
Thermal Inactivation of the Heat-Resistant Pathogens Salmonella Senftenberg 775W and Escherichia coli AW1.7 in Whey Concentrate
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultivation Conditions
2.2. Whey Concentrate
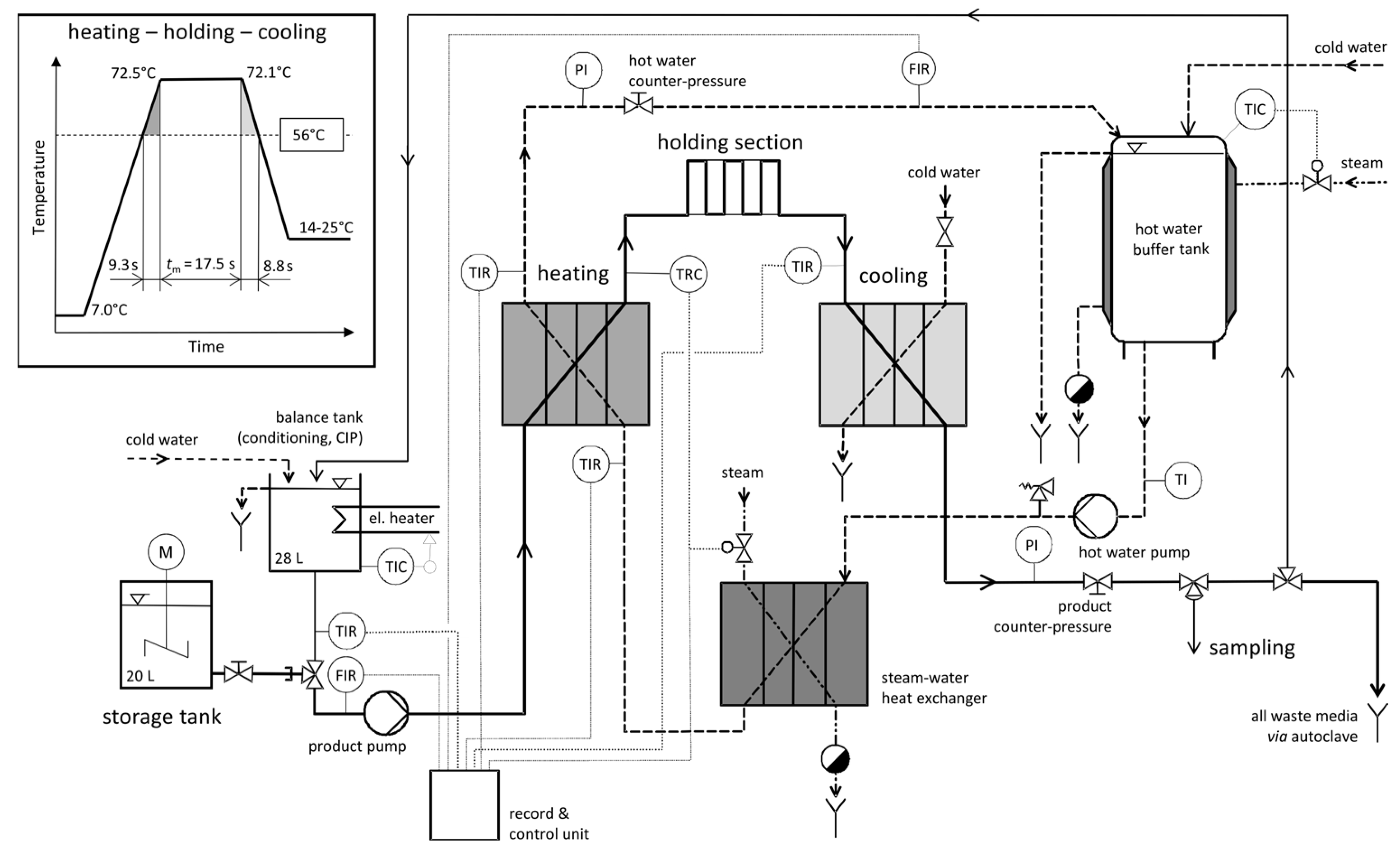
2.3. Pilot Plant Pasteurization and Heat Inactivation Studies
2.4. Detection of Survivors and Calculation of Reduction
2.5. Statistical Evaluation
3. Results
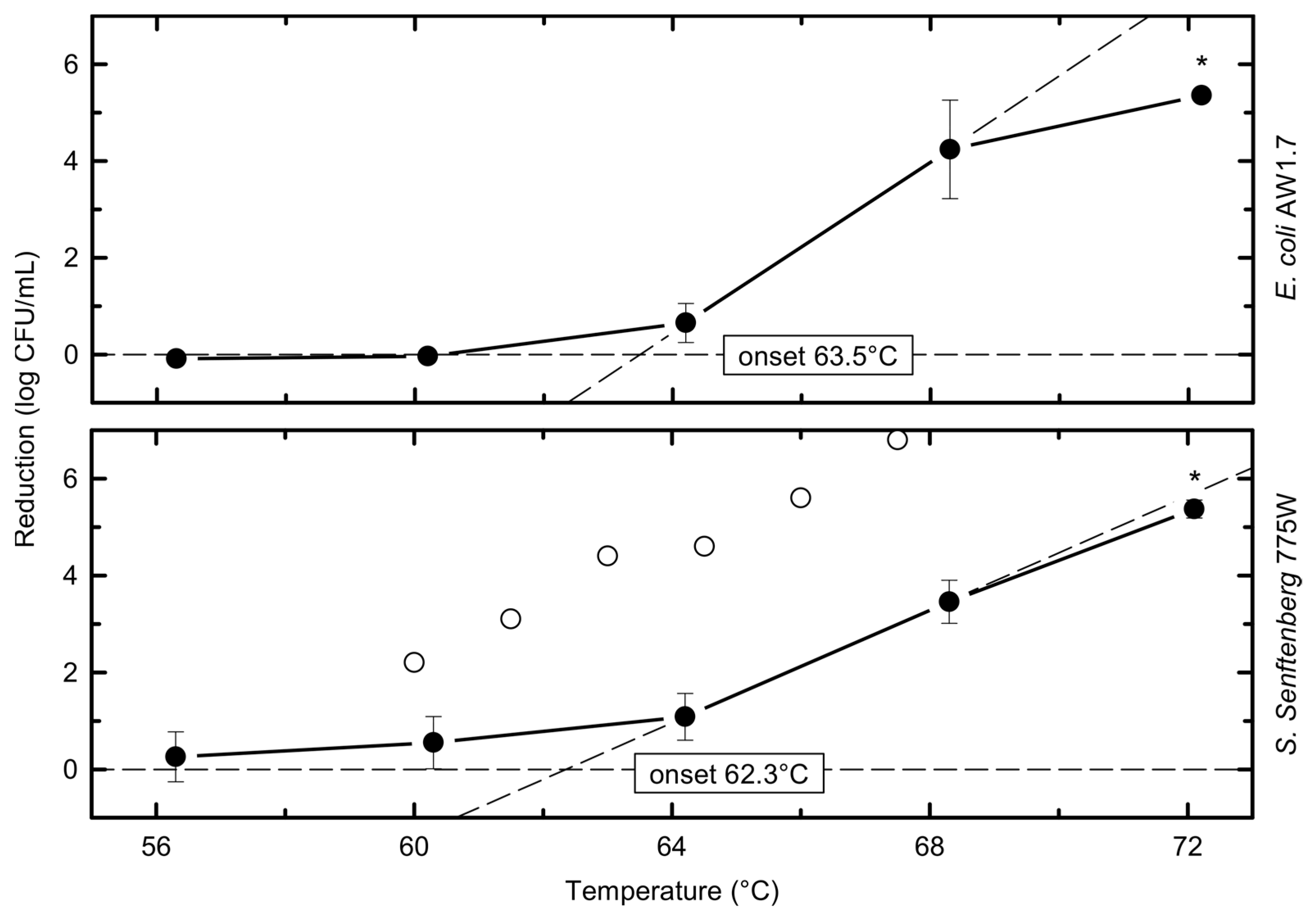
Bacterial Inactivation by Temperature
4. Discussion
4.1. Inactivation of Highly Heat-Resistant Salmonella and E. coli
4.2. Impact of the Heating System and the Matrix Whey Concentrate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kells, H.R.; Lear, S.A. Thermal death time curve of Mycobacterium tuberculosis var bovis in artificially infected milk. Appl. Microbiol. 1960, 8, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.E. Bacterial Diseases—The Impact of Milk Processing to Reduce Risks; Bulletin of the International Dairy Federation; International Dairy Federation: Brussels, Belgium, 2002; pp. 20–25. [Google Scholar]
- Pearce, L.E.; Smythe, B.W.; Crawford, R.A.; Oakley, E.; Hathaway, S.C.; Shepherd, J.M. Pasteurization of milk: The heat inactivation kinetics of milk-borne dairy pathogens under commercial-type conditions of turbulent flow. J. Dairy Sci. 2012, 95, 20–35. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S. Microbiological considerations: Pasteurized milk. Int. J. Dairy Sci. 2015, 10, 206–218. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services/Public Health Service/Food and Drug Administration. Grade “A” Pasteurized Milk Ordinance, Including Provisions from the Grade “A” Condensed and Dry Milk Products and Condensed and Dry Whey: Supplement I to the Grade “A” Pasteurized Milk Ordinance. Available online: https://www.fda.gov/media/140394 (accessed on 9 March 2024).
- CAC/RCP 57–2004; Codex Alimentarius. Code of Hygienic Practice for Milk and Milk Products. Food and Agriculture Organization: Rome, Italy, 2004.
- Hudson, J.; Wong, T.; Lake, R. Pasteurisation of Dairy Products: Times, Temperatures and Evidence for Control of Pathogens. Available online: http://www.foodsafety.govt.nz/elibrary/industry/Pasteurisation_Dairy-Science_Research.pdf (accessed on 16 May 2023).
- Kornacki, J.L.; Marth, E.H. Thermal inactivation of Salmonella senftenberg and Micrococcus freudenreichii in retentates from ultrafiltered milks. LWT—Food Sci. Technol. 1993, 26, 21–27. [Google Scholar] [CrossRef]
- Dega, C.A.; Goepfert, J.M.; Amundson, C.H. Heat resistance of salmonellae in concentrated milk. Appl. Microbiol. 1972, 23, 415–420. [Google Scholar] [CrossRef]
- Beuchat, L.R.; Komitopoulou, E.; Beckers, H.; Betts, R.P.; Bourdichon, F.; Fanning, S.; Joosten, H.M.; Kuile, B.H. Low-water activity foods: Increased concern as vehicles of foodborne pathogens. J. Food Prot. 2013, 76, 150–172. [Google Scholar] [CrossRef]
- Santillana Farakos, S.M.; Hicks, J.W.; Frank, J.F. Temperature resistance of Salmonella in low-water activity whey protein powder as influenced by salt content. J. Food Prot. 2014, 77, 631–634. [Google Scholar] [CrossRef]
- European centre for disease prevention and control, European food safety authority. Multi-country outbreak of monophasic Salmonella Typhimurium sequence type (ST) 34 linked to chocolate products—12 April 2022. oint ECDC-EFSA rapid outbreak assessment. EFS3 2022, 19, 7318E. [Google Scholar] [CrossRef]
- Podolak, R.; Enache, E.; Stone, W.; Black, D.G.; Elliott, P.H. Sources and risk factors for contamination, survival, persistence, and heat resistance of Salmonella in low-moisture foods. J. Food Prot. 2010, 73, 1919–1936. [Google Scholar] [CrossRef]
- Mañas, P.; Pagán, R.; Sala, F.J.; Condón, S. Low molecular weight milk whey components protect Salmonella senftenberg 775W against heat by a mechanism involving divalent cations. J. Appl. Microbiol. 2001, 91, 871–877. [Google Scholar] [CrossRef]
- Ng, H.; Bayne, H.G.; Garibaldi, J.A. Heat Resistance of Salmonella: The Uniqueness of Salmonella senftenberg 775W. Appl. Microbiol. 1969, 17, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Stephan, R.; Hummerjohann, J.; Blanco, J.; Zweifel, C. In vitro characterization of Shiga toxin-producing and generic Escherichia coli in respect of cheese-production relevant stresses. Arch. Leb. J. Food Saf. Food Qual. 2012, 63, 136–141. [Google Scholar] [CrossRef]
- Dlusskaya, E.A.; McMullen, L.M.; Gänzle, M.G. Characterization of an extremely heat-resistant Escherichia coli obtained from a beef processing facility. J. Appl. Microbiol. 2011, 110, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Mercer, R.G.; Zheng, J.; Garcia-Hernandez, R.; Ruan, L.; Gänzle, M.G.; McMullen, L.M. Genetic determinants of heat resistance in Escherichia coli. Front. Microbiol. 2015, 6, 932. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.V.; Harhay, G.P.; Bono, J.L.; Smith, T.P.L.; Harhay, D.M. Genome sequence of the thermotolerant foodborne pathogen Salmonella enterica Serovar Senftenberg ATCC 43845 and phylogenetic analysis of loci encoding increased protein quality control mechanisms. mSystems 2017, 2, e00190-16. [Google Scholar] [CrossRef] [PubMed]
- Guragain, M.; Bono, J.L.; Bosilevac, J.M. Complete closed genome sequence of the extremely heat-resistant strain Escherichia coli AW1.7. Microbiol. Resour. Announc. 2021, 10, e0050221. [Google Scholar] [CrossRef]
- Hammer, P.; Kiesner, C.; Walte, H.; Knappstein, K.; Teufel, P. Heat resistance of Mycobacterium avium ssp. paratuberculosis in raw milk tested in a pilot-plant pasteurizer. Kiel. Milchwirtsch. Forschungsberichte 2002, 54, 275–303. [Google Scholar]
- Peng, S.; Hummerjohann, J.; Stephan, R.; Hammer, P. Short communication: Heat resistance of Escherichia coli strains in raw milk at different subpasteurization conditions. J. Dairy Sci. 2013, 96, 3543–3546. [Google Scholar] [CrossRef]
- D’Aoust, J.-Y.; Emmons, D.B.; McKELLAR, R.; Timbers, G.E.; Todd, E.C.D.; Sewell, A.M.; Warburton, D.W. Thermal Inactivation of Salmonella Species in Fluid Milk 1. J. Food Prot. 1987, 50, 494–501. [Google Scholar] [CrossRef]
- Doyle, M.E.; Mazzotta, A.S. Review of studies on the thermal resistance of Salmonellae. J. Food Prot. 2000, 63, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.B.; Bradshaw, J.G.; Peeler, J.T. Thermal resistance of egg-associated epidemic strains of Salmonella enteritidis. J. Food Sci. 1991, 56, 391–393. [Google Scholar] [CrossRef]
- Stoeckel, M.; Abduh, S.B.M.; Atamer, Z.; Hinrichs, J. Inactivation of Bacillus spores in batch vs continuous heating systems at sterilisation temperatures. Int. J. Dairy Technol. 2014, 67, 334–341. [Google Scholar] [CrossRef]
- Liu, Y.; Gill, A.; McMullen, L.; Gänzle, M.G. Variation in heat and pressure resistance of verotoxigenic and nontoxigenic Escherichia coli. J. Food Prot. 2015, 78, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Solowey, M.; Sutton, R.R.; Calesnick, E.J. Heat resistance of Salmonella organisms isolated from spray-dried whole-egg powder. Food Technol. 1948, 2, 9–14. [Google Scholar]



{kind=link}
{kind=link}
| Microorganism | Strain | Serovar | MLST | Genome Reference |
|---|---|---|---|---|
| Salmonella enterica subsp. enterica | 775W (DSM 10062) | Senftenberg | ST185 | [19] |
| Escherichia coli | AW1.7 (DSM 108612) | O128ac:H12 | ST6002 | [20] |
| Escherichia coli AW1.7 | Salmonella Senftenberg 775W | ||||||
|---|---|---|---|---|---|---|---|
| Temperature (°C) | log10 Count (Mean ± SD) | D-Value | Temperature (°C) | log10 Count (Mean ± SD) | D-Value | ||
| Initial (N0) | Reduction * | (min) | Initial (N0) | Reduction | (min) | ||
| 56.3 ± 0.1 | 5.4 ± 0.1 | −0.1 ± 0.0 | - | 56.3 ± 0.2 | 5.5 ± 0.2 | 0.3 ± 0.5 | 1.12 |
| 60.2 ± 0.1 | 0.0 ± 0.1 | - | 60.3 ± 0.1 | 0.6 ± 0.5 | 0.53 | ||
| 64.2 ± 0.1 | 0.7 ± 0.4 | 0.45 | 64.2 ± 0.1 | 1.1 ± 0.5 | 0.27 | ||
| 68.3 ± 0.1 | 4.2 ± 1.0 | 0.07 | 68.3 ± 0.2 | 3.5 ± 0.4 | 0.08 | ||
| 72.2 ± 0.2 | 5.4 ± 0.1 | 0.05 | 72.1 ± 0.4 | 5.4 ± 0.2 | 0.05 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiedler, G.; Nöbel, S.; Matzen, S.; Samtlebe, M.; Franz, C.M.A.P. Thermal Inactivation of the Heat-Resistant Pathogens Salmonella Senftenberg 775W and Escherichia coli AW1.7 in Whey Concentrate. Appl. Microbiol. 2024, 4, 510-519. https://doi.org/10.3390/applmicrobiol4010036
Fiedler G, Nöbel S, Matzen S, Samtlebe M, Franz CMAP. Thermal Inactivation of the Heat-Resistant Pathogens Salmonella Senftenberg 775W and Escherichia coli AW1.7 in Whey Concentrate. Applied Microbiology. 2024; 4(1):510-519. https://doi.org/10.3390/applmicrobiol4010036
Chicago/Turabian StyleFiedler, Gregor, Stefan Nöbel, Sönke Matzen, Meike Samtlebe, and Charles M. A. P. Franz. 2024. "Thermal Inactivation of the Heat-Resistant Pathogens Salmonella Senftenberg 775W and Escherichia coli AW1.7 in Whey Concentrate" Applied Microbiology 4, no. 1: 510-519. https://doi.org/10.3390/applmicrobiol4010036