
Complementation of an Escherichia coli K-12 Mutant Strain Deficient in KDO Synthesis by Forming D-Arabinose 5-Phosphate from Glycolaldehyde with Fructose 6-Phosphate Aldolase (FSA)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Reference |
|---|---|---|
| E. coli K-12 | ||
| BW25113 | lacIq rrnBT14 ∆lacZWJ16 hsdR514 ∆araBADAH33 ∆rhaBADLD78 | [16] |
| BW25113 ∆gutQ | BW25113, ΔgutQ::FRT | [18] |
| BW25113 ∆gutQ ∆kdsD::kan | BW25113, ΔgutQ::FRT ∆kdsD::FRT-kan-FRT | This study a |
| BW25113 ∆gutQ ∆kdsD | BW25113, ΔgutQ::FRT ∆kdsD::FRT | This study a |
| BW25113 ∆gutQ ∆kdsD ∆rbsK::Ptac-fsaAA129S | BW25113, ΔgutQ::FRT ∆kdsD::FRT ∆rbsK::Ptac-fsaAA129S | This study b |
2.2. Recombinant DNA Techniques
| Plasmid | Characteristics | Reference |
|---|---|---|
| pJF119EH | Ptac, lacIq, RBS, AmpR | [19] |
| pJF119fsaA | fsaA gene cloned into pJF119EH (XbaI/HindIII) | [20] |
| pJF119fsaAA129S | fsaAA129S gene cloned into pJF119EH (XbaI/HindIII) | [20] |
| pJF119fsaB | fsaB gene cloned into pJF119EH (EcoRI/HindIII) | This study |
| pKD46 | repA101 (Ts), araC, ParaB-ϒ-β-exo (red recombinase), AmpR | [16] |
| pCO1-cat | Ptac-aroC-aroA, oriR6’ (first 36 bp), FRT-cat-FRT, lacIq, AmpR, CmR | [21] |
| pCP20 | FLP+, λ cl857+, λ pR Repts, AmpR, CmR | [22] |
| pCas | repA101 (Ts), Pcas-cas9, ParaB-ϒ-β-exo (red recombinase), lacIq, Ptrc-sgRNA-pMB1, KmR | [17] |
| pTargetF-Cm-rbsK | pMB1, sgRNA-rbsK, CmR | [23] |
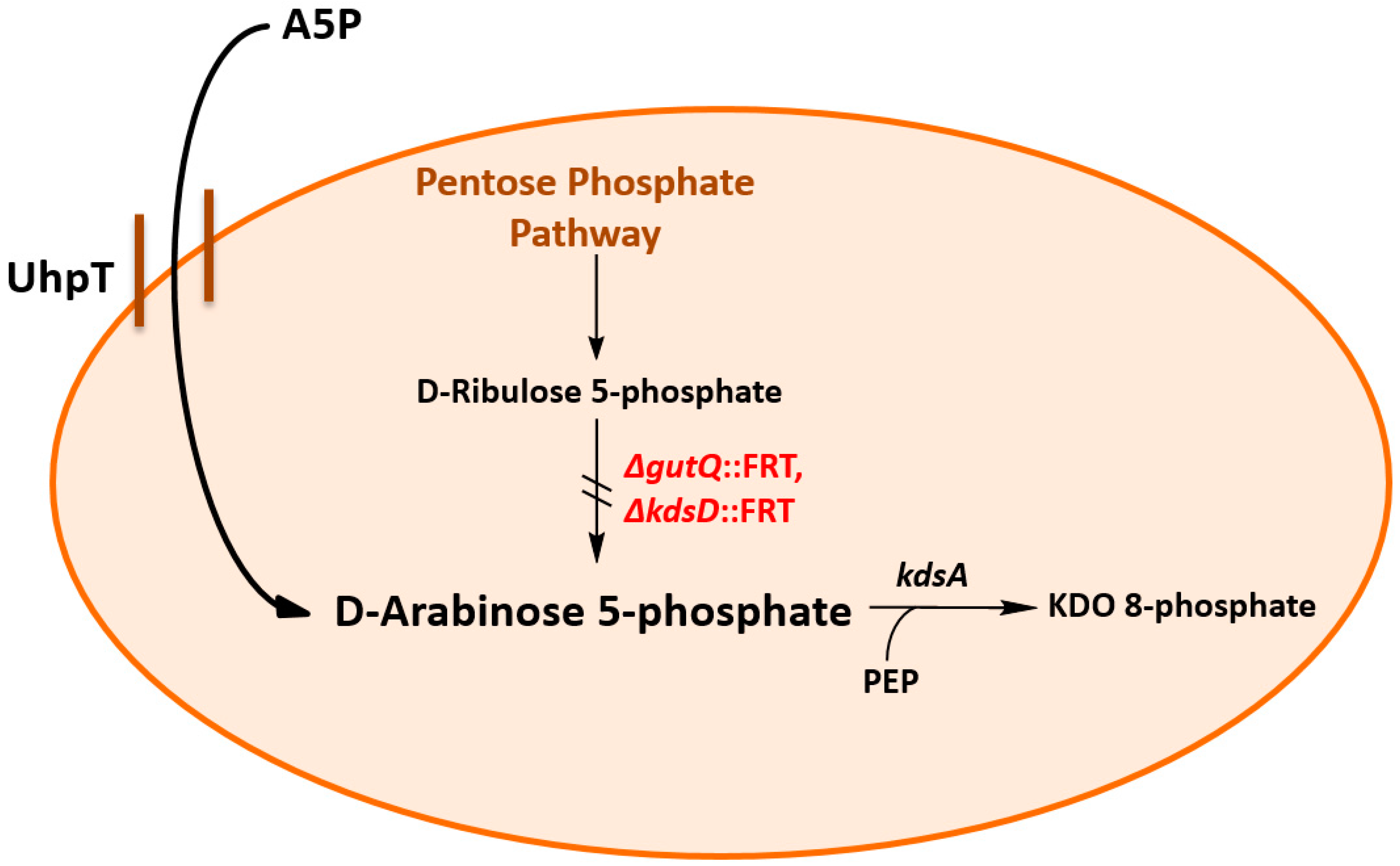
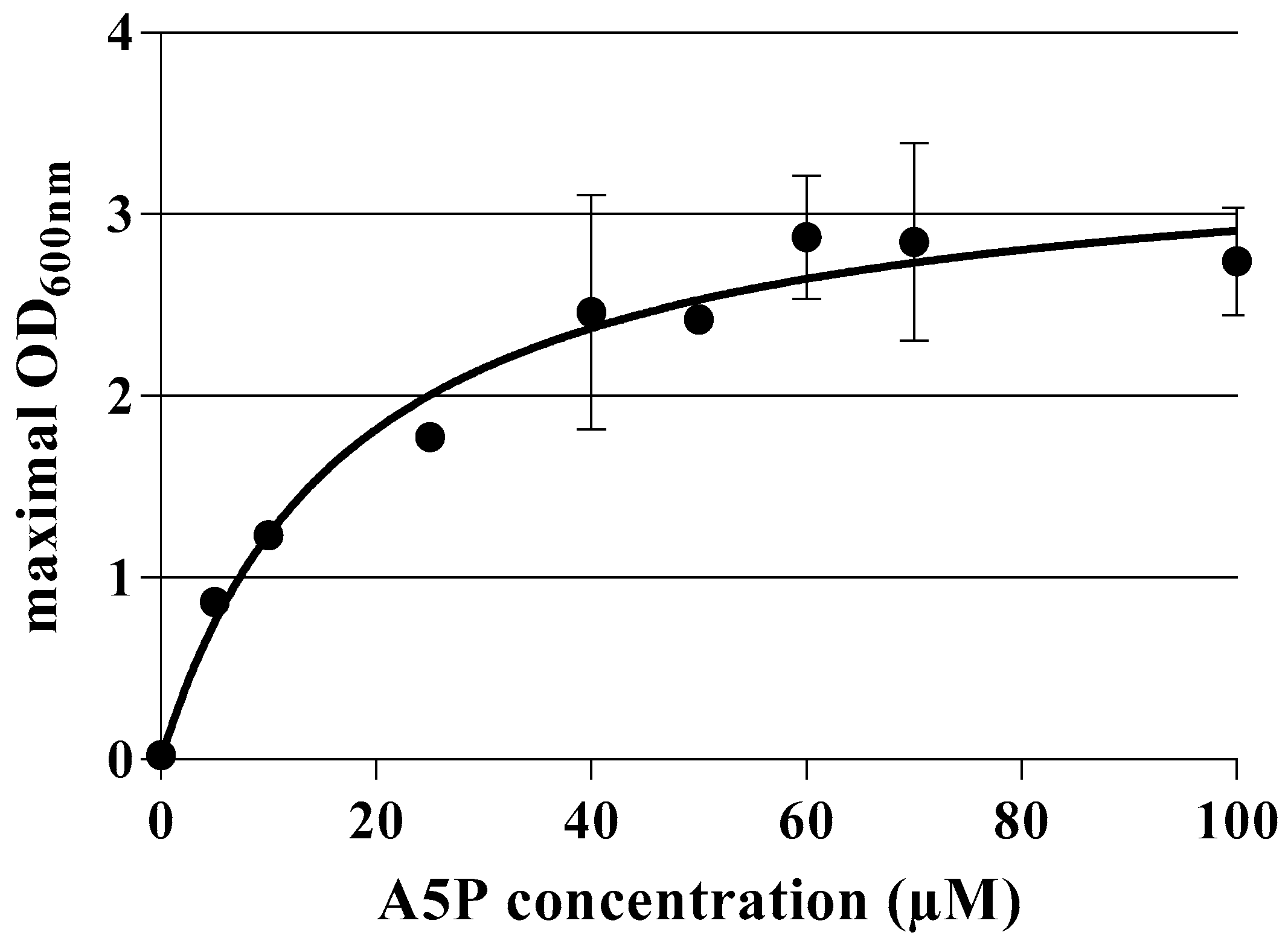
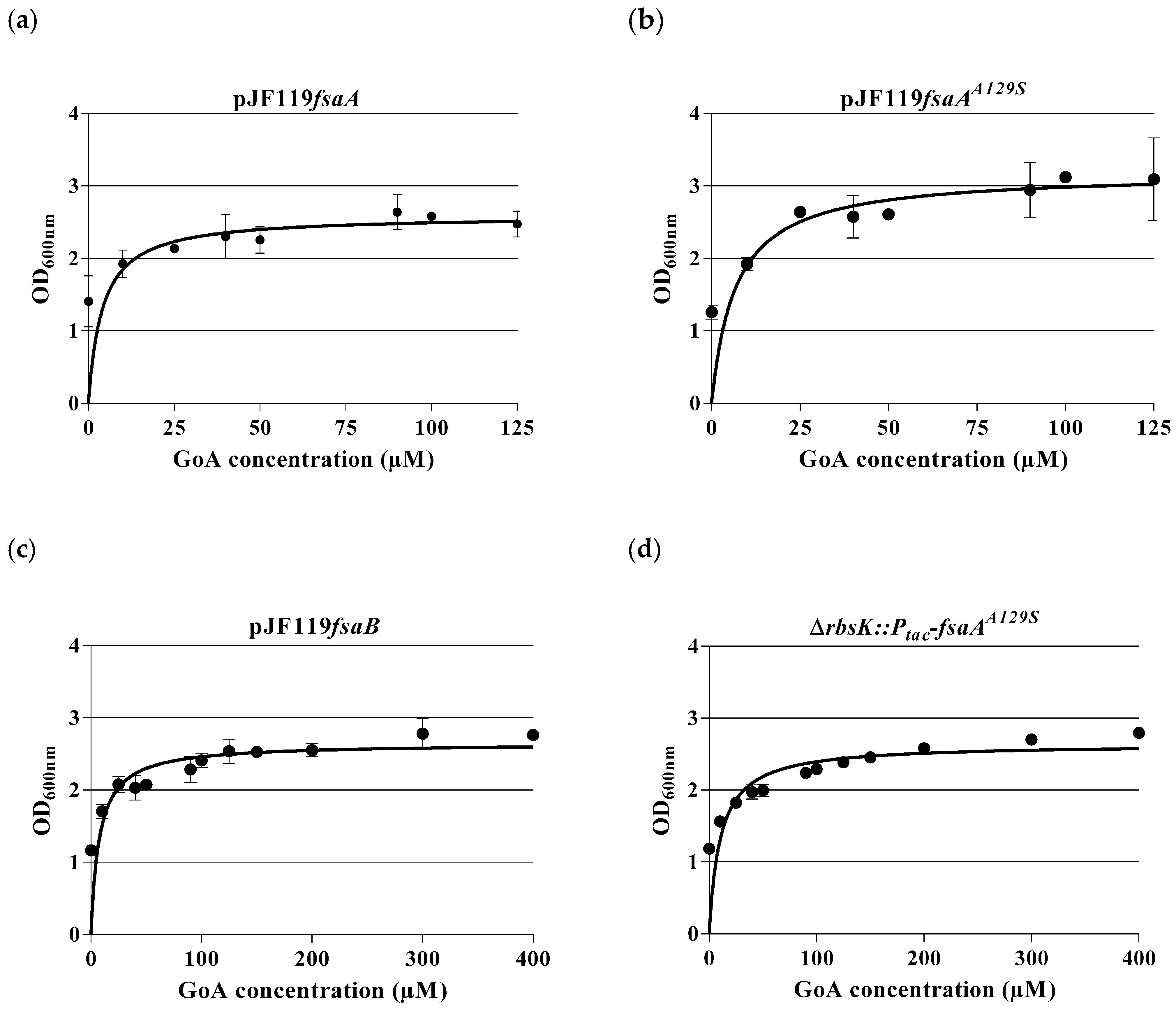
2.3. Dose–Response Curves for A5P and GoA
3. Results
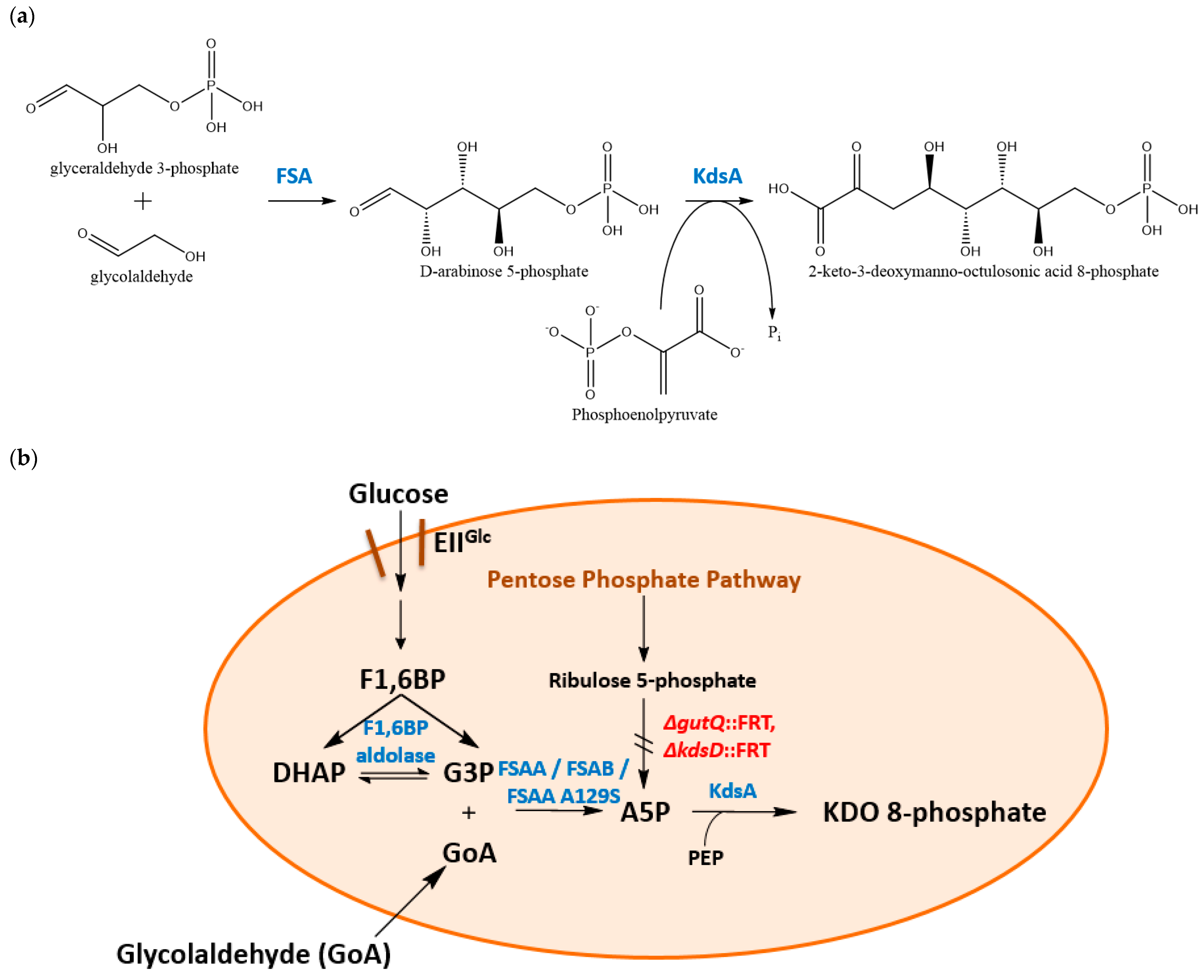
3.1. Construction and Characterization of an E. coli K-12 Double Mutant Strain Deficient in API
3.2. Physiological Observations in kdsD gutQ Double Mutants with or without FSA Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Trent, M.S. Biosynthesis and export of bacterial lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Meredith, T.C.; Aggarwal, P.; Mamat, U.; Lindner, B.; Woodard, R. Redefining the requisite lipopolysaccharide structure in Escherichia coli. ACS Chem. Biol. 2006, 1, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Cipolla, L.; Gabrielli, L.; Bini, D.; Russo, L.; Shaikh, N. Kdo: A critical monosaccharide for bacteria viability. Nat. Prod. Rep. 2010, 27, 1618–1629. [Google Scholar] [CrossRef]
- Müller-Loennies, S.; Lindner, B.; Brade, H. Structural analysis of oligosaccharides from lipopolysaccharide (LPS) of Escherichia coli K12 strain W3100 reveals a link between inner and outer core LPS biosynthesis. J. Biol. Chem. 2003, 278, 34090–34101. [Google Scholar] [CrossRef] [PubMed]
- Meredith, T.C.; Woodard, R.W. Escherichia coli YrbH is a D-arabinose 5-phosphate isomerase. J. Biol. Chem. 2003, 278, 32771–32777. [Google Scholar] [CrossRef] [PubMed]
- Meredith, T.C.; Woodard, R.W. Identification of GutQ from Escherichia coli as a D-arabinose 5-phosphate isomerase. J. Bacteriol. 2005, 187, 6936–6942. [Google Scholar] [CrossRef] [PubMed]
- Sperandeo, P.; Pozzi, C.; Dehò, G.; Polissi, A. Non-essential KDO biosynthesis and new essential cell envelope biogenesis genes in the Escherichia coli yrbG-yhbG locus. Res. Microbiol. 2006, 157, 547–558. [Google Scholar] [CrossRef]
- Schürmann, M.; Sprenger, G.A. Fructose-6-phosphate aldolase is a novel class I aldolase from Escherichia coli and is related to a novel group of bacterial transaldolases. J. Biol. Chem. 2001, 276, 11055–11061. [Google Scholar] [CrossRef]
- Samland, A.K.; Sprenger, G.A. Synthetic potential of dihydroxyacetone-utilizing aldolases. In Industrial Biocatalysis; Grunwald, P., Ed.; Pan Stanford Publishing Pte. Ltd.: Singapore, 2015; pp. 783–816. [Google Scholar]
- Garrabou, X.; Castillo, J.A.; Guérard-Hélaine, C.; Parella, T.; Joglar, J.; Lemaire, M.; Clapés, P. Asymmetric self- and cross-aldol reactions of glycolaldehyde catalyzed by D-fructose-6-phosphate aldolase. Angew. Chem. 2009, 121, 5629–5633. [Google Scholar] [CrossRef]
- Sánchez-Moreno, I.; Hélaine, V.; Poupard, N.; Charmantray, F.; Légeret, B.; Hecquet, L.; García-Junceda, E.; Wohlgemuth, R.; Guérard-Hélaine, C.; Lemaire, M. One-pot cascade reactions using fructose-6-phosphate aldolase: Efficient synthesis of D-arabinose 5-phosphate, D-fructose 6-phosphate and analogues. Adv. Synth. Catal. 2012, 354, 1725–1730. [Google Scholar] [CrossRef]
- Guérard-Hélaine, C.; Debacker, M.; Clapés, P.; Szekrenyi, A.; Hélaine, V.; Lemaire, M. Efficient biocatalytic processes for highly valuable terminally phosphorylated C5 to C9 D-ketoses. Green Chem. 2014, 16, 1109–1113. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Neidhardt, F.C.; Bloch, P.L.; Smith, D.F. Culture medium for enterobacteria. J. Bacteriol. 1974, 119, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef] [PubMed]
- Trachtmann, N. (University of Stuttgart, Stuttgart. Personal Communication, 2016.
- Fürste, J.P.; Pansegrau, W.; Frank, R.; Blöcker, H.; Scholz, P.; Bagdasarian, M.; Lanka, E. Molecular cloning of the plasmid RP4 primase region in a multi-host-range tacP expression vector. Gene 1986, 48, 119–131. [Google Scholar] [CrossRef]
- Inoue, T. Microbial Aldolases as C-C Bonding Enzymes: Investigation of Structural-Functional Characteristics and Application for Stereoselective Reactions. Ph.D. Dissertation, University of Stuttgart, Stuttgart, Germany, 2006. [Google Scholar] [CrossRef]
- Trachtmann, N.; Alvarez Fong, K.F.; Guitart Font, E.; Sprenger, G.A. Construction of chromosomally encoded lacZ and gfp reporter strains of Escherichia coli for the study of global regulation of metabolism. Eng. Life Sci. 2016, 16, 675–681. [Google Scholar] [CrossRef]
- Cherepanov, P.P.; Wackernagel, W. Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene 1995, 158, 9–14. [Google Scholar] [CrossRef]
- Guitart Font, E.; Sprenger, G.A. Opening a novel biosynthetic pathway to dihydroxyacetone and glycerol in Escherichia coli mutants through expression of a gene variant (fsaAA129S) for fructose 6-phosphate aldolase. Int. J. Mol. Sci. 2020, 21, 9625. [Google Scholar] [CrossRef]
- Eidels, L.; Rick, P.D.; Stimler, N.P.; Osborn, M.J. Transport of D-arabinose-5-phosphate and D-sedoheptulose-7-phosphate by the hexose phosphate transport system of Salmonella typhimurium. J. Bacteriol. 1974, 119, 138–143. [Google Scholar] [CrossRef]
- Kadner, R.J.; Murphy, G.P.; Stephens, C.M. Two mechanisms for growth inhibition by elevated transport of sugar phosphates in Escherichia coli. J. Gen. Microbiol. 1992, 138, 2007–2014. [Google Scholar] [CrossRef]
- Castillo, J.A.; Guérard-Hélaine, C.; Gutiérrez, M.; Garrabou, X.; Sancelme, M.; Schürmann, M.; Inoue, T.; Hélaine, V.; Charmantray, F.; Gefflaut, T.; et al. A mutant of D-fructose-6-phosphate aldolase (Ala129Ser) with improved affinity towards dihydroxyacetone for the synthesis of polyhydroxylated compounds. Adv. Synth. Catal. 2010, 352, 1039–1046. [Google Scholar] [CrossRef]
- Lachaux, C.; Frazao, C.J.R.; Kraußer, F.; Morin, N.; Walther, T.; Francois, J.M. A new synthetic pathway for the bioproduction of glycolic acid from lignocellulosic sugars aimed at maximal carbon conservation. Front. Bioeng. Biotechnol. 2019, 7, 359. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, M. Biochemische Charakterisierung und Struktur-Funktionsbeziehungen bakterieller Transaldolasen und Fruktose-6-Phosphat Aldolasen. Ph.D. Dissertation, University of Düsseldorf, Düsseldorf, Germany, 2001. (In Germany). [Google Scholar]
- King, J.R.; Woolston, B.M.; Stephanopoulos, G. Designing a new entry point into isoprenoid metabolism by exploiting fructose-6-phosphate aldolase side reactivity of Escherichia coli. ACS Synth. Biol. 2017, 6, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, M.; Schürmann, M.; Sprenger, G.A. Fructose 6-phosphate aldolase and 1-deoxy-D-xylulose 5-phosphate synthase from Escherichia coli as tools in enzymatic synthesis of 1-deoxysugars. J. Mol. Catal. B Enzym. 2002, 19–20, 247–252. [Google Scholar] [CrossRef]
- Lindner, S.N.; Aslan, S.; Müller, A.; Hoffart, E.; Behrens, P.; Edlich-Muth, C.; Blombach, B.; Bar-Even, A. A synthetic glycerol assimilation pathway demonstrates biochemical constraints of cellular metabolism. FEBS J. 2020, 287, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.; Bade, F.; Straube, E.; Rabe, K.; Frazao, C.J.R.; Walther, T. In vivo implementation of a synthetic metabolic pathway for the carbon-conserving conversion of glycolaldehyde to acetyl-CoA. Front. Bioeng. Biotechnol. 2023, 11, 1125544. [Google Scholar] [CrossRef] [PubMed]
- Yep, A.; Sorenson, R.J.; Wilson, M.R.; Showalter, H.D.H.; Larsen, S.D.; Keller, P.R.; Woodard, R.W. Enediol mimics as inhibitors of the D-arabinose 5-phosphate isomerase (KdsD) from Francisella tularensis. Bioorg. Med. Chem. Lett. 2011, 21, 2679–2682. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, L.; Merlo, S.; Airoldi, C.; Sperandeo, P.; Gianera, S.; Polissi, A.; Nicotra, F.; Holler, T.P.; Woodard, R.W.; Cipolla, L. Arabinose 5-phosphate isomerase as a target for antibacterial design: Studies with substrate analogues and inhibitors. Bioorg. Med. Chem. 2014, 22, 2576–2583. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Garrett, T.A.; Reynolds, C.M.; Shaw, W.A.; Moore, J.D.; Smith Jr, D.C.; Ribeiro, A.A.; Murphy, R.C.; Ulevitch, R.J.; Fearns, C.; et al. Kdo2-Lipid A of Escherichia coli, a defined endotoxin that activates macrophages via TLR-4. J. Lipid. Res. 2006, 47, 1097–1111. [Google Scholar] [CrossRef]
- Wang, J.; Ma, W.; Wang, Z.; Wang, X. Construction and characterization of an Escherichia coli mutant producing Kdo2-lipid A. Mar. Drugs 2014, 12, 1495–1511. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Pfirrmann, T.; Glomb, M.A. Transketolase A from E. coli significantly suppresses protein glycation by glycolaldehyde and glyoxal in vitro. J. Agric. Food Chem. 2017, 65, 8196–8202. [Google Scholar] [CrossRef] [PubMed]
- Noda-Garcia, L.; Liebermeister, W.; Tawfik, D.S. Metabolite-enzyme coevolution: From single enzymes to metabolite pathways and networks. Annu. Rev. Biochem. 2018, 87, 187–216. [Google Scholar] [CrossRef] [PubMed]
- Tani, Y.; Morita, H.; Ogata, K. Glycolaldehyde synthesizing pathway involved in vitamin B6 biosynthesis in Escherichia coli B. Agric. Biol. Chem. 1977, 41, 1749–1754. [Google Scholar] [CrossRef]
- Haussmann, C.; Rohdich, F.; Schmidt, E.; Bacher, A.; Richter, G. Biosynthesis of pteridines in Escherichia coli. J. Biol. Chem. 1998, 273, 17418–17424. [Google Scholar] [CrossRef]
- Lee, C.; Kim, I.; Lee, J.; Lee, K.-L.; Min, B.; Park, C. Transcriptional activation of the aldehyde reductase YqhD by YqhC and its implication in glyoxal metabolism of Escherichia coli K-12. J. Bacteriol. 2010, 192, 4205–4214. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, J.; Wu, Y.; Liu, D. Metabolic engineering of Corynebacterium glutamicum for the de novo production of ethylene glycol from glucose. Metab. Eng. 2016, 33, 12–18. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guitart Font, E.; Sprenger, G.A. Complementation of an Escherichia coli K-12 Mutant Strain Deficient in KDO Synthesis by Forming D-Arabinose 5-Phosphate from Glycolaldehyde with Fructose 6-Phosphate Aldolase (FSA). Appl. Microbiol. 2024, 4, 470-480. https://doi.org/10.3390/applmicrobiol4010032
Guitart Font E, Sprenger GA. Complementation of an Escherichia coli K-12 Mutant Strain Deficient in KDO Synthesis by Forming D-Arabinose 5-Phosphate from Glycolaldehyde with Fructose 6-Phosphate Aldolase (FSA). Applied Microbiology. 2024; 4(1):470-480. https://doi.org/10.3390/applmicrobiol4010032
Chicago/Turabian StyleGuitart Font, Emma, and Georg A. Sprenger. 2024. "Complementation of an Escherichia coli K-12 Mutant Strain Deficient in KDO Synthesis by Forming D-Arabinose 5-Phosphate from Glycolaldehyde with Fructose 6-Phosphate Aldolase (FSA)" Applied Microbiology 4, no. 1: 470-480. https://doi.org/10.3390/applmicrobiol4010032