
PluMu—A Mu-like Bacteriophage Infecting Actinobacillus pleuropneumoniae
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Genomic Comparisons
2.3. Precipitation of Viral Supernatants
2.4. PluMu Production
2.5. Transmission Electron Microscopy (TEM)
3. Results
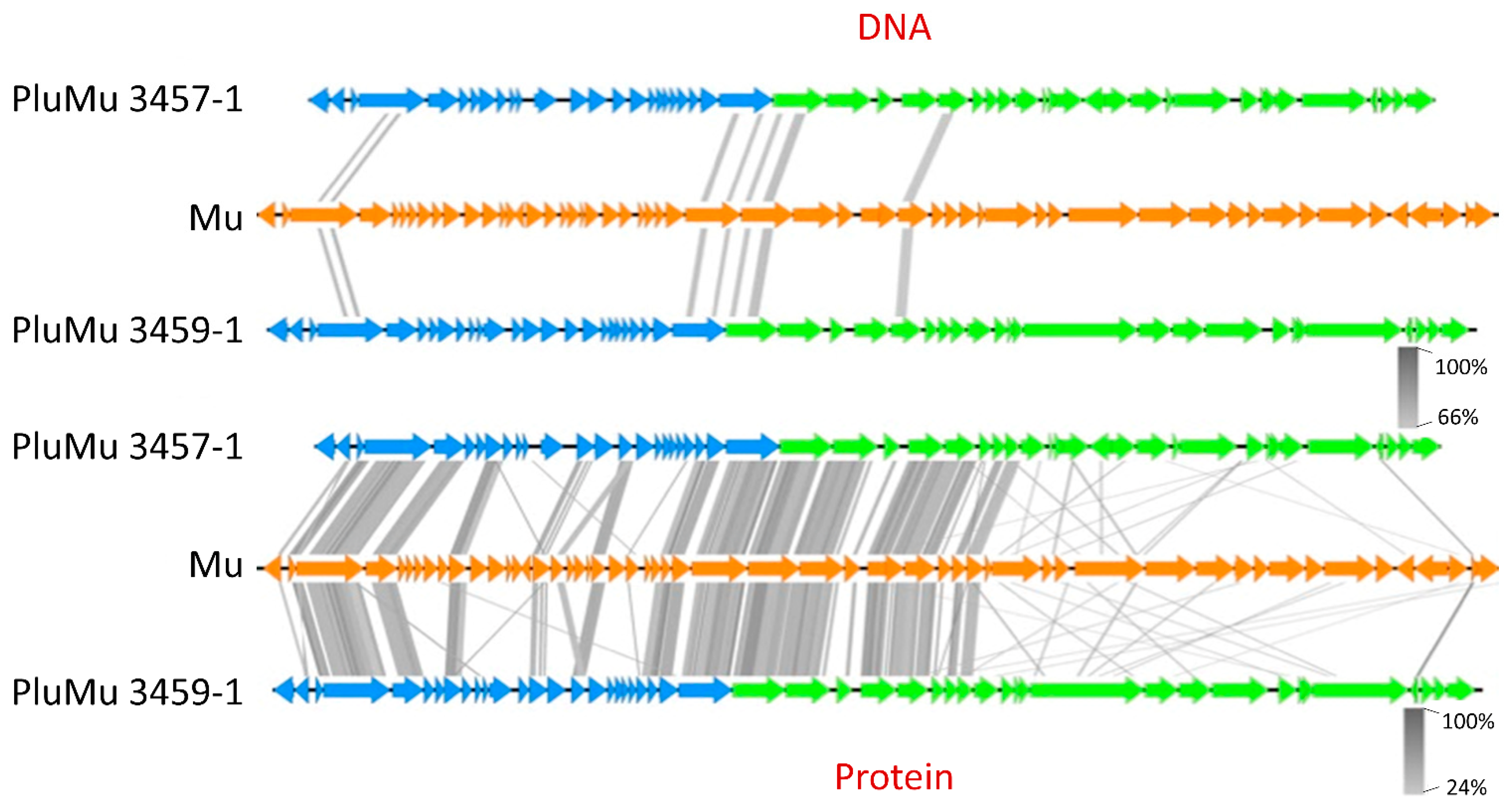
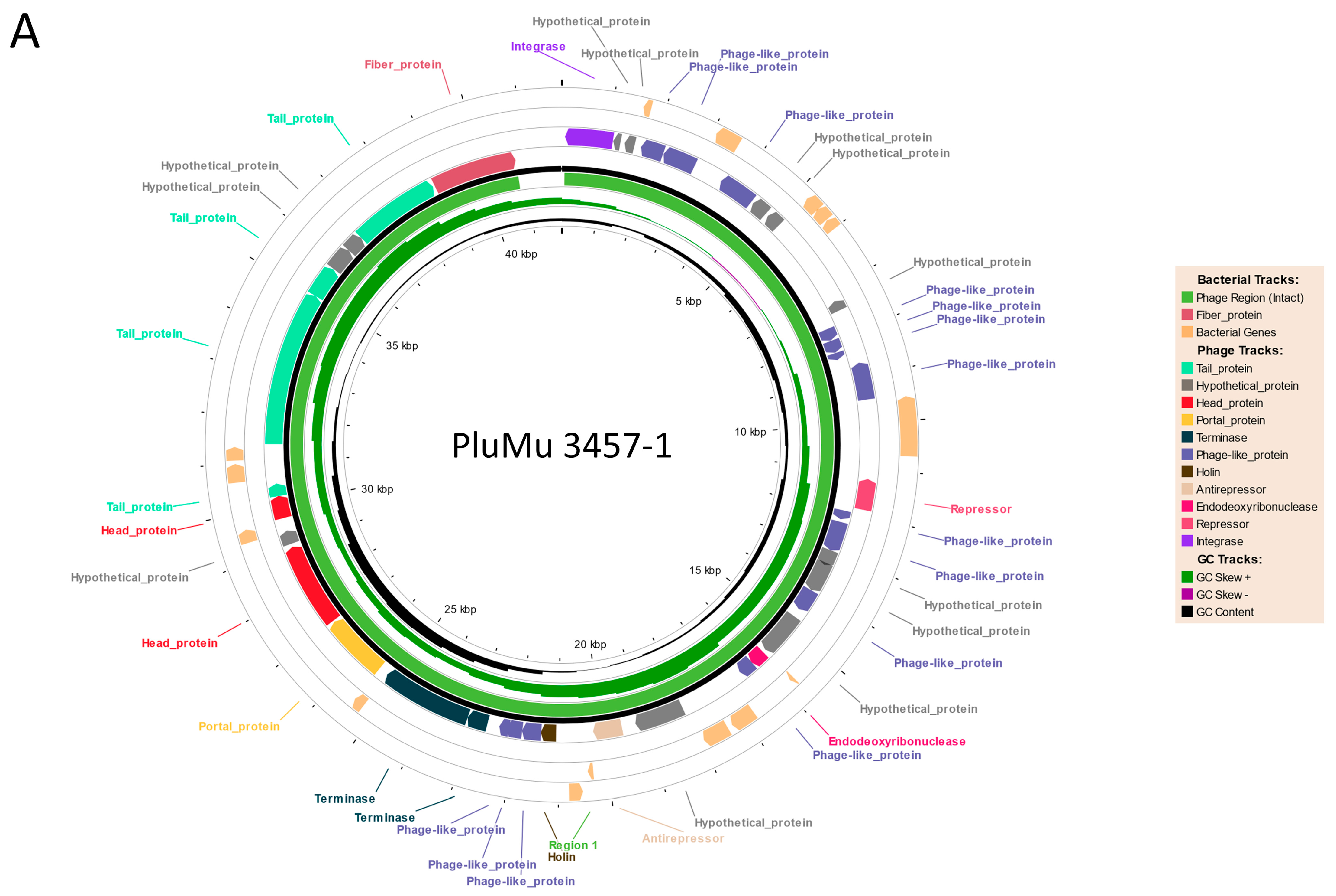
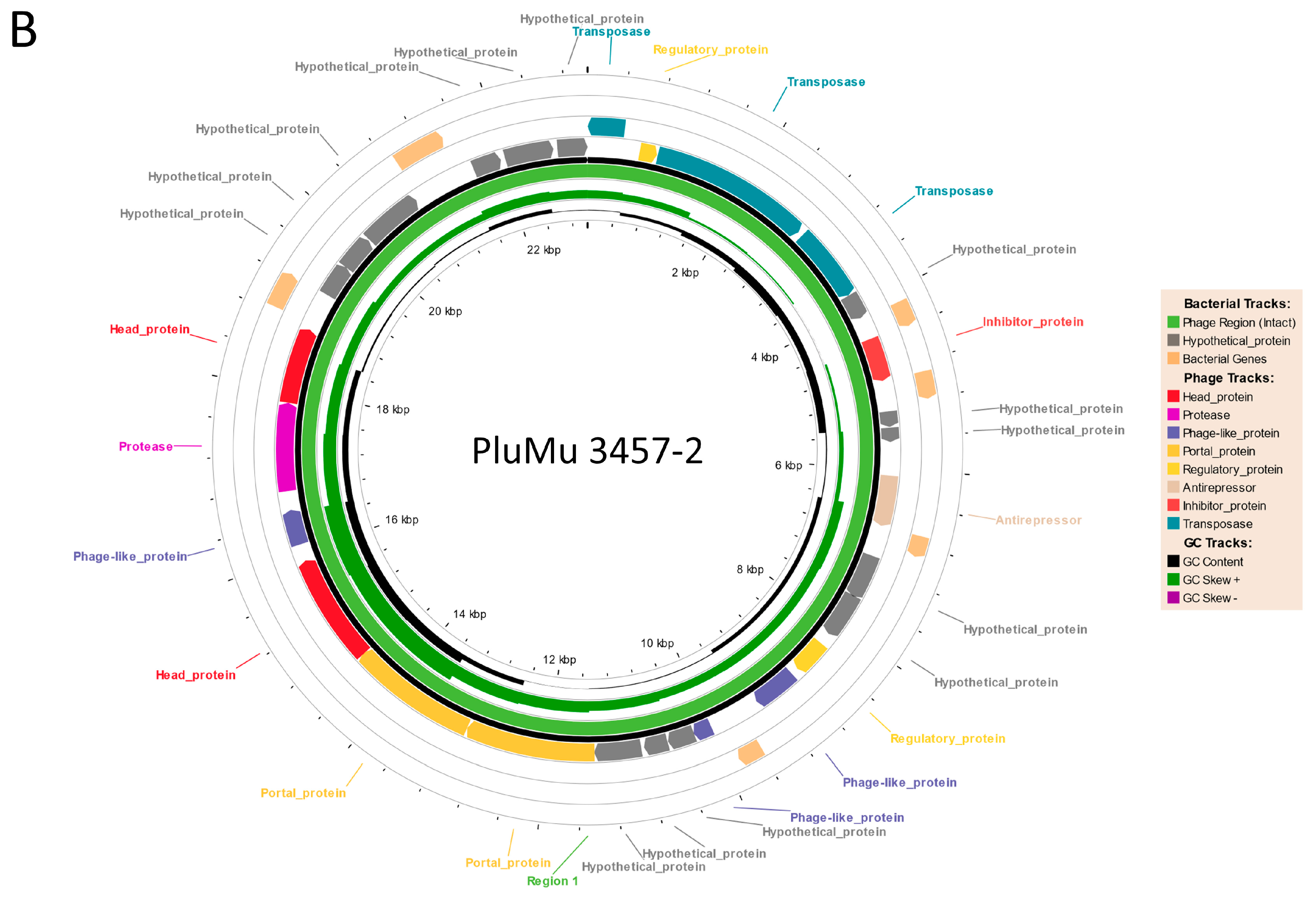
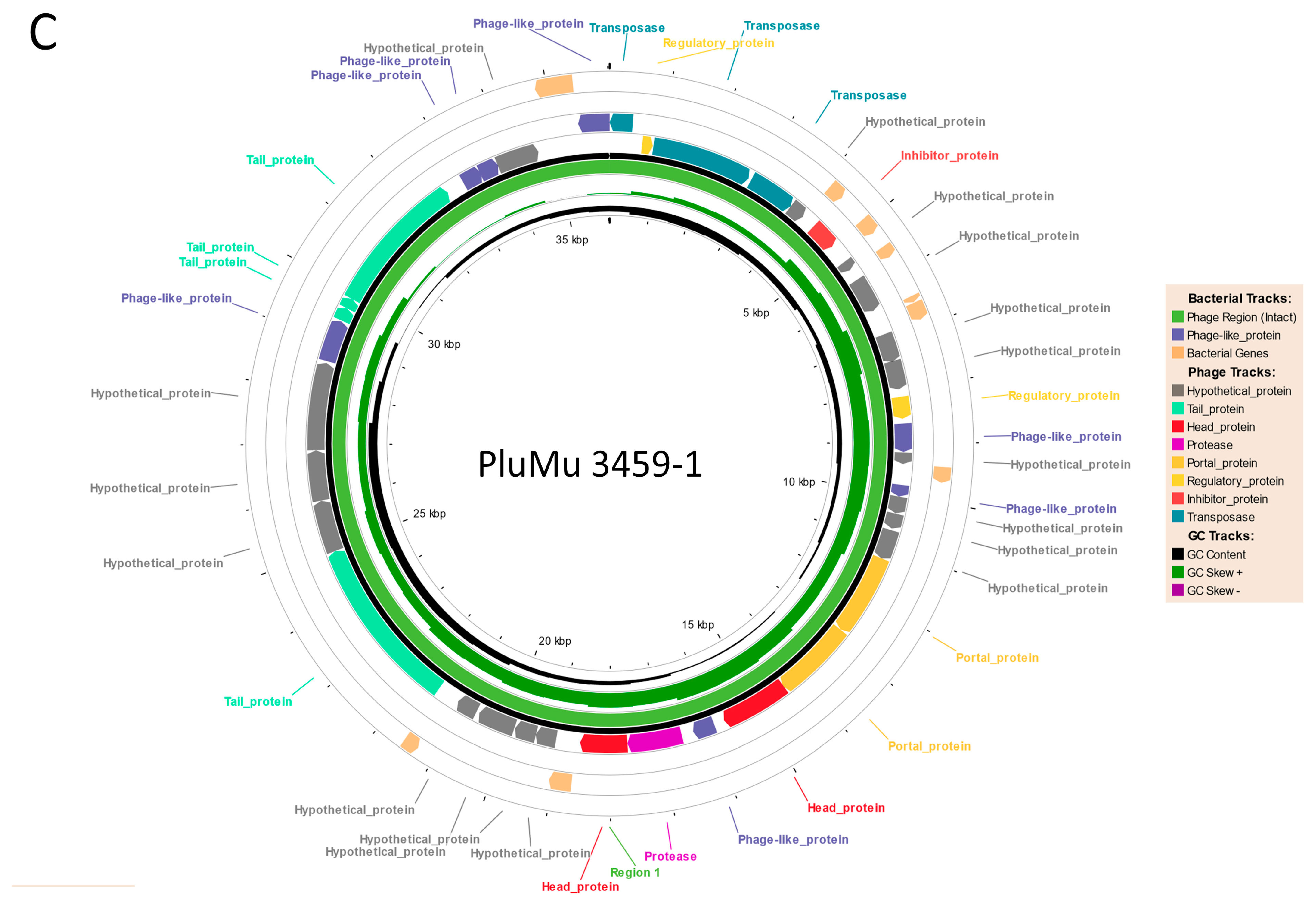
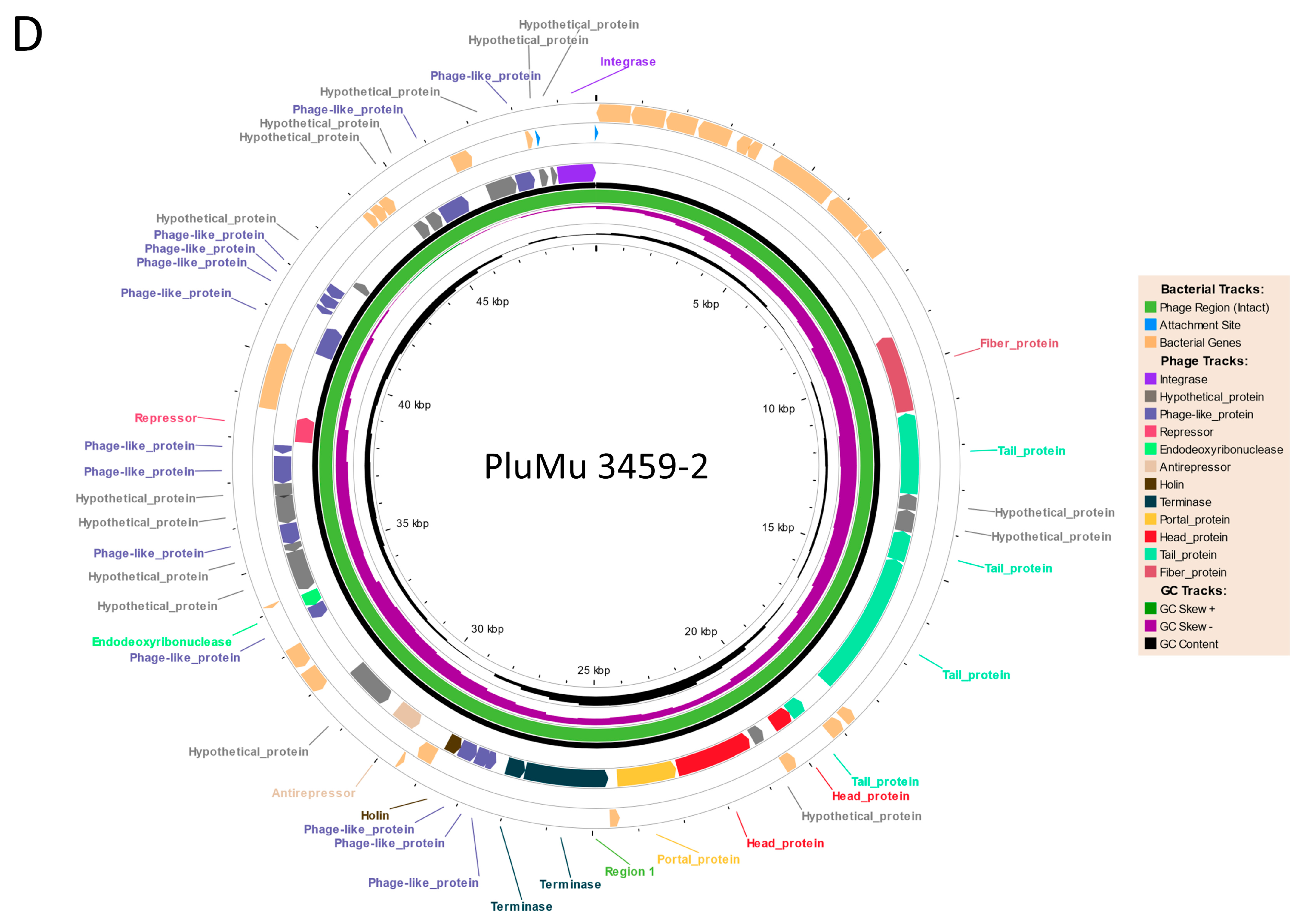
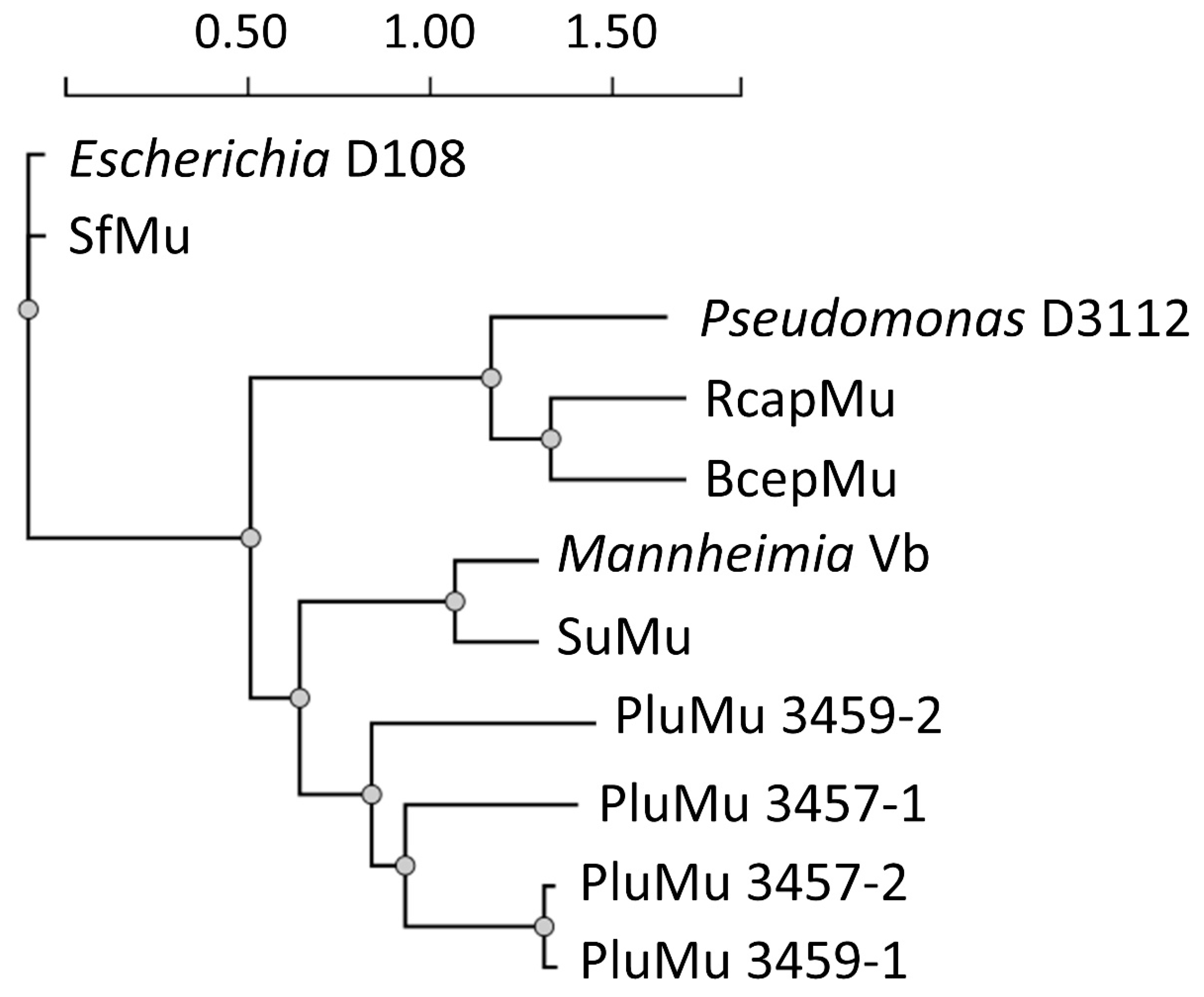
3.1. Genomic Comparisons
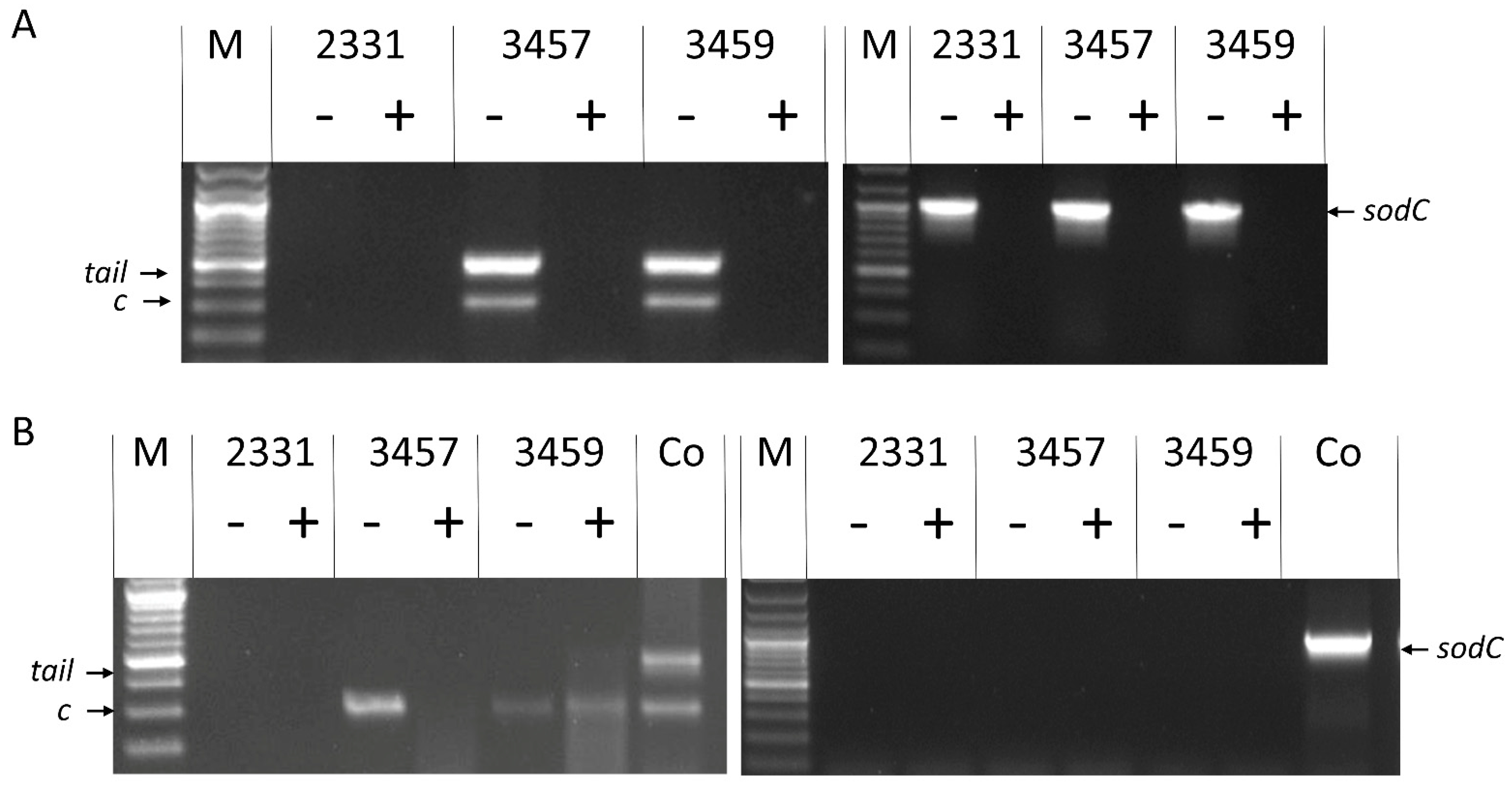
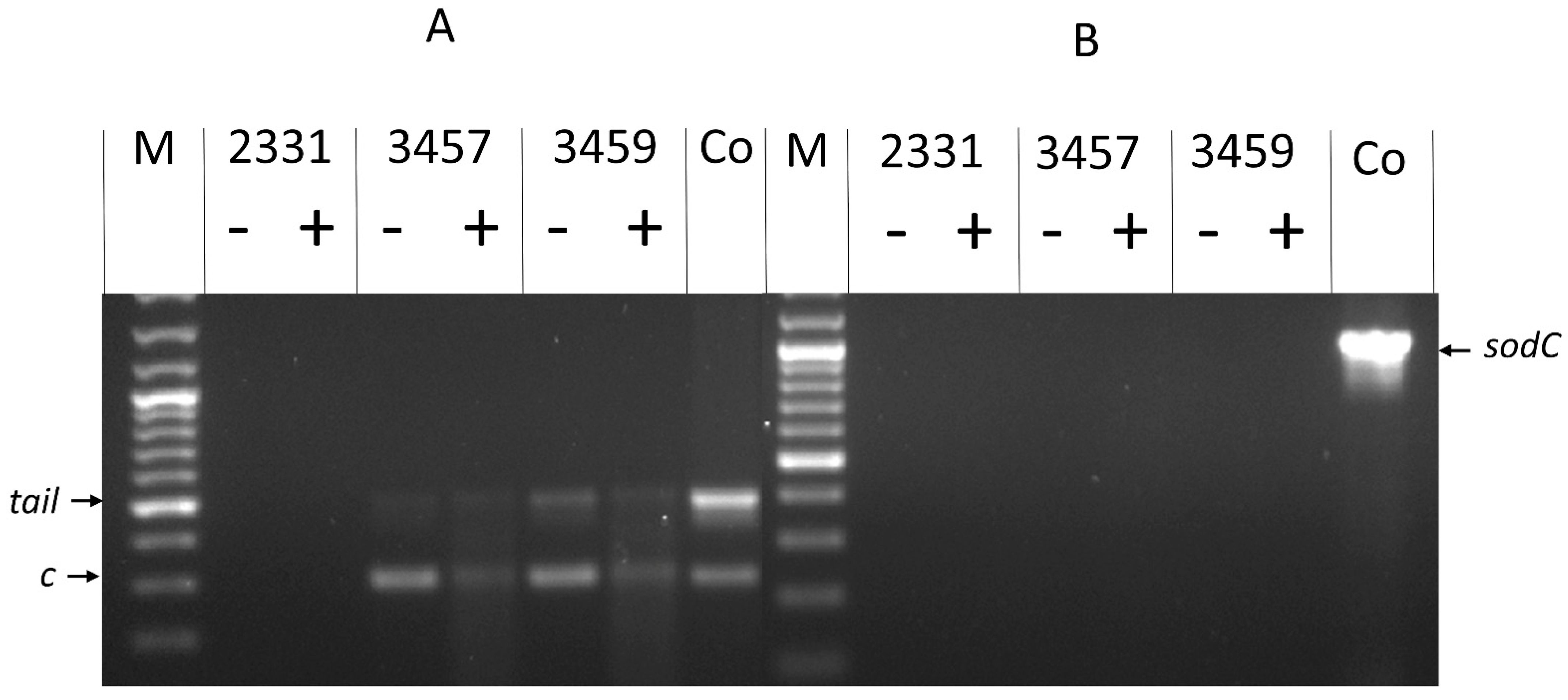
3.2. Molecular Analyses Indicate That 3457 and 3459 Produce Active Phage
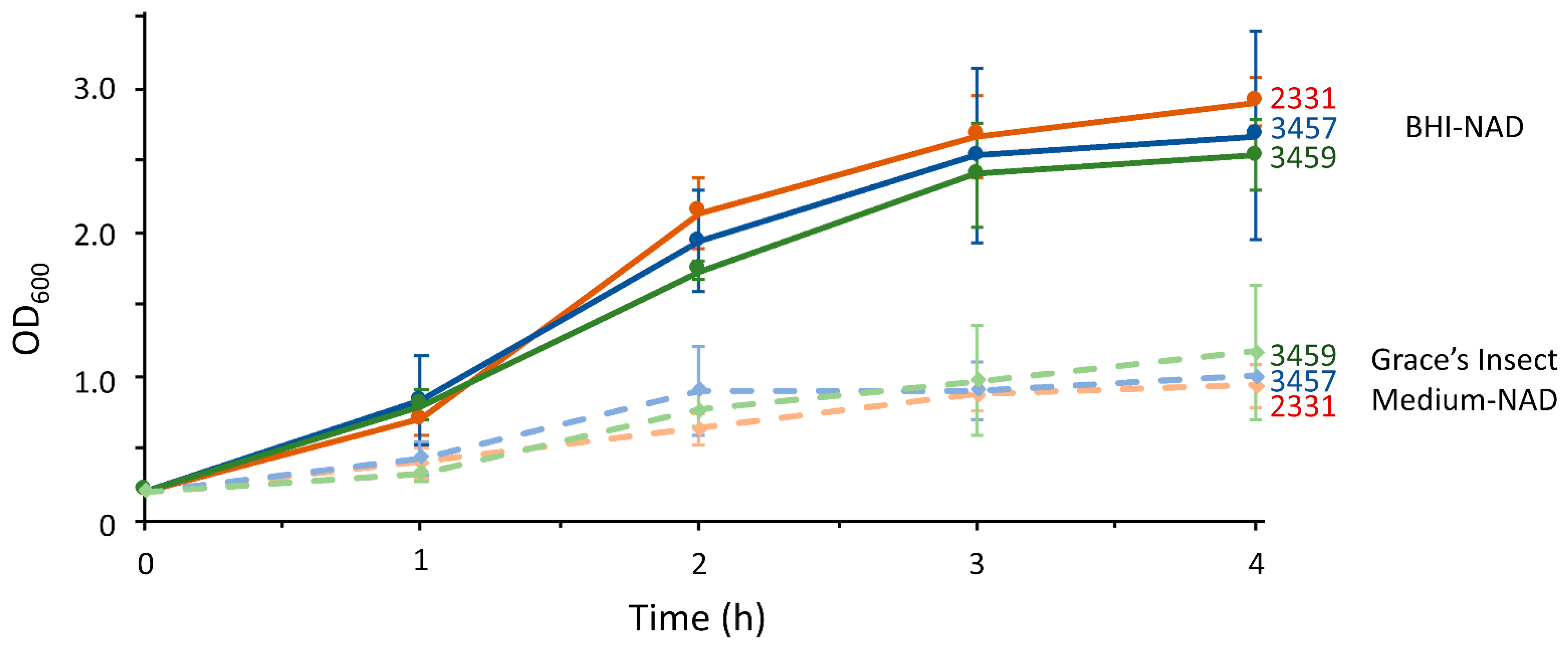
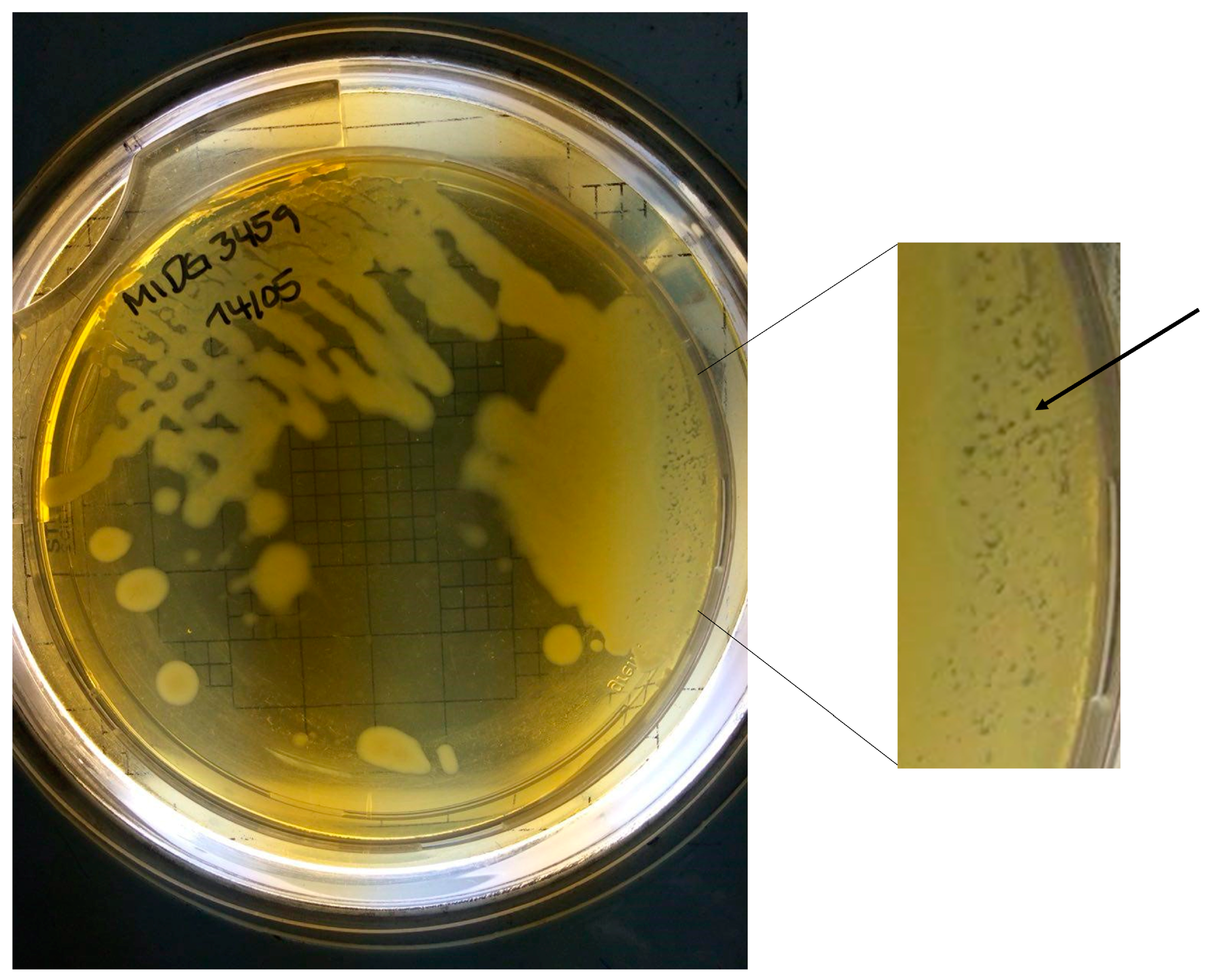
3.3. Plaque Formation in 3459 Grown on a BHI-NAD Plate
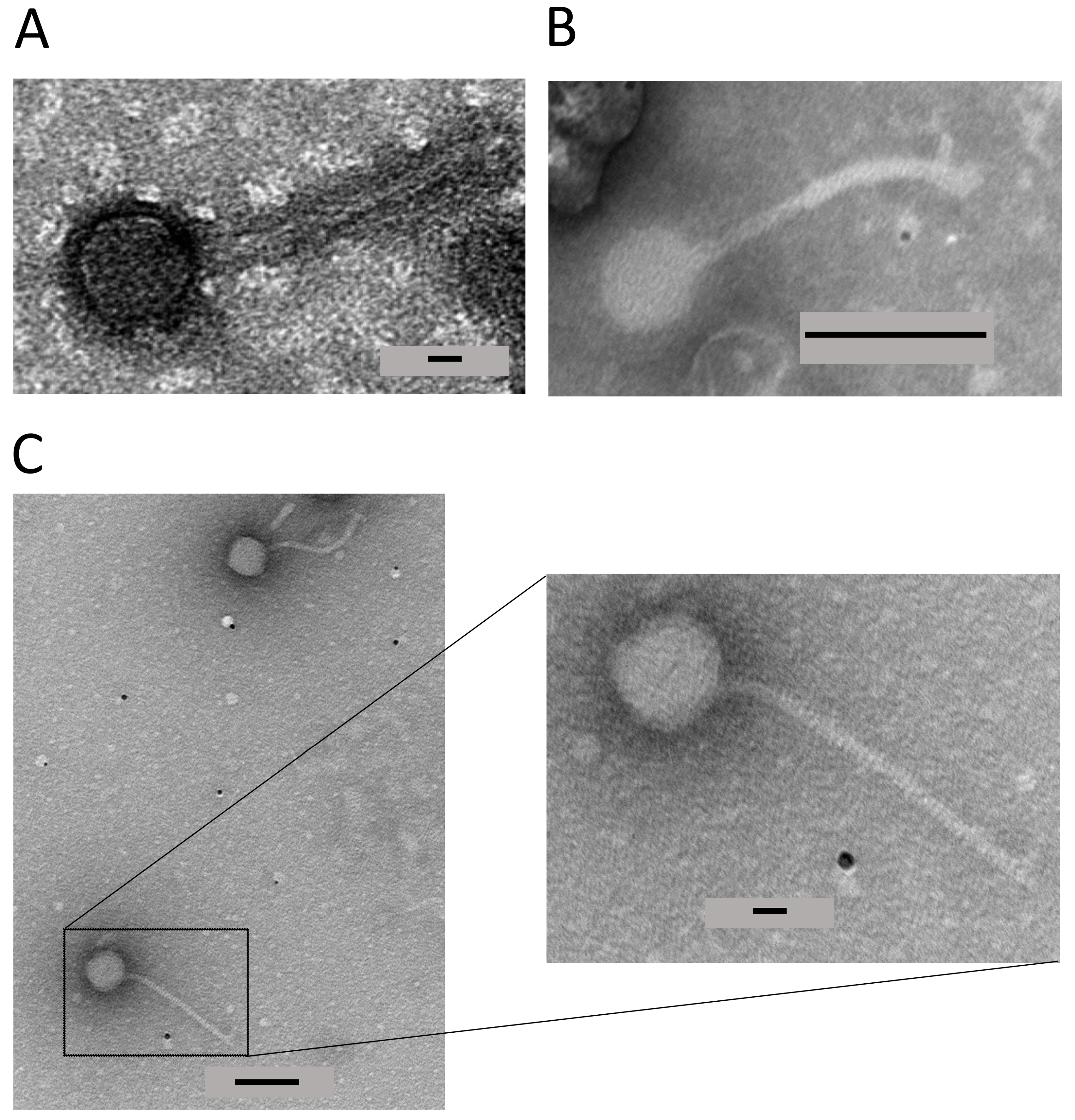
3.4. TEM Identifies a Mu-like Phage in 3459
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sassu, E.L.; Bossé, J.T.; Tobias, T.J.; Gottschalk, M.; Langford, P.R.; Hennig-Pauka, I. Update on Actinobacillus pleuropneumoniae-knowledge, gaps and challenges. Transbound. Emerg. Dis. 2018, 65 (Suppl. 1), 72–90. [Google Scholar] [CrossRef] [PubMed]
- Bossé, J.T.; Janson, H.; Sheehan, B.J.; Beddek, A.J.; Rycroft, A.N.; Kroll, J.S.; Langford, P.R. Actinobacillus pleuropneumoniae: Pathobiology and pathogenesis of infection. Microbes Infect. 2002, 4, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Chiers, K.; De Waele, T.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F. Virulence factors of Actinobacillus pleuropneumoniae involved in colonization, persistence and induction of lesions in its porcine host. Vet. Res. 2010, 41, 65. [Google Scholar] [CrossRef]
- Frey, J. Virulence in Actinobacillus pleuropneumoniae and RTX toxins. Trends Microbiol. 1995, 3, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M. The challenge of detecting herds sub-clinically infected with Actinobacillus pleuropneumoniae. Vet. J. 2015, 206, 30–38. [Google Scholar] [CrossRef]
- Hathroubi, S.; Loera-Muro, A.; Guerrero-Barrera, A.L.; Tremblay, Y.D.N.; Jacques, M. Actinobacillus pleuropneumoniae biofilms: Role in pathogenicity and potential impact for vaccination development. Anim. Health Res. Rev. 2018, 19, 17–30. [Google Scholar] [CrossRef]
- Langford, P.R.; Stringer, O.W.; Li, Y.; Bossé, J.T. Application of the MISTEACHINGS disease susceptibility framework to Actinobacillus pleuropneumoniae to identify research gaps: An exemplar of a veterinary pathogen. Anim. Health Res. Rev. 2021, 22, 120–135. [Google Scholar] [CrossRef]
- Loera-Muro, A.; Angulo, C. New trends in innovative vaccine development against Actinobacillus pleuropneumoniae. Vet. Microbiol. 2018, 217, 66–75. [Google Scholar] [CrossRef]
- Michael, G.B.; Bossé, J.T.; Schwarz, S. Antimicrobial resistance in Pasteurellaceae of veterinary origin. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Nahar, N.; Turni, C.; Tram, G.; Blackall, P.J.; Atack, J.M. Actinobacillus pleuropneumoniae: The molecular determinants of virulence and pathogenesis. Adv. Microb. Physiol. 2021, 78, 179–216. [Google Scholar] [CrossRef]
- Soto Perezchica, M.M.; Guerrero Barrera, A.L.; Avelar Gonzalez, F.J.; Quezada Tristan, T.; Macias Marin, O. Actinobacillus pleuropneumoniae, surface proteins and virulence: A review. Front. Vet. Sci. 2023, 10, 1276712. [Google Scholar] [CrossRef] [PubMed]
- Stringer, O.W.; Li, Y.; Bossé, J.T.; Langford, P.R. JMM Profile: Actinobacillus pleuropneumoniae: A major cause of lung disease in pigs but difficult to control and eradicate. J. Med. Microbiol. 2022, 71, 001483. [Google Scholar] [CrossRef] [PubMed]
- Boeters, M.; Garcia-Morante, B.; van Schaik, G.; Segales, J.; Rushton, J.; Steeneveld, W. The economic impact of endemic respiratory disease in pigs and related interventions—A systematic review. Porcine Health Manag. 2023, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Crehan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Vet. Res. 2020, 51, 80. [Google Scholar] [CrossRef]
- Stringer, O.W.; Bossé, J.T.; Lacouture, S.; Gottschalk, M.; Fodor, L.; Angen, Ø.; Velazquez, E.; Penny, P.; Lei, L.; Langford, P.R.; et al. Proposal of Actinobacillus pleuropneumoniae serovar 19, and reformulation of previous multiplex PCRs for capsule-specific typing of all known serovars. Vet. Microbiol. 2021, 255, 109021. [Google Scholar] [CrossRef]
- da Silva, G.C.; Gonçalves, O.S.; Rosa, J.N.; França, K.C.; Bossé, J.T.; Santana, M.F.; Langford, P.R.; Bazzolli, D.M.S. Mobile genetic elements drive antimicrobial resistance gene spread in Pasteurellaceae species. Front. Microbiol. 2021, 12, 773284. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Song, W.; Sun, H.X.; Zhang, C.; Cheng, L.; Peng, Y.; Deng, Z.; Wang, D.; Wang, Y.; Hu, M.; Liu, W.; et al. Prophage Hunter: An integrative hunting tool for active prophages. Nucleic Acids Res. 2019, 47, W74–W80. [Google Scholar] [CrossRef] [PubMed]
- Guitart-Matas, J.; Gonzalez-Escalona, N.; Maguire, M.; Vilaro, A.; Martinez-Urtaza, J.; Fraile, L.; Migura-Garcia, L. Revealing genomic insights of the unexplored porcine pathogen Actinobacillus pleuropneumoniae using whole genome sequencing. Microbiol. Spectr. 2022, 10, e0118522. [Google Scholar] [CrossRef] [PubMed]
- Prado, I.G.O.; da Silva, G.C.; Crispim, J.S.; Vidigal, P.M.P.; Nascimento, M.; Santana, M.F.; Bazzolli, D.M.S. Comparative genomics of Actinobacillus pleuropneumoniae serotype 8 reveals the importance of prophages in the genetic variability of the species. Int. J. Genom. 2020, 2020, 9354204. [Google Scholar] [CrossRef]
- Crespo Fernandez, R. PluMu: A Mu-like Bacteriophage Infecting the Major Pig Pathogen Actinobacillus pleuropneumoniae. MSc Thesis, Imperial College, London, UK, 2015. [Google Scholar]
- Taylor, A.L. Bacteriophage-induced mutation in Escherichia coli. Proc. Natl. Acad. Sci. USA 1963, 50, 1043–1051. [Google Scholar] [CrossRef]
- Jakhetia, R.; Verma, N.K. Identification and molecular characterisation of a novel Mu-Like bacteriophage, SfMu, of Shigella flexneri. PLoS ONE 2015, 10, e0124053. [Google Scholar] [CrossRef]
- Zehr, E.S.; Tabatabai, L.B.; Bayles, D.O. Genomic and proteomic characterization of SuMu, a Mu-like bacteriophage infecting Haemophilus parasuis. BMC Genom. 2012, 13, 331. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.H.; Cook, S.R.; Alexander, T.W.; Klima, C.L.; Niu, Y.D.; Selinger, L.B.; McAllister, T.A. Investigation of Mannheimia haemolytica bacteriophages relative to host diversity. J. Appl. Microbiol. 2013, 114, 1592–1603. [Google Scholar] [CrossRef] [PubMed]
- Bossé, J.T.; Chaudhuri, R.R.; Li, Y.; Leanse, L.G.; Fernandez Crespo, R.; Coupland, P.; Holden, M.T.; Bazzolli, D.M.; Maskell, D.J.; Tucker, A.W.; et al. Complete genome sequence of MIDG2331, a genetically tractable serovar 8 clinical isolate of Actinobacillus pleuropneumoniae. Genome Announc. 2016, 4, e01667-15. [Google Scholar] [CrossRef]
- Grace, T.D. Establishment of four strains of cells from insect tissues grown in vitro. Nature 1962, 195, 788–789. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Han, S.; Saha, S.; Oler, E.; Peters, H.; Grant, J.R.; Stothard, P.; Gautam, V. PHASTEST: Faster than PHASTER, better than PHAST. Nucleic Acids Res. 2023, 51, W443–W450. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Lipman, D.J. Protein database searches for multiple alignments. Proc. Natl. Acad. Sci. USA 1990, 87, 5509–5513. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Sun, W.; Hattman, S.; Kool, E. Interaction of the bacteriophage Mu transcriptional activator protein, C, with its target site in the mom promoter. J. Mol. Biol. 1997, 273, 765–774. [Google Scholar] [CrossRef]
- Langford, P.R.; Loynds, B.M.; Kroll, J.S. Cloning and molecular characterization of Cu,Zn superoxide dismutase from Actinobacillus pleuropneumoniae. Infect. Immun. 1996, 64, 5035–5041. [Google Scholar] [CrossRef] [PubMed]
- Bossé, J.T.; Sinha, S.; Li, M.S.; O’Dwyer, C.A.; Nash, J.H.; Rycroft, A.N.; Kroll, J.S.; Langford, P.R. Regulation of pga operon expression and biofilm formation in Actinobacillus pleuropneumoniae by sigmaE and H-NS. J. Bacteriol. 2010, 192, 2414–2423. [Google Scholar] [CrossRef] [PubMed]
- Harshey, R.M. Transposable phage Mu. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef]
- De Castro, E.; Hulo, C.; Masson, P.; Auchincloss, A.; Bridge, A.; Le Mercier, P. ViralZone 2024 provides higher-resolution images and advanced virus-specific resources. Nucleic Acids Res. 2024, 52, D817–D821. [Google Scholar] [CrossRef] [PubMed]
- Margolin, W.; Rao, G.; Howe, M.M. Bacteriophage Mu late promoters: Four late transcripts initiate near a conserved sequence. J. Bacteriol. 1989, 171, 2003–2018. [Google Scholar] [CrossRef] [PubMed]
- Zehr, E.S.; Tabatabai, L.B. Detection of a bacteriophage gene encoding a Mu-like portal protein in Haemophilus parasuis reference strains and field isolates by nested polymerase chain reaction. J. Vet. Diagn. Investig. 2011, 23, 538–542. [Google Scholar] [CrossRef]
- Herriott, R.M.; Meyer, E.Y.; Vogt, M.; Modan, M. Defined medium for growth of Haemophilus influenzae. J. Bacteriol. 1970, 101, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, A.; Bossé, J.T.; Kroll, J.S.; Langford, P.R. Analysis of differential protein expression in Actinobacillus pleuropneumoniae by Surface Enhanced Laser Desorption Ionisation--ProteinChip (SELDI) technology. Vet. Microbiol. 2004, 99, 215–225. [Google Scholar] [CrossRef]
- Ingham, A.; Zhang, Y.; Prideaux, C. Attenuation of Actinobacillus pleuropneumoniae by inactivation of aroQ. Vet. Microbiol. 2002, 84, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Niven, D.F.; Levesque, M. V-Factor-dependent growth of Actinobacillus pleuropneumoniae biotype 2 (Bertschinger 2008/76). Int. J. Syst. Bacteriol. 1988, 38, 319–320. [Google Scholar] [CrossRef]
- Wagner, T.K.; Mulks, M.H. Identification of the Actinobacillus pleuropneumoniae leucine-responsive regulatory protein and its involvement in the regulation of in vivo-induced genes. Infect. Immun. 2007, 75, 91–103. [Google Scholar] [CrossRef]
- Summer, E.J.; Gonzalez, C.F.; Carlisle, T.; Mebane, L.M.; Cass, A.M.; Savva, C.G.; LiPuma, J.; Young, R. Burkholderia cenocepacia phage BcepMu and a family of Mu-like phages encoding potential pathogenesis factors. J. Mol. Biol. 2004, 340, 49–65. [Google Scholar] [CrossRef]
- Fogg, P.C.; Hynes, A.P.; Digby, E.; Lang, A.S.; Beatty, J.T. Characterization of a newly discovered Mu-like bacteriophage, RcapMu, in Rhodobacter capsulatus strain SB1003. Virology 2011, 421, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Bossé, J.T.; Nash, J.H.; Kroll, J.S.; Langford, P.R. Harnessing natural transformation in Actinobacillus pleuropneumoniae: A simple method for allelic replacements. FEMS Microbiol. Lett. 2004, 233, 277–281. [Google Scholar] [CrossRef]
- Bossé, J.T.; Soares-Bazzolli, D.M.; Li, Y.; Wren, B.W.; Tucker, A.W.; Maskell, D.J.; Rycroft, A.N.; Langford, P.R.; BRaDP1T Consortium. The generation of successive unmarked mutations and chromosomal insertion of heterologous genes in Actinobacillus pleuropneumoniae using natural transformation. PLoS ONE 2014, 9, e111252. [Google Scholar] [CrossRef]
- Zalewska-Piatek, B. Phage therapy-challenges, opportunities and future prospects. Pharmaceuticals 2023, 16, 1638. [Google Scholar] [CrossRef]
- Stygar, A.H.; Niemi, J.K.; Oliviero, C.; Laurila, T.; Heinonen, M. Economic value of mitigating Actinobacillus pleuropneumoniae infections in pig fattening herds. Agric. Syst. 2016, 144, 113–121. [Google Scholar] [CrossRef]
- Shan, X.; Szabo, R.E.; Cordero, O.X. Mutation-induced infections of phage-plasmids. Nat. Commun. 2023, 14, 2049. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers | 5′–3′ | Amplicon Size (bp) |
|---|---|---|---|
| Mu c | c_For | CGCAGTTGTTAGTTGAAGTG | 264 |
| c_Rev | ATTTGATGTCGCTTCCGTTG | ||
| Mu tail | Tail_For | TTGGCACGATTATTGAAGCAAAACG | 448 |
| Tail_Rev | AAACACTCGTGACCCGTCATAAGG | ||
| sodC | SodC_For | GATTAAGCTACCTACAAGCGGTGCAC | 1035 |
| SodC_Rev | CCGTGTCGACATCCTTGTTTCCTTTTACAG |
| PluMu 3457-1 | PluMu 3457-2 | PluMu 3459-1 | PluMu 3459-2 | |
|---|---|---|---|---|
| Region Length (bp) | 41,862 | 23,041 | 36,028 | 49,839 |
| Completeness (score) | Intact (140) | Intact (100) | Intact (150) | Intact (140) |
| tRNA | 0 | 0 | 0 | 0 |
| Total proteins | 56 | 35 | 48 | 68 |
| Phage hit proteins | 41 | 29 | 39 | 42 |
| Hypothetical proteins | 12 | 14 | 17 | 13 |
| Bacterial proteins | 15 | 6 | 11 | 26 |
| Attachment site prediction | No | No | No | No |
| GC% | 39.6 | 40.3 | 41.1 | 39.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartsch, L.J.; Fernandez Crespo, R.; Wang, Y.; Skinner, M.A.; Rycroft, A.N.; Cooley, W.; Everest, D.J.; Li, Y.; Bossé, J.T.; Langford, P.R. PluMu—A Mu-like Bacteriophage Infecting Actinobacillus pleuropneumoniae. Appl. Microbiol. 2024, 4, 520-535. https://doi.org/10.3390/applmicrobiol4010037
Bartsch LJ, Fernandez Crespo R, Wang Y, Skinner MA, Rycroft AN, Cooley W, Everest DJ, Li Y, Bossé JT, Langford PR. PluMu—A Mu-like Bacteriophage Infecting Actinobacillus pleuropneumoniae. Applied Microbiology. 2024; 4(1):520-535. https://doi.org/10.3390/applmicrobiol4010037
Chicago/Turabian StyleBartsch, Lee Julia, Roberto Fernandez Crespo, Yunfei Wang, Michael A. Skinner, Andrew N. Rycroft, William Cooley, David J. Everest, Yanwen Li, Janine T. Bossé, and Paul R. Langford. 2024. "PluMu—A Mu-like Bacteriophage Infecting Actinobacillus pleuropneumoniae" Applied Microbiology 4, no. 1: 520-535. https://doi.org/10.3390/applmicrobiol4010037