
Early Growth Characterization and Antioxidant Responses of Phellodendron chinense Seedling in Response to Four Soil Types at Three Growth Stages
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Chemical Analysis
2.3. Seedling Culture and Growth Parameters Measurement
2.4. Determination of Chlorophyll Contents
2.5. Estimation of Soluble Protein (SP) and Total Polyphenols (TP) Contents
2.6. Determination of Malondialdehyde (MDA) and Soluble Sugar (SS) Contents
2.7. Determination of Superoxide Dismutase (SOD), Peroxidase (POD), Phenylalanine Ammonialyase (PAL), and Polyphenol Oxidase (PPO) Activities
2.8. Statistical Analysis
3. Results
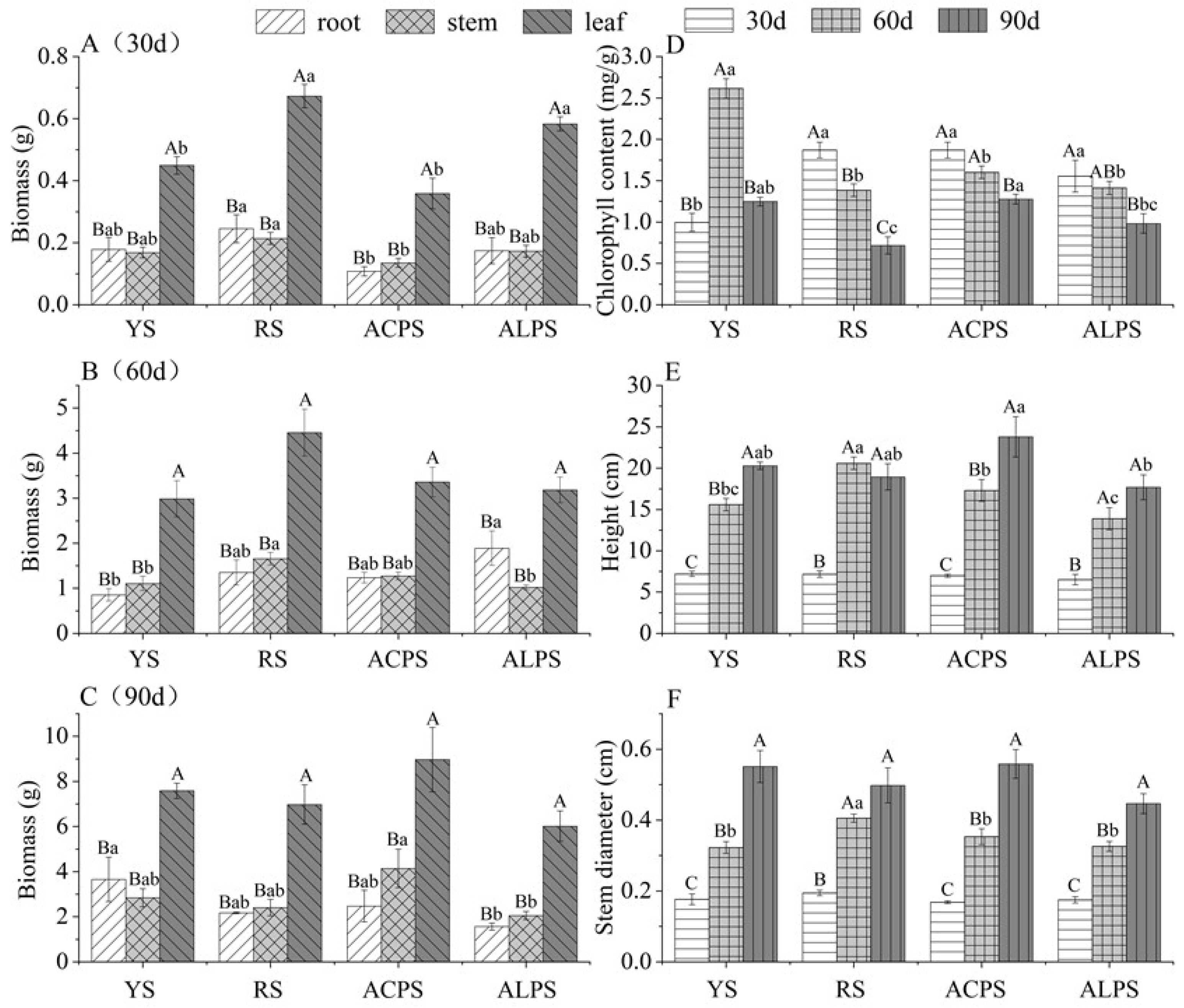
3.1. Growth Parameter and Chlorophyll Contents
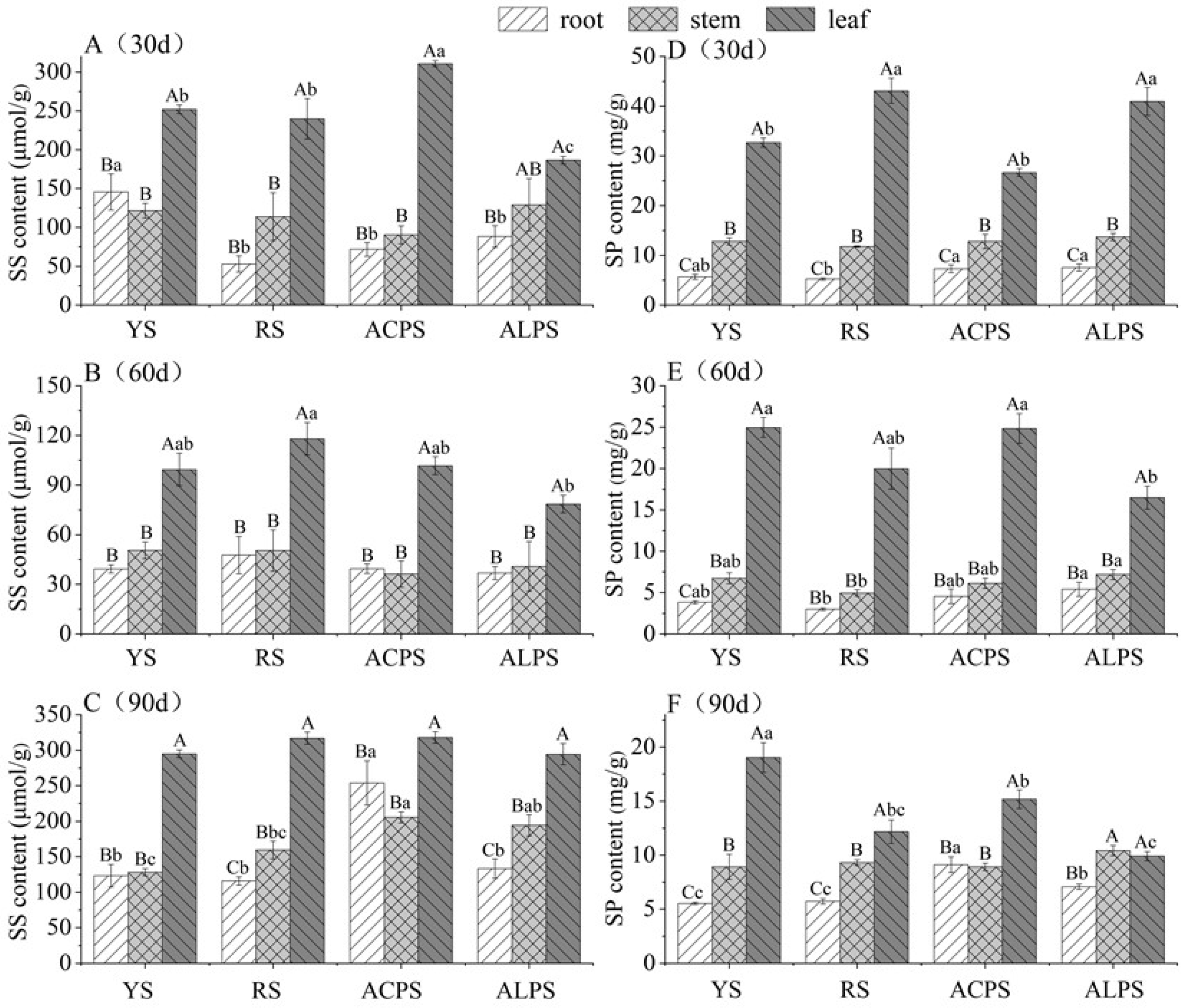
3.2. Soluble Sugar (SS) and Soluble Protein (SP) Contents
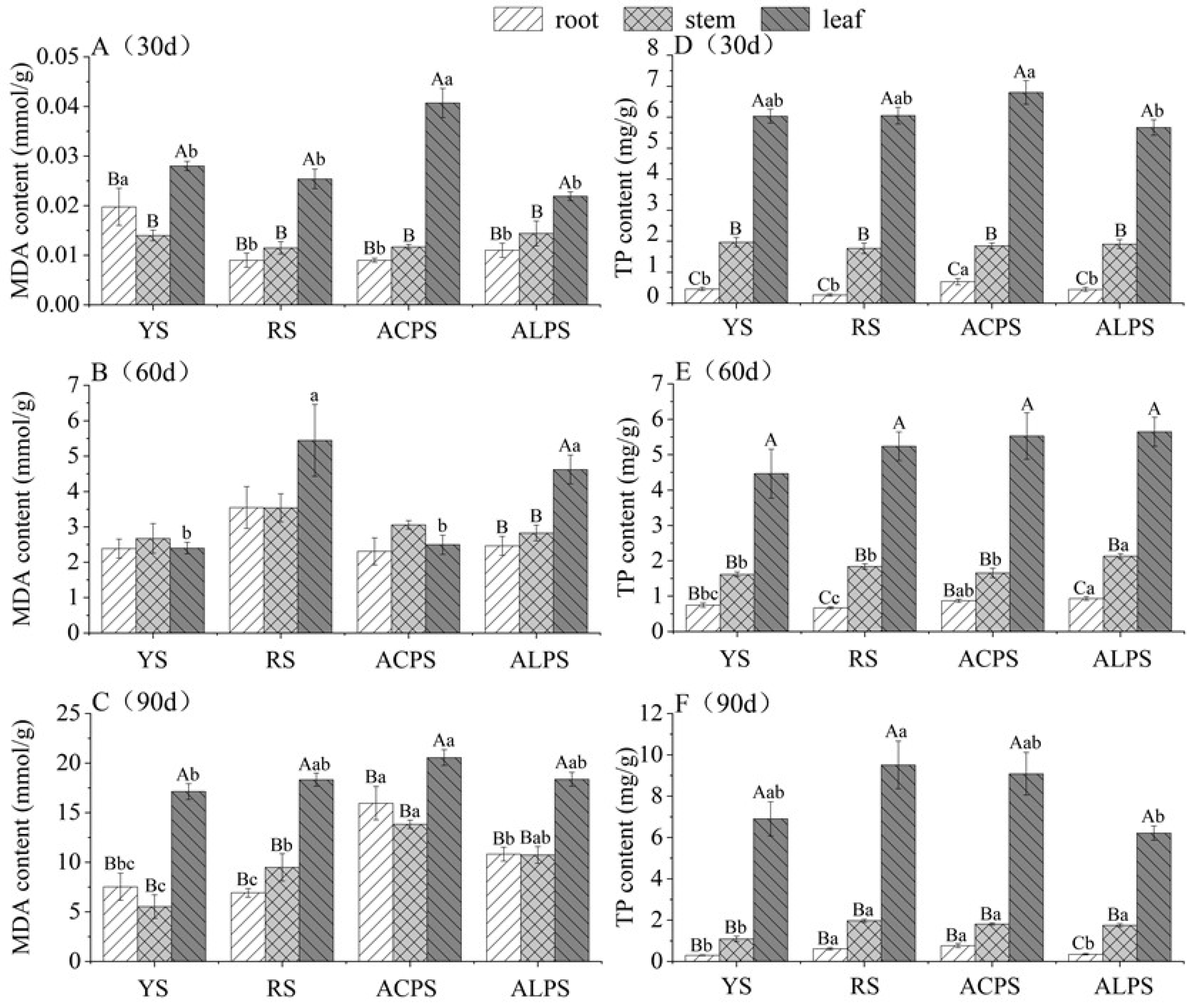
3.3. Malondialdehyde (MDA) and Total Polyphenol (TP) Contents
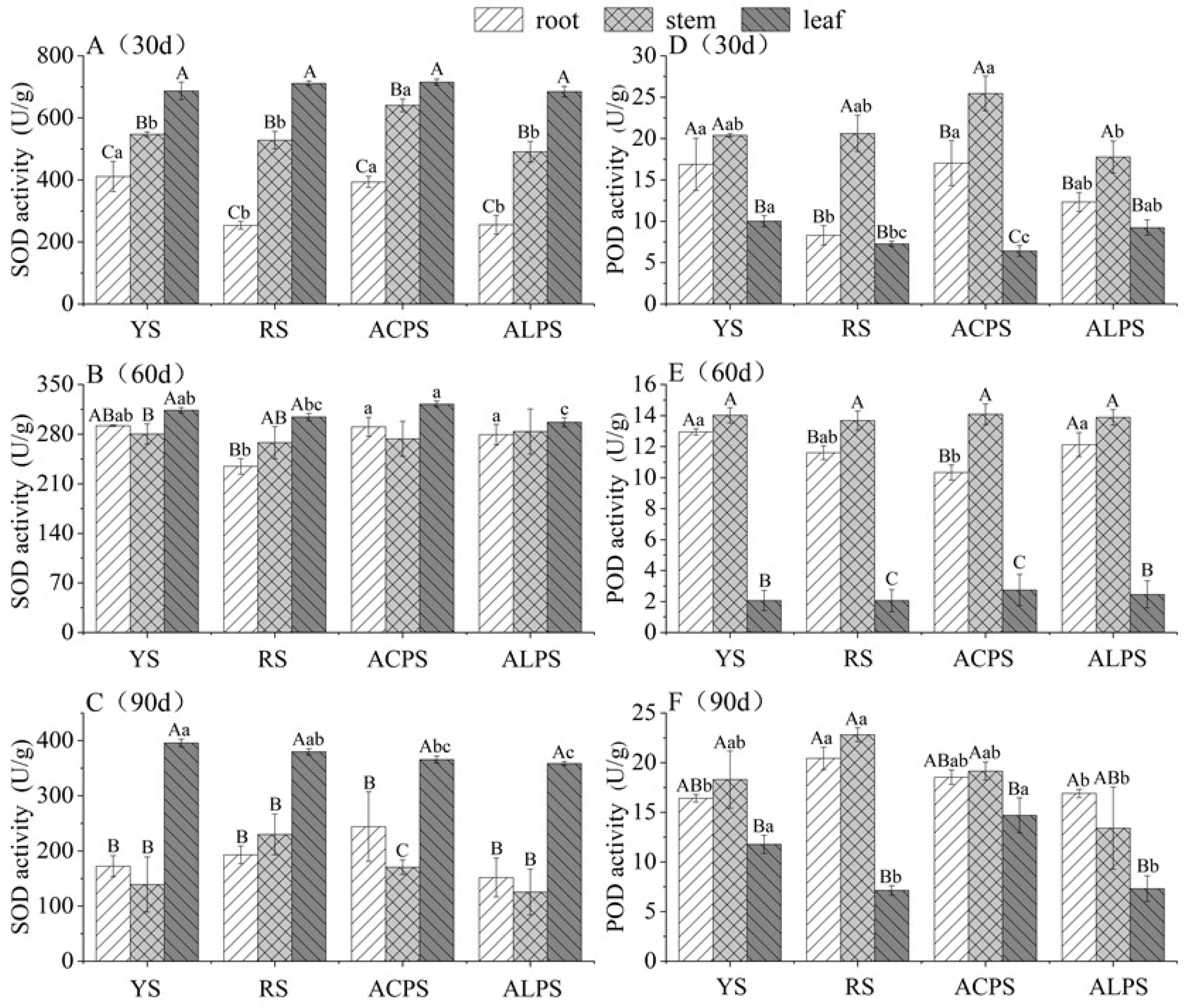
3.4. Superoxide Dismutase (SOD) and Peroxidase (POD) Activities
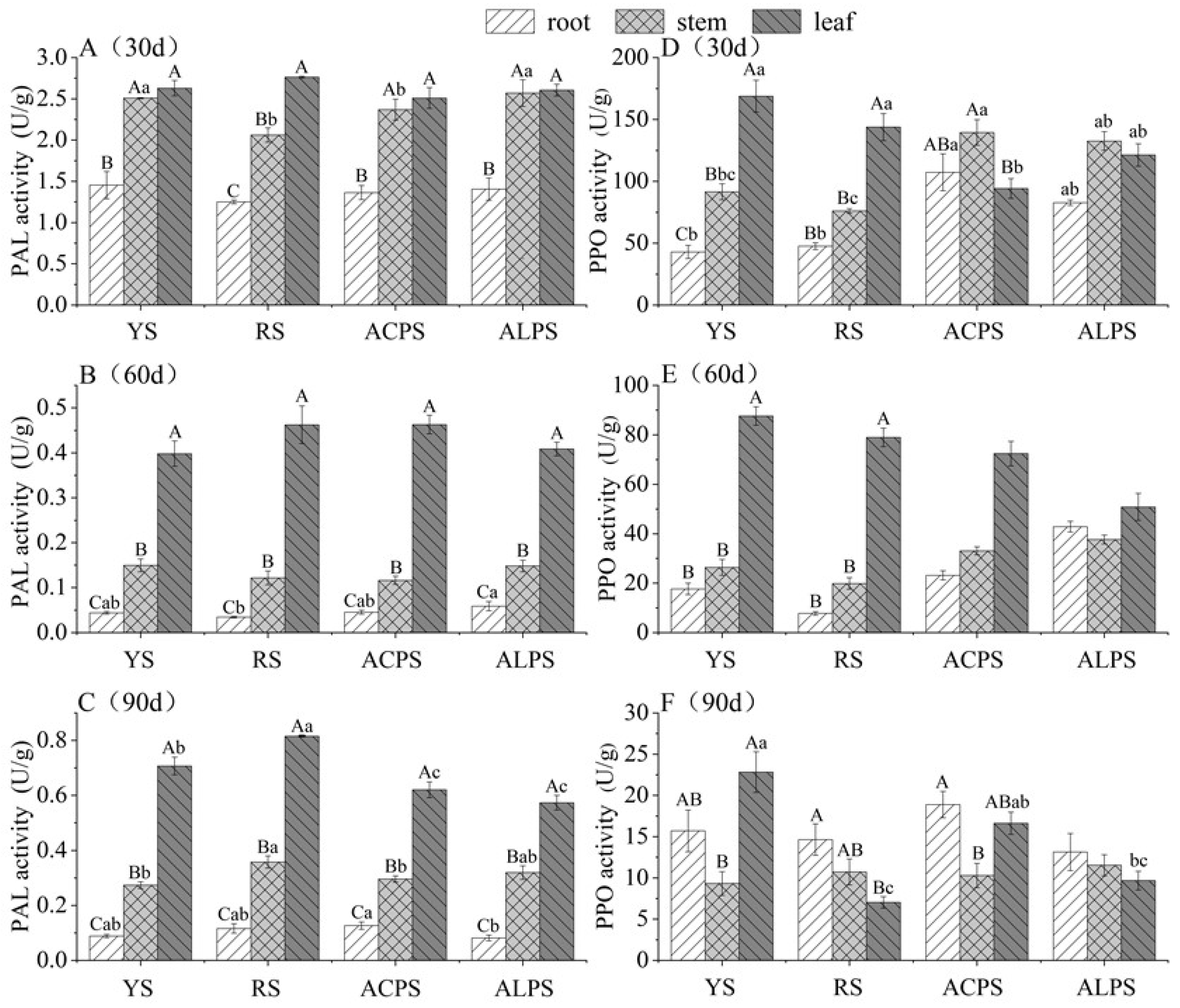
3.5. Phenylalanine Ammonia Lyase (PAL) and Polyphenol Oxidase (PPO) Activities
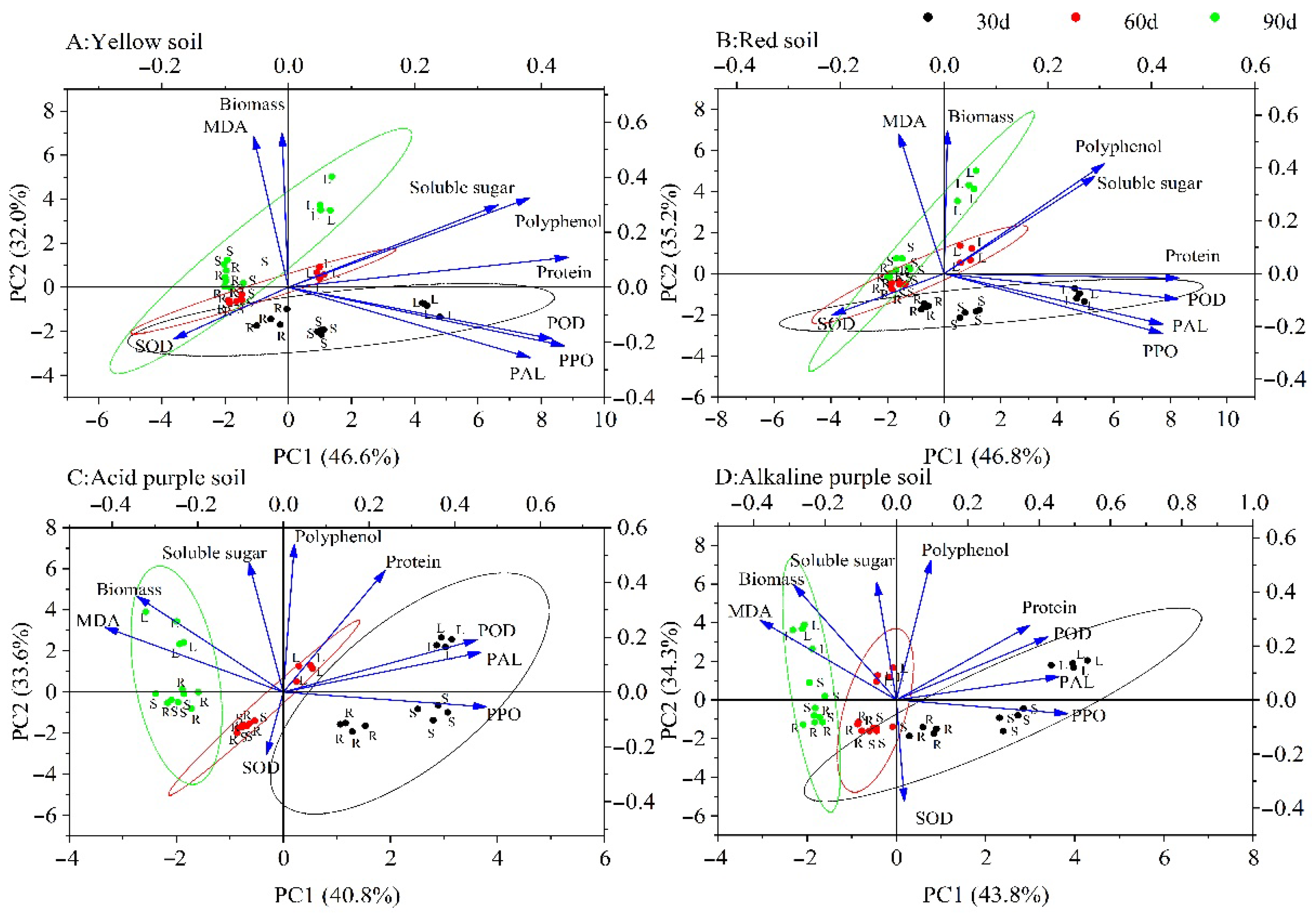
3.6. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahul, B.; Rishikesh, S.; Raghubanshi, A.S. Understanding the ecology of tree-seedling growth in dry tropical environment: A management perspective. Energy Ecol. Environ. 2016, 1, 296–309. [Google Scholar]
- Kahkashan, P.; Najat, B.; Iram, S.; Iffat, S. Influence of soil type on the growth parameters, essential oil yield and biochemical contents of Mentha arvensis L. J. Essent. Oil Bear. Plants 2016, 19, 76–81. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Omer, E.A.; Abou Hussein, E.A.; Hendawy, S.F.; Ezz, E.D.; Azza, A.; El Gendy, A.G. Effect of soil type and seasonal variation on growth, yield, essential oil and artemisinin content of Artemisia annua L. Int. Res. J. Hortic. 2013, 1, 15–27. [Google Scholar] [CrossRef]
- Zhi, X.; Song, Y.; Yu, D.; Qian, W.; He, M.; Lin, X.; Zhang, D.; Gao, S. Early growth characterization and C: N: P stoichiometry in Firmiana simplex seedlings in response to shade and soil types. Forests 2023, 14, 1481. [Google Scholar] [CrossRef]
- Breugel, M.; Craven, D.; Lai, H.R.; Baillon, M.; Turner, B.L.; Hall, J.S. Soil nutrients and dispersal limitation shape compositional variation in secondary tropical forests across multiple scales. J. Ecol. 2019, 107, 566–581. [Google Scholar] [CrossRef]
- Yavitt, J.B.; Wright, S.J. Seedling growth responses to water and nutrient augmentation in the understorey of a lowland moist forest, Panama. J. Trop. Ecol. 2008, 24, 19–26. [Google Scholar] [CrossRef]
- Sinclair, A.H. Availability of magnesium to ryegrass from soils during intensive cropping in the glasshouse. J. Agric. Sci. 1981, 96, 635–642. [Google Scholar] [CrossRef]
- Fetene, M. Intra- and inter-specific competition between seedlings of Acacia etbaica and a perennial grass (Hyparrenia hirta). J. Arid. Environ. 2003, 55, 441–451. [Google Scholar] [CrossRef]
- Bailly, C. The signalling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef] [PubMed]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.N.; Zhou, Z.Y.; Yan, S.Y.; Qin, Y. Effect of salt stress on physiological and biochemical characteristics of Amorpha fruticosa seedlings. Acta Pratacul. Sin. 2011, 20, 84–90. [Google Scholar]
- Rasheed, F.; Gondal, A.; Kudus, K.A.; Zafar, Z.; Nawaz, M.F.; Khan, W.R.; Abdullah, M.; Ibrahim, F.H.; Depardieu, C.; Pazi, A.M.M. Effects of soil water deficit on three tree species of the arid environment: Variations in growth, physiology, and antioxidant enzyme activities. Sustainability 2021, 13, 3336. [Google Scholar] [CrossRef]
- Kouki, R.; Ayachi, R.; Ferreira, R.; Sleimi, N. Behavior of Cucumis sativus L. in presence of aluminum stress: Germination, plant growth, and antioxidant enzymes. Food Sci. Nutr. 2021, 9, 3280–3288. [Google Scholar] [CrossRef]
- Chowardhara, B.; Borgohain, P.; Saha, B.; Awasthi, J.P.; Panda, S.K. Differential oxidative stress responses in Brassica juncea (L.) Czern and Coss cultivars induced by cadmium at germination and early seedling stage. Acta Physiol. Plant. 2020, 42, 105. [Google Scholar] [CrossRef]
- Luera, P.; Wahl-Villarreal, K.; Christoffersen, B.O.; Treviño, A.; Soti, P.; Gabler, C.A. Effects of scarification, phytohormones, soil type, and warming on the germination and/or seedling performance of three Tamaulipan Thornscrub forest species. Plants 2021, 10, 1489. [Google Scholar] [CrossRef]
- Sun, Y.; Lenon, G.B.; Yang, A.W.H. Phellodendri cortex: A phytochemical, pharmacological, and pharmacokinetic review. Evid. Based Complement. Altern. Med. 2019, 2019, 7621929. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lim, H.S.; Kim, Y.; Lee, J.; Kim, B.Y.; Jeong, S.J. Phytochemical quantification and the in vitro acetylcholinesterase inhibitory activity of Phellodendron chinense and its components. Molecules 2017, 22, 925. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.; Ren, H.; Yang, Y.; Gu, T.; He, J.; Gao, S. Effects of nitrogen on growth parameters and photosynthetic characteristics of P. chinense Schneid. seedlings in three soil types. J. Elem. 2023, 28, 279–294. [Google Scholar]
- Ball, D. Loss-on-ignition as an estimate of organic matter and organic carbon in non-calcareous soils. J. Soil Sci. 1964, 15, 84–92. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fabbri, A.; Galaverna, G.; Ganino, T. Polyphenol composition of olive leaves with regard to cultivar, time of collection and shoot type. Acta Hortic. 2008, 791, 95–102. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplast. I. Kinetics and stochiometery of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–190. [Google Scholar] [CrossRef]
- Gao, S.; Ou-yang, C.; Tang, L.; Zhu, J.; Xu, Y.; Wang, S.; Chen, F. Growth and antioxidant responses in Jatropha curcas seedling exposed to mercury toxicity. J. Hazard. Mater. 2010, 182, 591–597. [Google Scholar] [CrossRef]
- He, X.J.; Bowler, P.A.; Lin, L. Bioaccumulation and antioxidative responses in Hydrilla verticillata (L.f.) Royle exposed to copper toxicity. Fresen. Environ. Bull. 2022, 31, 6069–6078. [Google Scholar]
- Traus, I.; Mrak, T.; Ferlan, M.; Eleznik, P.; Kraigher, H. Influence of soil temperature on growth traits of European beech seedlings. Can. J. For. Res. 2015, 45, 246–251. [Google Scholar]
- Shu, X.; Zhang, K.R.; Zhang, Q.F.; Wang, W.B. Ecophysiological responses of Jatropha curcas L. seedlings to simulated acid rain under different soil types. Ecotoxicol. Environ. Saf. 2019, 185, 109705. [Google Scholar] [CrossRef]
- Zhi, X.; Yang, Y.; Zou, J.; Ma, N.; Liu, T.; Wang, H.; Hu, Y.; Gao, S. Responses of the growth and nutrient stoichiometry in Ricinus communis seedlings on four soil types. J. Elem. 2022, 27, 223–238. [Google Scholar] [CrossRef]
- Delfim, J.J.; Salvador, J.O.; Moraes, L.A.C.; Moreira, A. Effect of liming and phosphorus sources on soil fertility, growth, and nutritional status of guava seedlings in three tropical soils. J. Plant Nutr. 2022, 45, 1403–1412. [Google Scholar] [CrossRef]
- Simkin, A.J.; Kapoor, L.; Doss, C.G.P.; Hofmann, T.A.; Lawson, T.; Ramamoorthy, S. The role of photosynthesis related pigments in light harvesting, photoprotection and enhancement of photosynthetic yield in planta. Photosynth. Res. 2022, 152, 23–42. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, L.; Simkin, A.J.; George Priya Doss, C.; Siva, R. Fruit ripening: Dynamics and integrated analysis of carotenoids and anthocyanins. BMC Plant Biol. 2022, 22, 27. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.K.; Daiya, K.S.; Chawan, D.D. Effect of different soil types on plant growth, leaf pigments and sennoside content in Cassia species. Pharm. Weekbl. Sci. Ed. 1980, 2, 573–575. [Google Scholar] [CrossRef]
- Meena, S.K.; Rakshit, A.; Meena, V.S. Effect of seed bio-priming and N doses under varied soil type on nitrogen use efficiency (NUE) of wheat (Triticum aestivum L.) under greenhouse condition. Biocatal. Agric. Biotechnol. 2016, 6, 68–75. [Google Scholar] [CrossRef]
- Sebetha, E.T.; Mfanta, A.; Adebayo, R. Effect of different phosphorus fertilizer sources, cultivar and soil type on growth performance of soybean. Res. Crops 2018, 19, 616–621. [Google Scholar]
- Quan, M.; Liang, J. The influences of four types of soil on the growth, physiological and biochemical characteristics of lycoris aurea (L’her.) herb. Sci. Rep. 2017, 7, 43284. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shen, H. Effect of electrostatic field on seed germination and seedling growth of Sorbus pohuashanesis. J. For. Res. 2011, 22, 27–34. [Google Scholar] [CrossRef]
- Ogundola, A.F.; Bvenura, C.; Ehigie, A.F.; Afolayan, A.J. Effects of soil types on phytochemical constituents and antioxidant properties of Solanum nigrum. S. Afr. J. Bot. 2021, 151, 325–333. [Google Scholar] [CrossRef]
- Suwa, R.; Tajima, H.; Gima, S.; Uehara, N.; Watanabe, K.; Yabuta, S.; Tominaga, J.; Kawamitsu, Y. Polyphenol production in Peucedanum japonicum Thunb. varies with soil type and growth stage. Hortic. J. 2018, 87, 382–388. [Google Scholar] [CrossRef]
- Kaushik, D.; Aryadeep, R. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar]
- Gong, Q.; Wang, L.; Dai, T.; Zhou, J.; Kang, Q.; Chen, H.; Li, K.; Li, Z. Effects of copper on the growth, antioxidant enzymes and photosynthesis of spinach seedlings. Ecotoxicol. Environ. Saf. 2019, 171, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.H.; Fahad, S.; Khan, S.U.; Din, M.; Ullah, A.; Sabagh, A.E.; Hossain, A.; Llanes, A.; Liu, L. Copper-induced oxidative stress, initiation of antioxidants and phytoremediation potential of flax (Linum usitatissimum L.) seedlings grown under the mixing of two different soils of China. Environ. Sci. Pollut. Res. Int. 2020, 27, 5211–5221. [Google Scholar] [CrossRef]
- Yue, X.H.; Miao, L.F.; Yang, F.; Nawaz, M. Morphological and physiological responses of Dalbergia odorifera T. Chen seedlings to different culture substances. PLoS ONE 2020, 15, e0232051. [Google Scholar] [CrossRef] [PubMed]
- El-Badri, A.M.; Batool, M.A.A.; Mohamed, I.; Wang, Z.; Khatab, A.; Sherif, A.; Ahmad, H.; Khan, M.N.; Hassan, H.M.; Elrewainy, I.M.; et al. Antioxidative and metabolic contribution to salinity stress responses in two rapeseed cultivars during the early seedling stage. Antioxidants 2021, 10, 1227. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Arora, R. Dynamics of the antioxidant system during seed osmopriming, post-priming germination, and seedling establishment in Spinach (Spinacia oleracea). Plant Sci. 2011, 180, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Romero-Rodríguez, M.C.; Archidona-Yuste, A.; Abril, N.; Gil-Serrano, A.M.; Meijón, M.; Jorrín-Novo, J.V. Germination and early seedling development in Quercus ilex recalcitrant and non-dormant seeds: Targeted transcriptional, hormonal, and sugar analysis. Front. Plant Sci. 2018, 9, 1508. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, J.; Xiao, Z.; Lu, T.; Ren, X.; Wei, H. Effects of acid rain on plant growth: A meta-analysis. J. Environ. Manag. 2021, 297, 113213. [Google Scholar] [CrossRef]
- Dyhr-Jensen, K.; Brix, H. Effects of pH on ammonium uptake by Typha latifolia L. Plant Cell Environ. 1996, 19, 1431–1436. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Yellow Soil | Red Soil | Acidic Purple Soil | Alkaline Purple Soil |
|---|---|---|---|---|
| pH | 4.805 ± 0.018 b | 4.990 ± 0.075 b | 4.343 ± 0.044 c | 8.733 ± 0.105 a |
| Moisture content (%) | 0.115 ± 0.012 a | 0.021 ± 0.006 c | 0.120 ± 0.002 a | 0.05 ± 0.001 b |
| Total nitrogen (g·kg−1) | 17.492 ± 1.580 b | 11.301 ± 0.431 c | 28.513 ± 0.163 a | 15.906 ± 0.884 b |
| Ammonium nitrogen (mg·kg−1) | 19.784 ± 0.916 a | 18.667 ± 0.362 a | 11.123 ± 1.214 b | 13.259 ± 1.971 b |
| Nitrate nitrogen (mg·kg−1) | 55.696 ± 2.207 a | 7.190 ± 0.563 c | 26.410 ± 0.895 b | 2.051 ± 0.571 d |
| Total phosphorous (g·kg−1) | 0.076 ± 0.005 c | 0.069 ± 0.006 c | 0.235 ± 0.007 b | 0.301 ± 0.011 a |
| Available phosphorous (g·kg−1) | 0.0546 ± 0.004 b | 0.0338 ± 0.001 c | 0.0512 ± 0.002 b | 0.065 ± 0.002 a |
| Total potassium (g·kg−1) | 13.460 ± 0.251 b | 8.844 ± 0.775 c | 12.170 ± 0.200 b | 19.180 ± 0.851 a |
| Organic matter (g·kg−1) | 42.014 ± 8.623 a | 16.643 ± 0.445 b | 42.527 ± 2.655 a | 20.989 ± 0.425 b |
| Indexes | YS | RS | ACPS | ALPS | ||||
|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | PC1 | PC2 | PC1 | PC2 | |
| Biomass | −0.01 | 0.56 | 0.01 | 0.54 | −0.34 | 0.35 | −0.29 | 0.42 |
| Soluble protein | 0.44 | 0.11 | 0.45 | −0.02 | 0.24 | 0.44 | 0.37 | 0.28 |
| Soluble sugar | 0.33 | 0.30 | 0.29 | 0.37 | −0.08 | 0.47 | −0.06 | 0.43 |
| Total polyphenols | 0.38 | 0.32 | 0.31 | 0.42 | 0.03 | 0.54 | 0.10 | 0.52 |
| Malondialdehyde (MDA) | −0.05 | 0.55 | −0.09 | 0.53 | −0.42 | 0.24 | −0.38 | 0.29 |
| Superoxide dismutase (SOD) | −0.18 | −0.19 | −0.22 | −0.16 | −0.04 | −0.23 | 0.02 | −0.37 |
| Peroxidase (POD) | 0.42 | −0.19 | 0.45 | −0.10 | 0.45 | 0.19 | 0.42 | 0.23 |
| Phenylalanine ammonialyase (PAL) | 0.38 | −0.26 | 0.42 | −0.19 | 0.46 | 0.14 | 0.46 | 0.09 |
| Polyphenol oxidase (PPO) | 0.44 | −0.21 | 0.42 | −0.23 | 0.47 | −0.05 | 0.48 | −0.05 |
| Eigenvalue | 4.19 | 2.88 | 4.21 | 3.17 | 3.67 | 3.03 | 3.95 | 3.09 |
| Percentage of variance (%) | 46.58 | 32.01 | 46.80 | 35.17 | 40.81 | 33.63 | 43.84 | 34.30 |
| Cumulative (%) | 78.58 | 81.97 | 74.44 | 78.14 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Hu, Y.-Y.; Qian, W.-Z.; Wang, Y.-J.; Ren, H.-Y.; Gao, S.; Cao, G.-X. Early Growth Characterization and Antioxidant Responses of Phellodendron chinense Seedling in Response to Four Soil Types at Three Growth Stages. Forests 2023, 14, 1746. https://doi.org/10.3390/f14091746
Yang Y, Hu Y-Y, Qian W-Z, Wang Y-J, Ren H-Y, Gao S, Cao G-X. Early Growth Characterization and Antioxidant Responses of Phellodendron chinense Seedling in Response to Four Soil Types at Three Growth Stages. Forests. 2023; 14(9):1746. https://doi.org/10.3390/f14091746
Chicago/Turabian StyleYang, Yao, Yun-Yi Hu, Wen-Zhang Qian, Ya-Juan Wang, Hong-Yu Ren, Shun Gao, and Guo-Xing Cao. 2023. "Early Growth Characterization and Antioxidant Responses of Phellodendron chinense Seedling in Response to Four Soil Types at Three Growth Stages" Forests 14, no. 9: 1746. https://doi.org/10.3390/f14091746