
The Tale of Staphylococcus aureus Isolated from Mastitis Infections: The Effect of Antimicrobials and Bacterial Relatedness

Abstract
:1. Introduction
2. Materials and Methods
2.1. Staphylococcus aureus’s Isolation
2.2. Testing Iodine Treatments on Udder Skin with S. aureus Planktonic Cells
2.3. Iodine’s Effect on S. aureus Biofilms
2.4. Antibiotic Susceptibility Testing
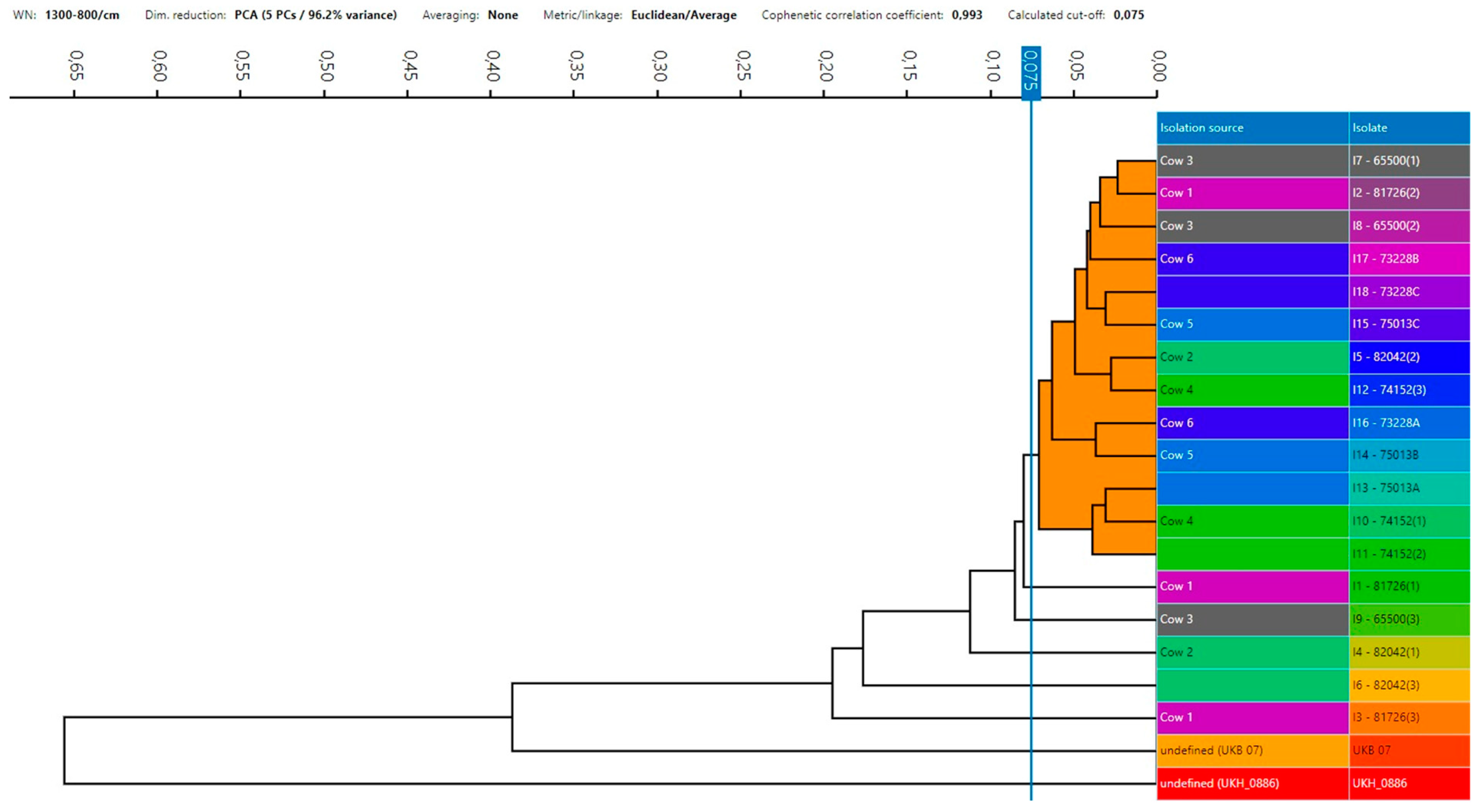
2.5. Testing S. aureus Isolates’ Relatedness Using Microbial Typing
2.6. Statistical Analysis
2.7. Ethical Statement
3. Results
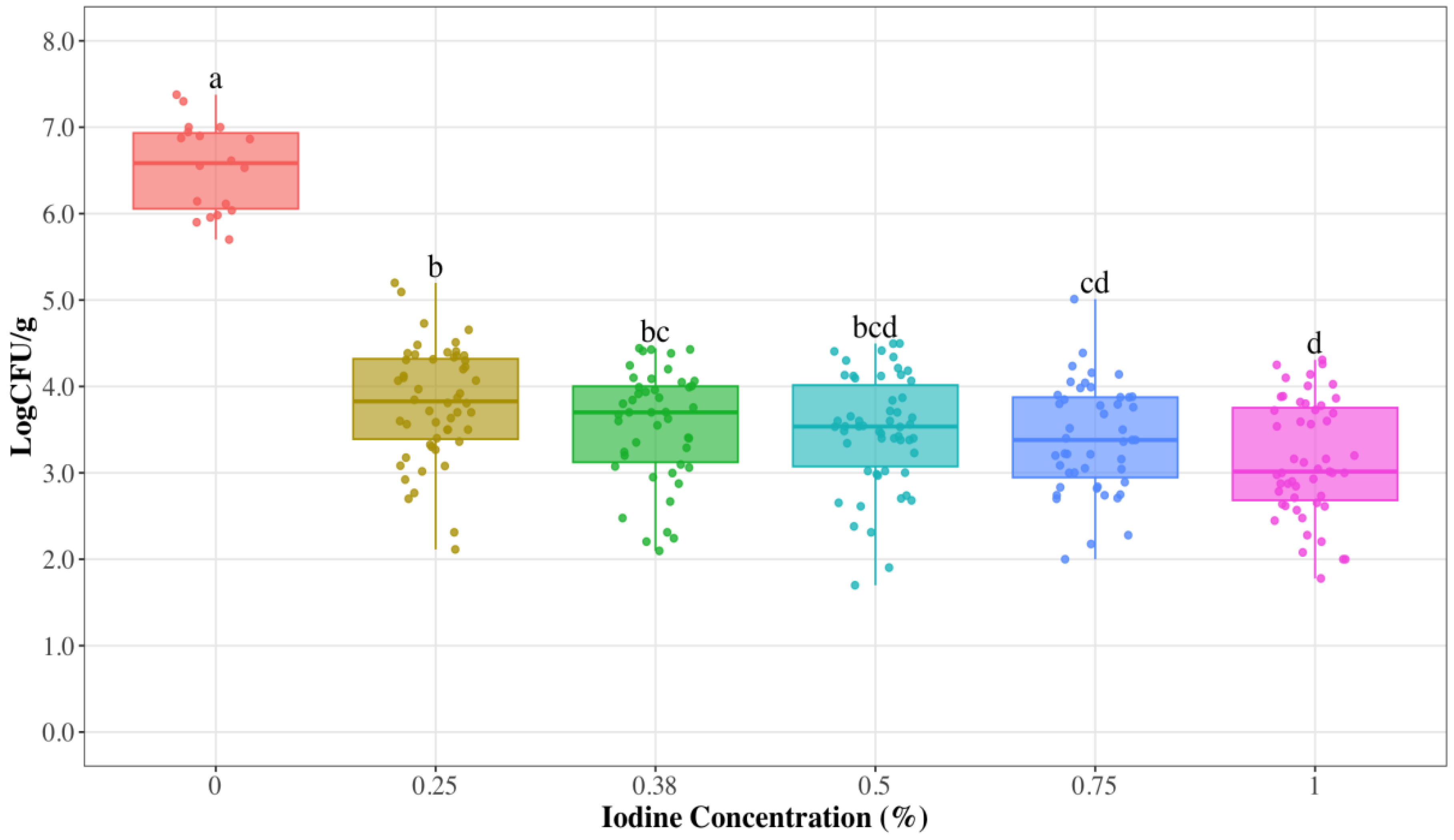
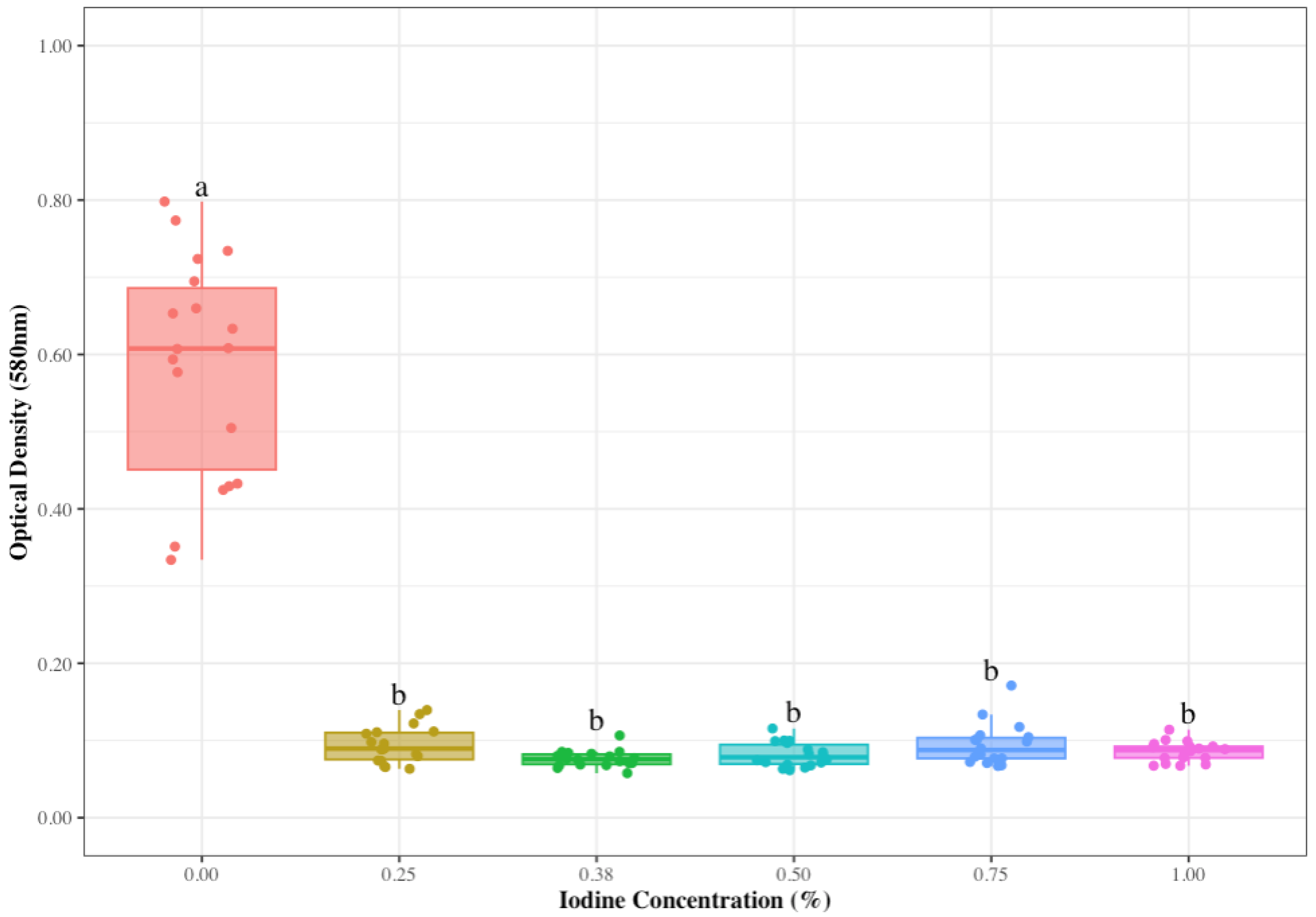
3.1. Reduced Iodine Concentrations to Control S. aureus on Udder Skin
3.2. Antimicrobial Susceptibility Testing
3.3. S. aureus Strains’ Relatedness
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seegers, H.; Fourichon, C.; Beaudeau, F. Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 2003, 34, 475–491. [Google Scholar] [CrossRef]
- Zigo, F.; Farkašová, Z.; Výrostková, J.; Regecová, I.; Ondrašovičová, S.; Vargová, M.; Sasáková, N.; Pecka-Kielb, E.; Bursová, Š.; Kiss, D.S. Dairy Cows’ Udder Pathogens and Occurrence of Virulence Factors in Staphylococci. Animals 2022, 12, 470. [Google Scholar] [CrossRef] [PubMed]
- Gogoi-Tiwari, J.; Williams, V.; Waryah, C.B.; Costantino, P.; Al-Salami, H.; Mathavan, S.; Wells, K.; Tiwari, H.K.; Hegde, N.; Isloor, S.; et al. Mammary gland pathology subsequent to acute infection with strong versus weak biofilm forming Staphylococcus aureus bovine mastitis isolates: A pilot study using non-invasive mouse mastitis model. PLoS ONE 2017, 12, e0170668. [Google Scholar] [CrossRef] [PubMed]
- Cobirka, M.; Tancin, V.; Slama, P. Epidemiology and classification of mastitis. Animals 2020, 10, 2212. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.N.; Han, S.G. Bovine mastitis: Risk factors, therapeutic strategies, and alternative treatments—A review. Asian-Australas. J. Anim. Sci. 2020, 33, 1699–1713. [Google Scholar] [CrossRef]
- Castro, V.S.; Figueiredo, E.; McAllister, T.; Stanford, K. Farm to fork impacts of super-shedders and high-event periods on food safety. Trends Food Sci. Technol. 2022, 127, 129–142. [Google Scholar] [CrossRef]
- Renault, V.; Humblet, M.-F.; Pham, P.; Saegerman, C. Biosecurity at cattle farms: Strengths, weaknesses, opportunities and threats. Pathogens 2021, 10, 1315. [Google Scholar] [CrossRef]
- Casseri, E.; Bulut, E.; Soto, S.L.; Wemette, M.; Stout, A.; Safi, A.G.; Lynch, R.; Moroni, P.; Ivanek, R. Understanding antibiotic resistance as a perceived threat towards dairy cattle through beliefs and practices: A survey-based study of dairy farmers. Antibiotics 2022, 11, 997. [Google Scholar] [CrossRef]
- Biscarini, F.; Cremonesi, P.; Castiglioni, B.; Stella, A.; Bronzo, V.; Locatelli, C.; Moroni, P. A randomized controlled trial of teat-sealant and antibiotic dry-cow treatments for mastitis prevention shows similar effect on the healthy milk microbiome. Front. Vet. Sci. 2020, 7, 581. [Google Scholar] [CrossRef]
- Ózsvári, L.; Ivanyos, D. The use of teat disinfectants and milking machine cleaning products in commercial Holstein-Friesian farms. Front. Vet. Sci. 2022, 9, 956843. [Google Scholar] [CrossRef]
- Foret, C.J.; Corbellini, C.; Young, S.; Janowicz, P. Efficacy of two iodine teat dips based on reduction of naturally occurring new intramammary infections. J. Dairy Sci. 2005, 88, 426–432. [Google Scholar] [CrossRef]
- Behiry, A.E.; Schlenker, G.; Szabo, I.; Roesler, U. In vitro susceptibility of Staphylococcus aureus strains isolated from cows with subclinical mastitis to different antimicrobial agents. J. Vet. Sci. 2012, 13, 153. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.M.M.R.; Pinheiro, E.S.C.; Gentilini, M.; Benavides, M.L.; Santos, M.V. Efficacy of a high free iodine barrier teat disinfectant for the prevention of naturally occurring new intramammary infections and clinical mastitis in dairy cows. J. Dairy Sci. 2017, 100, 3930–3939. [Google Scholar] [CrossRef] [PubMed]
- Vargová, M.; Zigo, F.; Výrostková, J.; Farkašová, Z.; Rehan, I. Biofilm-Producing Ability of Staphylococcus aureus Obtained from Surfaces and Milk of Mastitic Cows. Vet. Sci. 2023, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.S.; Okello, E.; Elashmawy, W.R.; Williams, D.R.; Anderson, R.J.; Rossitto, P.; Tonooka, K.; Glenn, K.; Karle, B.; Lehenbauer, T.W. Effectiveness of intramammary antibiotics, internal teat sealants, or both at dry-off in dairy cows: Clinical mastitis and culling outcomes. Antibiotics 2022, 11, 954. [Google Scholar] [CrossRef] [PubMed]
- Krömker, V.; Leimbach, S. Mastitis treatment-reduction in antibiotic usage in dairy cows. Reprod. Domest. Anim. 2017, 52, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Gulmez Saglam, A.; Sahin, M.; Celik, E.; Celebi, O.; Akca, D.; Otlu, S. The role of staphylococci in subclinical mastitis of cows and lytic phage isolation against to Staphylococcus aureus. Vet. World 2017, 10, 1481–1485. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, W.; Fayez, M.; Alhumam, N.; Elsohaby, I.; Quadri, S.A.; Mkrtchyan, H. The effect of staphylococcal mastitis including resistant strains on serum procalcitonin, neopterin, acute phase response and stress biomarkers in Holstein dairy cows. PeerJ 2021, 9, e11511. [Google Scholar] [CrossRef] [PubMed]
- De Vliegher, S.; Fox, L.K.; Piepers, S.; McDougall, S.; Barkema, H.W. Invited review: Mastitis in dairy heifers: Nature of the disease, potential impact, prevention, and control. J. Dairy Sci. 2012, 95, 1025–1040. [Google Scholar] [CrossRef]
- Zhao, X.; Lacasse, P. Mammary tissue damage during bovine mastitis: Causes and control1. J. Anim. Sci. 2008, 86 (Suppl. 13), 57–65. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef]
- Côté-Gravel, J.; Malouin, F. Symposium review: Features of Staphylococcus aureus mastitis pathogenesis that guide vaccine development strategies. J. Dairy Sci. 2019, 102, 4727–4740. [Google Scholar] [CrossRef]
- Fursova, K.; Sorokin, A.; Sokolov, S.; Dzhelyadin, T.; Shulcheva, I.; Shchannikova, M.; Nikanova, D.; Artem, O.; Zinovieva, N.; Brovko, F. Virulence factors and phylogeny of Staphylococcus aureus associated with bovine mastitis in russia based on genome sequences. Front. Vet. Sci. 2020, 7, 135. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. M100 Performance Standards for Antimicrobial Susceptibility Testing, 23rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2023. [Google Scholar]
- Enger, B.D.; Fox, L.K.; Gay, J.M.; Johnson, K.A. Reduction of teat skin mastitis pathogen loads: Differences between strains, dips, and contact times. J. Dairy Sci. 2015, 98, 1354–1361. [Google Scholar] [CrossRef]
- Azizoglu, R.O.; Lyman, R.; Anderson, K.L. Bovine Staphylococcus aureus: Dose response to iodine and chlorhexidine and effect of iodine challenge on antibiotic susceptibility. J. Dairy Sci. 2013, 96, 993–999. [Google Scholar] [CrossRef]
- Skowron, K.; Sękowska, A.; Kaczmarek, A.; Grudlewska, K.; Budzyńska, A.; Białucha, A.; Gospodarek-Komkowska, E. Comparison of the effectiveness of dipping agents on bacteria causing mastitis in cattle. Ann. Agric. Environ. Med. 2019, 26, 39–45. [Google Scholar] [CrossRef]
- Enger, B.D.; White, R.R.; Nickerson, S.C.; Fox, L.K. Identification of factors influencing teat dip efficacy trial results by meta-analysis. J. Dairy Sci. 2016, 99, 9900–9911. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.R.; Garvey, M.; Flynn, J.; Jordan, K.; Gleeson, D. Are some teat disinfectant formulations more effective against specific bacteria isolated on teat skin than others? Acta Vet. Scand. 2019, 61, 1–5. [Google Scholar] [CrossRef]
- Eggers, M. Infectious Disease Management and Control with Povidone Iodine. Infect. Dis. Ther. 2019, 8, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Melchior, M.B.; Vaarkamp, H.; Fink-Gremmels, J. Biofilms: A role in recurrent mastitis infections? Vet. J. 2006, 171, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Tang, X.; Dong, W.; Sun, N.; Yuan, W. A Review of Biofilm Formation of Staphylococcus aureus and Its Regulation Mechanism. Antibiotics 2022, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Salimena, A.P.S.; Lange, C.C.; Camussone, C.; Signorini, M.; Calvinho, L.F.; Brito, M.A.V.P.; Borges, C.A.V.; Guimarães, A.S.; Ribeiro, J.B.; Mendonça, L.C.; et al. Genotypic and phenotypic detection of capsular polysaccharide and biofilm formation in Staphylococcus aureus isolated from bovine milk collected from Brazilian dairy farms. Vet. Res. Commun. 2016, 40, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Darwish, S.F.; Asfour, H.A.E. Investigation of biofilm forming ability in staphylococci causing bovine mastitis using phenotypic and genotypic assays. Sci. World J. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Melchior, M.B.; Fink-Gremmels, J.; Gaastra, W. Extended antimicrobial susceptibility assay for Staphylococcus aureus isolates from bovine mastitis growing in biofilms. Vet. Microbiol. 2007, 125, 141–149. [Google Scholar] [CrossRef]
- Tremblay, Y.D.N.; Caron, V.; Blondeau, A.; Messier, S.; Jacques, M. Biofilm formation by coagulase-negative staphylococci: Impact on the efficacy of antimicrobials and disinfectants commonly used on dairy farms. Vet. Microbiol. 2014, 172, 511–518. [Google Scholar] [CrossRef]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef]
- Pyörälä, S. Treatment of mastitis during lactation. Ir. Vet. J. 2009, 62, S40. [Google Scholar] [CrossRef]
- Niemi, R.E.; Vilar, M.J.; Dohoo, I.R.; Hovinen, M.; Simojoki, H.; Rajala-Schultz, P.J. Antibiotic dry cow therapy, somatic cell count, and milk production: Retrospective analysis of the associations in dairy herd recording data using multilevel growth models. Prev. Vet. Med. 2020, 180, 105028. [Google Scholar] [CrossRef]
- Bouchard, D.S.; Rault, L.; Berkova, N.; Le Loir, Y.; Even, S. Inhibition of Staphylococcus aureus invasion into bovine mammary epithelial cells. Appl. Environ. Microbiol. 2013, 79, 877–885. [Google Scholar] [CrossRef]
- Karzis, J.; Petzer, I.-M.; Naidoo, V.; Donkin, E.F. The spread and antimicrobial resistance of Staphylococcus aureus in South African dairy herds—A review. Onderstepoort J. Vet. Res. 2021, 88, 10. [Google Scholar] [CrossRef]
- Abdi, R.D.; Gillespie, B.E.; Vaughn, J.; Merrill, C.; Headrick, S.I.; Ensermu, D.B.; D’Souza, D.H.; Agga, G.E.; Almeida, R.A.; Oliver, S.P.; et al. Antimicrobial Resistance of Staphylococcus aureus isolates from dairy cows and genetic diversity of resistant isolates. Foodborne Pathog. Dis. 2018, 15, 449–458. [Google Scholar] [CrossRef]
- Guzmán-Rodríguez, J.J.; León-Galván, M.F.; Barboza-Corona, J.E.; Valencia-Posadas, M.; Loeza-Lara, P.D.; Sánchez-Ceja, M.; Ochoa-Zarzosa, A.; López-Meza, J.E.; Gutiérrez-Chávez, A.J. Analysis of virulence traits of Staphylococcus aureus isolated from bovine mastitis in semi-intensive and family dairy farms. J. Vet. Sci. 2020, 21, e77. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Pérez, V.K.C.; Costa, G.M.D.; Guimarães, A.S.; Heinemann, M.B.; Lage, A.P.; Dorneles, E.M.S. Relationship between virulence factors and antimicrobial resistance in Staphylococcus aureus from bovine mastitis. J. Glob. Antimicrob. Resist. 2020, 22, 792–802. [Google Scholar] [CrossRef]
- Grunert, T.; Stessl, B.; Wolf, F.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M. Distinct phenotypic traits of Staphylococcus aureus are associated with persistent, contagious bovine intramammary infections. Sci. Rep. 2018, 8, 15968. [Google Scholar] [CrossRef]
- Hong, J.S.; Kim, D.; Jeong, S.H. Performance evaluation of the IR Biotyper® system for clinical microbiology: Application for detection of Staphylococcus aureus Sequence Type 8 strains. Antibiotics 2022, 11, 909. [Google Scholar] [CrossRef] [PubMed]
- Garzoni, C.; Kelley, W.L. Staphylococcus aureus: New evidence for intracellular persistence. Trends Microbiol. 2009, 17, 59–65. [Google Scholar] [CrossRef] [PubMed]
- de Jong, N.W.M.; van Kessel, K.P.M.; van Strijp, J.A.G. Immune Evasion by Staphylococcus aureus. Microbiol. Y Spectr. 2019, 7, 20. [Google Scholar]
- LeBlanc, S.J.; Lissemore, K.D.; Kelton, D.F.; Duffield, T.F.; Leslie, K.E. Major Advances in Disease Prevention in Dairy Cattle. J. Dairy Sci. 2006, 89, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, R.A.J.; Fox, L.K.; Lysnyansky, I. Mycoplasma mastitis in cattle: To cull or not to cull. Vet. J. 2016, 216, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Ganda, E.K.; Bisinotto, R.S.; Decter, D.H.; Bicalho, R.C. Evaluation of an on-farm culture system (accumast) for fast identification of milk pathogens associated with clinical mastitis in dairy cows. PLoS ONE 2016, 11, e0155314. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Breakpoints (µg/mL) | ||
|---|---|---|---|
| Susceptible (S) | Intermediate (I) | Resistant (R) | |
| Erythromycin (ERY) | ≤2 | 4 | ≥8 |
| Clindamycin (CLI) | ≤0.5 | 1–2 | ≥4 |
| Quinupristin/dalfopristin (SYN) | ≤1 | 2 | ≥4 |
| Daptomycin (DAP) | ≤1 | ||
| Vancomycin (VAN) | ≤2 | 4–8 | ≥16 |
| Tetracycline (TET) | ≤4 | 8 | ≥16 |
| Ampicillin (AMP) 1 | N/A | ||
| Gentamicin (GEN) | ≤4 | 8 | ≥16 |
| Levofloxacin (LEVO) | ≤1 | 2 | ≥4 |
| Linezolid (LZD) | ≤4 | ≥8 | |
| Ceftriaxone (AXO) 1 | N/A | ||
| Streptomycin (STR) 1 | N/A | ||
| Penicillin (PEN) | ≤0.12 | ≥0.25 | |
| Rifampin (RIF) | ≤1 | 2 | ≥4 |
| Gatifloxacin (GAT) | ≤0.5 | 1 | ≥2 |
| Ciprofloxacin (CIP) | ≤1 | ≥4 | |
| Trimethoprim/sulfamethoxazole (SXT) | ≤2/38 | ≥4/76 | |
| Oxacillin + 2%NaCl (OXA+) | ≤2 | ≥4 | |
| Iodine Concentration (%) | N | S. aureus Concentration Log CFU/g ± SE 1 | Average Reduction Log CFU/g |
|---|---|---|---|
| 0.25 | 52 | 3.81 ± 0.09 | 2.73 |
| 0.38 | 46 | 3.55 ± 0.09 | 2.99 |
| 0.5 | 54 | 3.48 ± 0.09 | 3.06 |
| 0.75 | 47 | 3.39 ± 0.09 | 3.15 |
| 1 | 51 | 3.16 ± 0.09 | 3.38 |
| Iodine Concentration (%) | N | Biofilm Concentration (OD ± SE 1) | Biofilm Formation Odds Ratio |
|---|---|---|---|
| 0.25 | 18 | 0.09 ± 0.01 | 5.78 |
| 0.38 | 18 | 0.08 ± 0.00 | 6.50 |
| 0.50 | 18 | 0.08 ± 0.00 | 6.50 |
| 0.75 | 18 | 0.09 ± 0.01 | 5.78 |
| 1.00 | 18 | 0.09 ± 0.00 | 5.78 |
| Antibiotic | Exact (Clopper–Pearson) | Minimum Inhibitory Concentration MIC (μg/mL) Distribution (n = 30) * | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R (%) | 95% CI | 0.5/9.5 | 1/19 | 2/38 | 4/76 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 500 | 1000 | |
| ERY | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | |||||||||||||
| CLI | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | |||||||||||||
| SYN | 0 | 0–0.1157033 | 1 | 29 | 0 | 0 | 0 | 0 | ||||||||||||
| DAP | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | 0 | ||||||||||||
| VAN | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||||||||||
| TET | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | ||||||||||||||
| AMP | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||||||||||
| GEN | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | |||||||||||||
| LEVO | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | 0 | ||||||||||||
| LZD | 0 | 0–0.1157033 | 0 | 0 | 30 | 0 | 0 | 0 | ||||||||||||
| AXO | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | ||||||||||||||
| STR (N/A) | 0 | |||||||||||||||||||
| PEN | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||||||||||
| RIF | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | ||||||||||||||
| GAT | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | ||||||||||||||
| CIP | 0 | 0–0.1157033 | 30 | 0 | 0 | |||||||||||||||
| STX | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | ||||||||||||||
| OXA+ | 0 | 0–0.1157033 | 30 | 0 | 0 | 0 | 0 | 0 | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perdomo, A.; Salazar, M.; Janardhanan, R.; Calle, A. The Tale of Staphylococcus aureus Isolated from Mastitis Infections: The Effect of Antimicrobials and Bacterial Relatedness. Appl. Microbiol. 2024, 4, 496-509. https://doi.org/10.3390/applmicrobiol4010035
Perdomo A, Salazar M, Janardhanan R, Calle A. The Tale of Staphylococcus aureus Isolated from Mastitis Infections: The Effect of Antimicrobials and Bacterial Relatedness. Applied Microbiology. 2024; 4(1):496-509. https://doi.org/10.3390/applmicrobiol4010035
Chicago/Turabian StylePerdomo, Angela, Maria Salazar, Rasmi Janardhanan, and Alexandra Calle. 2024. "The Tale of Staphylococcus aureus Isolated from Mastitis Infections: The Effect of Antimicrobials and Bacterial Relatedness" Applied Microbiology 4, no. 1: 496-509. https://doi.org/10.3390/applmicrobiol4010035