stress, Volume 1: Chilling, Freezing, and High Temperature Stresses; Academic Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Nobel, P.S. Extreme temperatures and thermal tolerances for seedlings of desert succulents. Oecologia 1984, 62, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ballantyne, A.P.; Cooper, L.A. Biophysical feedback of global forest fires on surface temperature. Nat. Commun. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Kolb, P.F.; Robberecht, R. High temperature and drought stress effects on survival of Pinus ponderosa seedlings. Tree Physiol. 1996, 16, 665–672. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates Incorporated. Oxford University Press: Oxfort, UK, 2015. [Google Scholar]
- Cunningham, S.C.; Read, J. Foliar temperature tolerance of temperate and tropical evergreen rain forest trees of Australia. Tree Physiol. 2006, 26, 1435–1443. [Google Scholar] [CrossRef]
- Hüve, K.; Bichele, I.; Rasulov, B.; Niinemets, U. When it is too hot for photosynthesis: Heat-induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H2O2 formation. Plant Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.; Sillmann, J.; Fischer, E.M. Top ten European heatwaves since 1950 and their occurrence in the coming decades. Environ. Res. Lett. 2015, 10, 124003. [Google Scholar] [CrossRef]
- López, R.; Ramírez-Valiente, J.A.; Pita, P. How plants cope with heatwaves in a drier environment. Flora 2022, 295, 152148. [Google Scholar] [CrossRef]
- Marchin, R.M.; Backes, D.; Ossola, A.; Leishman, M.R.; Tjoelker, M.G.; Ellsworth, D.S. Extreme heat increases stomatal conductance and drought-induced mortality risk in vulnerable plant species. Glob. Chang. Biol. 2022, 28, 1133–1146. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.E.; Tjoelker, M.G.; Vårhammar, A.; Medlyn, B.E.; Reich, P.B.; Leigh, A.; Pfautsch, S.; Blackman, C.J.; López, R.; Aspinwall, M.J. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Glob. Chang. Biol. 2018, 24, 2390–2402. [Google Scholar] [CrossRef]
- Urli, M.; Porte, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef]
- West, A.G.; Nel, J.A.; Bond, W.J.; Midgley, J.J. Experimental evidence for heat plume-induced cavitation and xylem deformation as a mechanism of rapid post-fire tree mortality. New Phytol. 2016, 211, 828–838. [Google Scholar] [CrossRef]
- Mayr, S. Relevance of time and spatial scales in plant hydraulics. Tree Physiol. 2021, 41, 1781–1784. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Rodrigues, A.C.; Uncles, N.; Rossi, L. Hydraulic segmentation does not protect stems from acute water loss during fire. Tree Physiol. 2021, 41, 1785–1793. [Google Scholar] [CrossRef]
- Kavanagh, K.L.; Dickinson, M.B.; Bova, A.S. A way forward for fire-caused tree mortality prediction: Modeling a physiological consequence of fire. Fire Ecol. 2010, 6, 80–94. [Google Scholar] [CrossRef]
- Song, J.; Trueba, S.; Yin, X.-H.; Cao, K.-F.; Brodribb, T.J.; Hao, G.-Y. Hydraulic vulnerability segmentation in compound-leaved trees: Evidence from an embolism visualization technique. Plant Physiol. 2022, 189, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.J.; Midgley, J.J. The evolutionary ecology of sprouting in woody plants. Int. J. Plant Sci. 2003, 164, S103–S114. [Google Scholar] [CrossRef]
- Budde, K.B.; González-Martínez, S.C.; Navascués, M.; Burgarella, C.; Mosca, E.; Lorenzo, Z.; Zabal-Aguirre, M.; Vendramin, G.G.; Verdú, M.; Pausas, J.G. Increased fire frequency promotes stronger spatial genetic structure and natural selection at regional and local scales in Pinus halepensis Mill. Ann. Bot. 2017, 119, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Karavani, A.; Boer, M.M.; Baudena, M.; Colinas, C.; Díaz-Sierra, R.; Pemán, J.; de Luis, M.; Enríquez-de-Salamanca, Á.; Resco de Dios, V. Fire-induced deforestation in drought-prone Mediterranean forests: Drivers and unknowns from leaves to communities. Ecol. Monogr. 2018, 88, 141–169. [Google Scholar] [CrossRef]
- Lombardi, E.; Shestakova, T.A.; Santini, F.; Resco de Dios, V.; Voltas, J. Harnessing tree-ring phenotypes to disentangle gene by environment interactions and their climate dependencies in a circum-Mediterranean pine. Ann. Bot. 2022, 130, 509–523. [Google Scholar] [CrossRef]
- Santini, F.; Climent, J.M.; Voltas, J. Phenotypic integration and life history strategies among populations of Pinus halepensis: An insight through structural equation modelling. Ann. Bot. 2019, 124, 1161–1171. [Google Scholar] [CrossRef]
- Burrows, G.E. Gymnosperm Resprouting—A Review. Plants 2021, 10, 2551. [Google Scholar] [CrossRef]
- Pensa, A.; Capra, L.; Giordano, G. Ash clouds temperature estimation. Implication on dilute and concentrated PDCs coupling and topography confinement. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Brito, P.; Lorenzo, J.R.; González-Rodríguez, Á.M.; Morales, D.; Wieser, G.; Jimenez, M.S. Canopy transpiration of a Pinus canariensis forest at the tree line: Implications for its distribution under predicted climate warming. Eur. J. For. Res. 2014, 133, 491–500. [Google Scholar] [CrossRef]
- López, R.; López de Heredia, U.; Collada, C.; Cano, F.J.; Emerson, B.C.; Cochard, H.; Gil, L. Vulnerability to cavitation, hydraulic efficiency, growth and survival in an insular pine (Pinus canariensis). Ann. Bot. 2013, 111, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Climent, J.; Chambel, M.R.; López, R.; Mutke, S.; Alía, R.; Gil, L. Population divergence for heteroblasty in the Canary Island pine (Pinus canariensis, Pinaceae). Am. J. Bot. 2006, 93, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Romero, C.; Guzmán-Reyna, R.R.; Rehfeldt, G.E. Altitudinal genetic variation among Pinus oocarpa populations in Michoacán, Mexico: Implications for seed zoning, conservation, tree breeding and global warming. For. Ecol. Manag. 2006, 229, 340–350. [Google Scholar] [CrossRef]
- Mutke, S.; Sánchez, J.G.; Chambel, M.R.; Prada, M.A.; Álvarez, D.; Iglesias, S.; Gil, L. Phenotypic plasticity is stronger than adaptative differentiation among Mediterranean stone pine provenances. For. Syst. 2010, 19, 354–366. [Google Scholar] [CrossRef]
- Battipaglia, G.; Savi, T.; Ascoli, D.; Castagneri, D.; Esposito, A.; Mayr, S.; Nardini, A. Effects of prescribed burning on ecophysiological, anatomical and stem hydraulic properties in Pinus pinea L. Tree Physiol. 2016, 36, 1019–1031. [Google Scholar] [CrossRef]
- Gil, L.; López, R.; García-Mateos, Á.; González-Doncel, I. Seed provenance and fire-related reproductive traits of Pinus pinaster in central Spain. Int. J. Wildland Fire 2010, 18, 1003–1009. [Google Scholar] [CrossRef]
- López, R.; Rodríguez-Calcerrada, J.; Gil, L. Physiological and morphological response to water deficit in seedlings of five provenances of Pinus canariensis: Potential to detect variation in drought-tolerance. Trees 2009, 23, 509–519. [Google Scholar] [CrossRef]
- Ripullone, F.; Guerrieri, M.R.; Nole’, A.; Magnani, F.; Borghetti, M. Stomatal conductance and leaf water potential responses to hydraulic conductance variation in Pinus pinaster seedlings. Trees 2007, 21, 371–378. [Google Scholar] [CrossRef]
- Fischer, E.; Sippel, S.; Knutti, R. Increasing probability of record-shattering climate extremes. Nat. Clim. Chang. 2021, 11, 689–695. [Google Scholar] [CrossRef]
- Matías, L.; Castro, J.; Villar-Salvador, P.; Quero, J.L.; Jump, A.S. Differential impact of hotter drought on seedling performance of five ecologically distinct pine species. Plant Ecol. 2017, 218, 201–212. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pittermann, J. Drought experience and cavitation resistance in six shrubs from the Great Basin, Utah. Basic Appl. Ecol. 2000, 1, 31–41. [Google Scholar] [CrossRef]
- Bouche, P.S.; Jansen, S.; Cochard, H.; Burlett, R.; Capdeville, G.; Delzon, S. Embolism resistance of conifer roots can be accurately measured with the flow-centrifuge method. J. Plant Hydraul. 2015, 2, e002. [Google Scholar] [CrossRef]
- Oliveras, I.; Martínez-Vilalta, J.; Jimenez-Ortiz, T.; José Lledó, M.; Escarré, A.; Piñol, J. Hydraulic properties of Pinus halepensis, Pinus pinea and Tetraclinis articulata in a dune ecosystem of Eastern Spain. Plant Ecol. 2003, 169, 131–141. [Google Scholar] [CrossRef]
- Fu, X.; Meinzer, F.C.; Woodruff, D.R.; Liu, Y.Y.; Smith, D.D.; McCulloh, K.A.; Howard, A.R. Coordination and trade-offs between leaf and stem hydraulic traits and stomatal regulation along a spectrum of isohydry to anisohydry. Plant Cell Environ. 2019, 42, 2245–2258. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Gleason, S.M.; Butler, D.W.; Ziemińska, K.; Waryszak, P.; Westoby, M. Stem xylem conductivity is key to plant water balance across Australian angiosperm species. Funct. Ecol. 2012, 26, 343–352. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Meinzer, F.C.; Qi, J.H.; Goldstein, G.; Cao, K.F. Midday stomatal conductance is more related to stem rather than leaf water status in subtropical deciduous and evergreen broadleaf trees. Plant Cell Environ. 2013, 36, 149–158. [Google Scholar] [CrossRef]
- McCulloh, K.A.; Johnson, D.M.; Meinzer, F.C.; Woodruff, D.R. The dynamic pipeline: Hydraulic capacitance and xylem hydraulic safety in four tall conifer species. Plant Cell Environ. 2014, 37, 1171–1183. [Google Scholar] [CrossRef]
- Meinzer, F.C.; James, S.A.; Goldstein, G.; Woodruff, D. Whole-tree water transport scales with sapwood capacitance in tropical forest canopy trees. Plant Cell Environ. 2003, 26, 1147–1155. [Google Scholar] [CrossRef]
- Skelton, R. Of storage and stems: Examining the role of stem water storage in plant water balance. Plant Physiol. 2019, 179, 1433–1434. [Google Scholar] [CrossRef]
- López, R.; Cano, F.J.; Choat, B.; Cochard, H.; Gil, L. Plasticity in Vulnerability to Cavitation of Pinus canariensis Occurs Only at the Driest End of an Aridity Gradient. Front. Plant Sci. 2016, 7, 769. [Google Scholar] [CrossRef] [PubMed]
- Lechthaler, S.; Kiorapostolou, N.; Pitacco, A.; Anfodillo, T.; Petit, G. The total path length hydraulic resistance according to known anatomical patterns: What is the shape of the root-to-leaf tension gradient along the plant longitudinal axis? J. Theor. Biol. 2020, 502, 110369. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Marín, B.; Ruiz-Medina, M.A.; Miranda, J.C.; González-Rodríguez, Á.M. Coexistent heteroblastic needles of adult Pinus canariensis C. Sm. ex DC. in buch trees differ structurally and physiologically. Forests 2021, 12, 341. [Google Scholar] [CrossRef]
- Sack, L.; Holbrook, N.M. Leaf hydraulics. Annu. Rev. Plant Biol. 2006, 57, 361–381. [Google Scholar] [CrossRef] [PubMed]
- Sack, L.; Scoffoni, C. Measurement of leaf hydraulic conductance and stomatal conductance and their responses to irradiance and dehydration using the Evaporative Flux Method (EFM). JoVE 2012, e4179. [Google Scholar]
- Pivovaroff, A.L.; Sack, L.; Santiago, L.S. Coordination of stem and leaf hydraulic conductance in southern California shrubs: A test of the hydraulic segmentation hypothesis. New Phytol. 2014, 203, 842–850. [Google Scholar] [CrossRef]
- Buckley, T.N.; John, G.P.; Scoffoni, C.; Sack, L. How Does Leaf Anatomy Influence Water Transport outside the Xylem? Plant Physiol. 2015, 168, 1616–1635. [Google Scholar] [CrossRef]
- Domec, J.-C.; Palmroth, S.; Oren, R. Effects of Pinus taeda leaf anatomy on vascular and extravascular leaf hydraulic conductance as influenced by N-fertilization and elevated CO2. J. Plant Hydraul. 2016, 3, e007. [Google Scholar] [CrossRef]
- López, R.; Climent, J.; Gil, L. From desert to cloud forest: The non-trivial phenotypic variation of Canary Island pine needles. Trees 2008, 22, 7. [Google Scholar] [CrossRef]
- Luo, Y.; Ho, C.L.; Helliker, B.R.; Katifori, E. Leaf water storage and robustness to intermittent drought: A spatially explicit capacitive model for leaf hydraulics. Front. Plant Sci. 2021, 12, 725995. [Google Scholar] [CrossRef]
- Li, X.; Delzon, S.; Torres-Ruiz, J.; Badel, E.; Burlett, R.; Cochard, H.; Jansen, S.; King, A.; Lamarque, L.J.; Lenoir, N. Lack of vulnerability segmentation in four angiosperm tree species: Evidence from direct X-ray microtomography observation. Ann. For. Sci. 2020, 77, 1–12. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Petruzzellis, F.; Savi, T.; Bacaro, G.; Nardini, A. A simplified framework for fast and reliable measurement of leaf turgor loss point. Plant Physiol. Biochem. 2019, 139, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Pan, S.; Tyree, M.T. The impact of xylem cavitation on water potential isotherms measured by the pressure chamber technique in Metasequoia glyptostroboides Hu & WC Cheng. J. Exp. Bot. 2016, 67, 4571–4580. [Google Scholar]
- Paula, S.; Pausas, J.G. Leaf traits and resprouting ability in the Mediterranean basin. Funct. Ecol. 2006, 20, 941–947. [Google Scholar] [CrossRef]
- Blackman, C.J.; Brodribb, T.J. Two measures of leaf capacitance: Insights into the water transport pathway and hydraulic conductance in leaves. Funct. Plant Biol. 2011, 38, 118–126. [Google Scholar] [CrossRef]
- Nobel, P.S. Chapter 2—Water. In Physicochemical and Environmental Plant Physiology (Fourth Edition); Nobel, P.S., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 44–99. [Google Scholar]
- Deligoz, A.; Gur, M. Morphological, physiological and biochemical responses to drought stress of Stone pine (Pinus pinea L.) seedlings. Acta Physiol. Plant. 2015, 37, 243. [Google Scholar] [CrossRef]
- Domec, J.C.; Palmroth, S.; Ward, E.; Maier, C.A.; Thérézien, M.; Oren, R. Acclimation of leaf hydraulic conductance and stomatal conductance of Pinus taeda (loblolly pine) to long-term growth in elevated CO2 (free-air CO2 enrichment) and N-fertilization. Plant Cell Environ. 2009, 32, 1500–1512. [Google Scholar] [CrossRef]
- Kunert, N.; Tomaskova, I. Leaf turgor loss point at full hydration for 41 native and introduced tree and shrub species from Central Europe. J. Plant Ecol. 2020, 13, 754–756. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Woodruff, D.R.; Marias, D.E.; McCulloh, K.A.; Sevanto, S. Dynamics of leaf water relations components in co-occurring iso-and anisohydric conifer species. Plant Cell Environ. 2014, 37, 2577–2586. [Google Scholar] [CrossRef]
- Nguyen-Queyrens, A.; Bouchet-Lannat, F. Osmotic adjustment in three-year-old seedlings of five provenances of maritime pine (Pinus pinaster) in response to drought. Tree Physiol. 2003, 23, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, L.J.; Stokes, T.A.; Ramirez, M.R.; Mendonca, C.C. Drought tolerance of a Pinus palustris plantation. For. Ecol. Manag. 2019, 451, 117557. [Google Scholar] [CrossRef]
- Lang, I.; Sassmann, S.; Schmidt, B.; Komis, G. Plasmolysis: Loss of turgor and beyond. Plants 2014, 3, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Echeverría, A.; Anfodillo, T.; Soriano, D.; Rosell, J.A.; Olson, M.E. Constant theoretical conductance via changes in vessel diameter and number with height growth in Moringa oleifera. J. Exp. Bot. 2019, 70, 5765–5772. [Google Scholar] [CrossRef]
- Lintunen, A.; Kalliokoski, T. The effect of tree architecture on conduit diameter and frequency from small distal roots to branch tips in Betula pendula, Picea abies and Pinus sylvestris. Tree Physiol. 2010, 30, 1433–1447. [Google Scholar] [CrossRef]
- Olson, M.E.; Rosell, J.A. Vessel diameter–stem diameter scaling across woody angiosperms and the ecological causes of xylem vessel diameter variation. New Phytol. 2013, 197, 1204–1213. [Google Scholar] [CrossRef]
- Kuusk, V.; Niinemets, Ü.; Valladares, F. A major trade-off between structural and photosynthetic investments operative across plant and needle ages in three Mediterranean pines. Tree Physiol. 2018, 38, 543–557. [Google Scholar] [CrossRef]
- Ehmig, M.; Coiro, M.; Linder, H.P. Ecophysiological strategy switch through development in heteroblastic species of mediterranean ecosystems—An example in the African Restionaceae. Ann. Bot. 2019, 123, 611–623. [Google Scholar] [CrossRef]
- Rose, K.M.; Mickelbart, M.V.; Jacobs, D.F. Plasticity of phenotype and heteroblasty in contrasting populations of Acacia koa. Ann. Bot. 2019, 124, 399–409. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pita, P.; López, R.; Gil, L. The Effect of Hot Wind on Needle and Stem Water Status: Response Strategies in Resprouting and Non-Resprouting Pine Species. Forests 2023, 14, 2174. https://doi.org/10.3390/f14112174
Pita P, López R, Gil L. The Effect of Hot Wind on Needle and Stem Water Status: Response Strategies in Resprouting and Non-Resprouting Pine Species. Forests. 2023; 14(11):2174. https://doi.org/10.3390/f14112174
Chicago/Turabian StylePita, Pilar, Rosana López, and Luis Gil. 2023. "The Effect of Hot Wind on Needle and Stem Water Status: Response Strategies in Resprouting and Non-Resprouting Pine Species" Forests 14, no. 11: 2174. https://doi.org/10.3390/f14112174