
Biogenic Amine Formation in Artisan Galotyri PDO Acid-Curd Cheeses Fermented with Greek Indigenous Starter and Adjunct Lactic Acid Bacteria Strain Combinations: Effects of Cold (4 °C) Ripening and Biotic Factors Compromising Cheese Safety


Abstract
:1. Introduction
2. Materials and Methods
2.1. LAB Strains, Selection Criteria, and Culture Conditions
2.2. Evaluation of the BA Formation Potential of the Selected Starter and Adjunct LAB Strain Combinations in Milk Model Fermentations
2.3. Measurement of pH and Determination of Sugars and Organic Acids in SRM Samples
2.4. Artisan Galotyri PDO Cheese Manufacture and Sampling
2.5. Measurement of pH and Gross Composition of the Fresh and Ripened Cheese Samples
2.6. Microbiological Analyses of the Fresh and Ripened Cheese Samples
2.7. Determination of Biogenic Amines in SRM and Galotyri Cheese Samples
2.8. Statistical Analysis
3. Results
3.1. Minimal BA Formation by All LAB Strain Combinations in SRM Cocultures
3.2. Variations in pH and the Gross Composition of the Galotyri PDO Cheese Samples Attributed to Certain Unstable Processing Factors of the Artisan Manufacturing Method
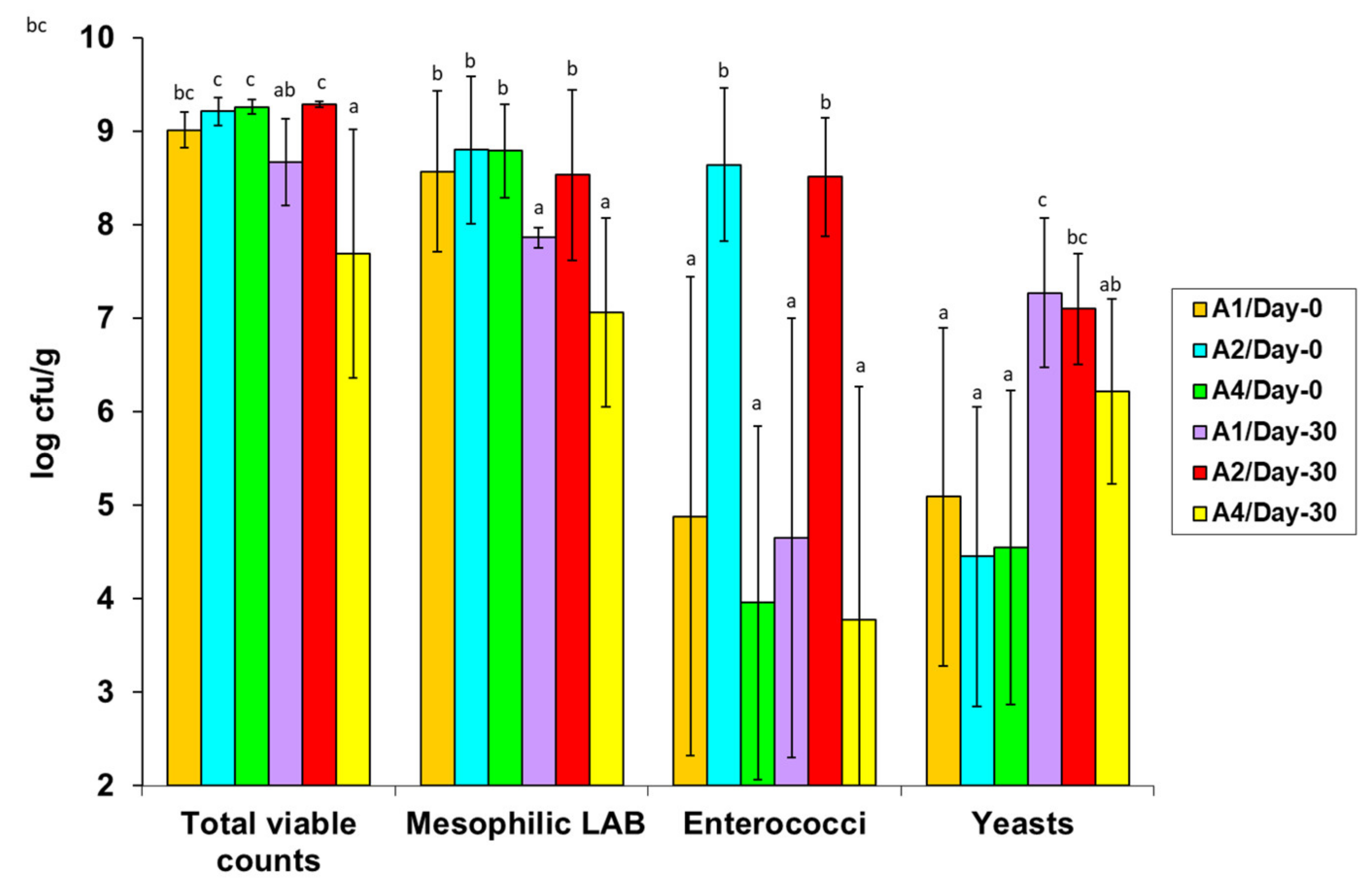
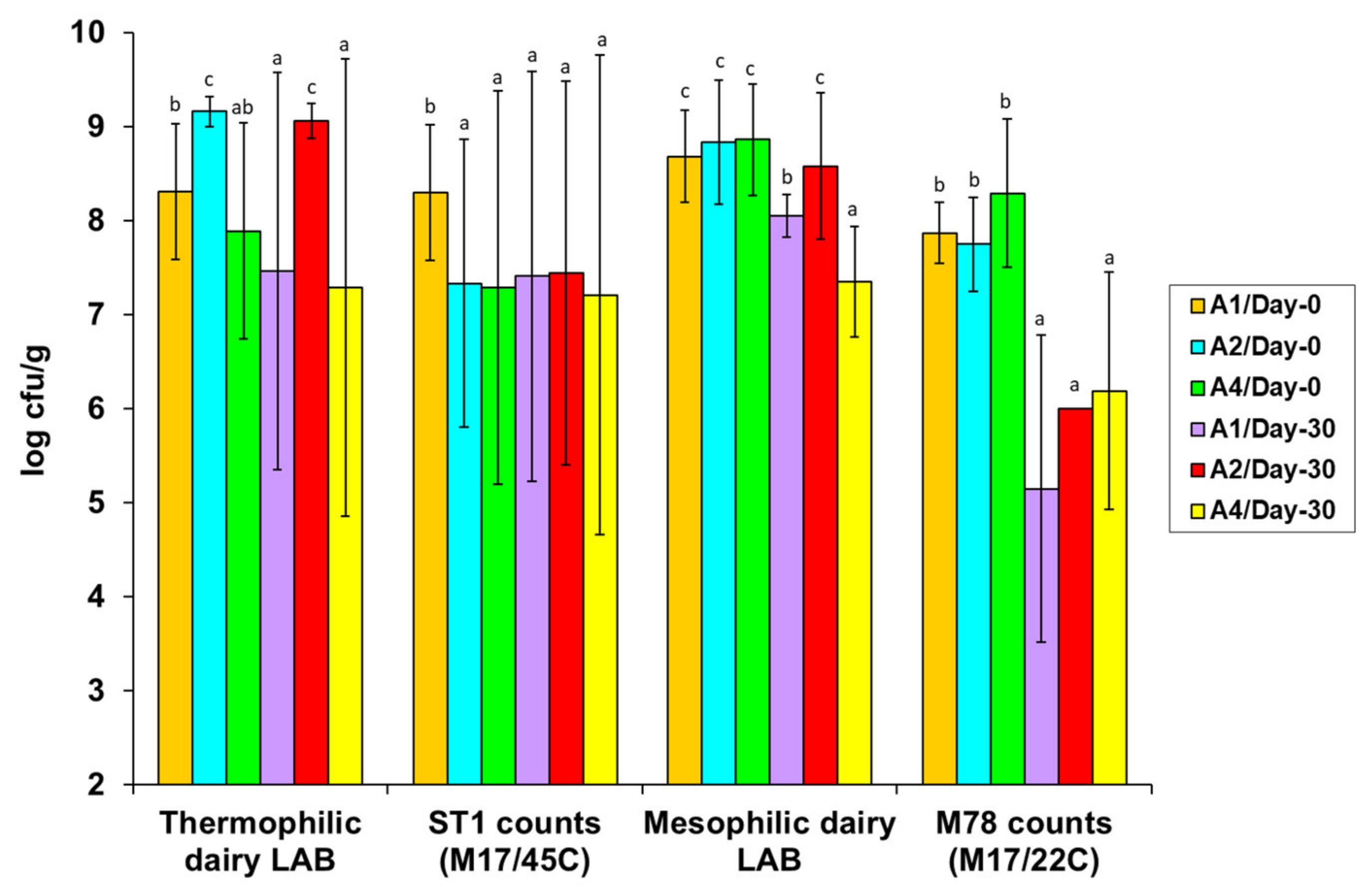
3.3. Microbial Quantification in the Artisan Galotyri PDO Cheese Samples
3.3.1. Effects of the Artisan Processing Technology and Hygiene on the Microbiological Quality of the Fresh Cheeses—Variations in the Populations of the Natural Starter and Adjunct Strains
3.3.2. Effects of Cold Ripening on the Microbiological Characteristics of the Ripened Cheeses—Variations in the Surviving Populations of the Natural Starter and Adjunct Strains
3.4. Major Variations in the Biogenic Amine (BA) Amounts of the Artisan Galotyri Cheese Samples Associated with the Microbial Quality of the Fresh Cheeses and Cold Ripening Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Linares, D.M.; Martín, M.C.; Ladero, V.; Alvarez, M.A.; Fernández, M. Biogenic amines in dairy products. Crit. Rev. Food Sci. Nutr. 2011, 51, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef]
- Gardini, F.; Özogul, Y.; Suzzi, G.; Tabanelli, G.; Özogul, F. Technological factors affecting biogenic amine content in foods: A review. Front. Microbiol. 2016, 7, 1218. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; del Río, B.; Ladero, V.; Martínez, N.; Fernández, M.; Martin, M.C.; Álvarez, M.A. Factors influencing biogenic amines accumulation in dairy products. Front. Microbiol. 2012, 3, 180. [Google Scholar] [CrossRef] [PubMed]
- Moniente, M.; Botello-Morte, L.; García-Gonzalo, D.; Virto, R.; Pagán, R.; Ferreira, V.; Ontañón, I. Combination of SPE and fluorescent detection of AQC-derivatives for the determination at sub- mg/L levels of biogenic amines in dairy products. Food Res. Int. 2023, 165, 112448. [Google Scholar] [CrossRef] [PubMed]
- Moniente, M.; Botello-Morte, L.; García-Gonzalo, D.; Pagán, R.; Ontañón, I. Analytical strategies for the determination of biogenic amines in dairy products. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3612–3646. [Google Scholar] [CrossRef] [PubMed]
- Benkerroum, N. Biogenic amines in dairy products: Origin, incidence, and control means. Compr. Rev. Food Sci. Food Saf. 2016, 15, 801–826. [Google Scholar] [CrossRef]
- Moniente, M.; García-Gonzalo, D.; Ontañón, I.; Pagán, R.; Botello-Morte, L. Histamine accumulation in dairy products: Microbial causes, techniques for the detection of histamine-producing microbiota, and potential solutions. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1481–1523. [Google Scholar] [CrossRef]
- Bunkova, L.; Adamcova, G.; Hudcova, K.; Velichova, H.; Pachlova, V.; Lorencova, E.; Bunka, F. Monitoring of biogenic amines in cheeses manufactured at small-scale farms and in fermented dairy products in the Czech Republic. Food Chem. 2013, 141, 548–551. [Google Scholar] [CrossRef]
- Burdychova, R.; Komprda, T. Biogenic amine-forming microbial communities in cheese. FEMS Microbiol. Lett. 2007, 276, 149–155. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, L338, 1–26. [Google Scholar]
- European Food Safety Authority (EFSA). Scientific opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393–2486. [Google Scholar]
- Ten Brink, B.; Damink, C.; Joosten, H.; Huis in’t Veld, J.H.J. Occurrence and formation of biologically active amines in foods. Int. J. Food Microbiol. 1990, 11, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Pircher, A.; Bauer, F.; Paulsen, P. Formation of cadaverine, histamine, putrescine and tyramine by bacteria isolated from meat, fermented sausages and cheeses. Eur. Food Res. Technol. 2007, 226, 225–231. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, V.; Moret, S.; Rondinini, G. The capacity of Enterobacteriaceae species to produce biogenic amines in cheese. Lett. Appl. Microbiol. 2000, 31, 169–173. [Google Scholar] [CrossRef]
- Coton, M.; Delbés-Paus, C.; Irlinger, E.; Desmasures, N.; Le Fleche, A.; Stahl, V.; Montel, M.C.; Coton, E. Diversity and assessment of potential risk factors of Gram-negative isolates associated with French cheeses. Food Microbiol. 2011, 29, 88–98. [Google Scholar] [CrossRef]
- Espinosa-Pesqueira, D.; Roig-Sagués, A.X.; Hernández-Herrero, M.M. Screening method to evaluate amino acid-decarboxylase activity of bacteria present in Spanish artisanal ripened cheeses. Foods 2018, 7, 182. [Google Scholar] [CrossRef]
- Ryser, L.T.; Arias-Roth, E.; Berthoud, H.; Delbès-Paus, C.; Chassard, C.; Bruggmann, R.; Irmler, S. Cadaverine, putrescine, and histamine formation by Morganella morganii in raclette-type cheese. Int. Dairy J. 2022, 129, 105362. [Google Scholar] [CrossRef]
- Wyder, M.T.; Bachmann, H.P.; Puhan, Z. Role of selected yeasts in cheese ripening: An evaluation in foil wrapped Raclette cheese. Lebensm. Wiss. Technol. 1999, 32, 333–343. [Google Scholar] [CrossRef]
- Roig-Sagués, A.X.; Molina, A.P.; Hernández-Herrero, M.M. Histamine and tyramine-forming microorganisms in Spanish traditional cheeses. Eur. Food Res. Technol. 2002, 215, 96–100. [Google Scholar] [CrossRef]
- Gardini, F.; Tofalo, R.; Belletti, N.; Iucci, I.; Suzzi, G.; Torriani, S.; Guerzoni, M.E.; Lanciotti, R. Characterization of yeasts involved in the ripening of Pecorino Crotonese cheese. Food Microbiol. 2006, 23, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Montel, M.C.; Buchin, S.; Mallet, A.; Delbés-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Gardini, F.; Martuscelli, M.; Caruso, M.C.; Galgano, F.; Crudele, M.A.; Favati, F.; Guerzoni, M.E.; Suzzi, G. Effects of pH, temperature, and NaCl concentration on the growth kinetics, proteolytic activity and biogenic amine production of Enterococcus faecalis. Int. J. Food Microbiol. 2001, 64, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Bargossi, E.; Gardini, F.; Gatto, V.; Montanari, C.; Torriani, S.; Tabanelli, G. The capability of tyramine production and correlation between phenotypic and genetic characteristics of Enterococcus faecium and Enterococcus faecalis strains. Front. Microbiol. 2015, 6, 1371. [Google Scholar] [CrossRef] [PubMed]
- Houicher, A.; Kuley, E.; Bensid, A.; Yazgan, H.; Özogul, F. In vitro study of biogenic amine production and gastrointestinal stress tolerance by some enterococci strains. J. Sci. Food Agric. 2023, 104, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Fernández, M.; Calles-Enríquez, M.; Sánchez-Llana, E.; Cañedo, E.; Martín, M.C.; Alvarez, M.A. Is the production of biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Joosten, H.; Northolt, M. Detection, growth and amine-producing capacity of lactobacilli in cheese. Appl. Environ. Microbiol. 1989, 55, 2356–2359. [Google Scholar] [CrossRef] [PubMed]
- Ascone, P.; Mauer, J.; Haldemann, J.; Irmler, S.; Berthoud, H.; Portmann, R.; Fröhlich-Wyder, M.T.; Wechsler, D. Prevalence and diversity of histamine-forming Lactobacillus parabuchneri strains in raw milk and cheese—A case study. Int. Dairy J. 2017, 70, 26–33. [Google Scholar] [CrossRef]
- Wechsler, D.; Irmel, S.; Berthoud, H.; Portmann, R.; Badertscher, R.; Bisig, W.; Schafroth, K.; Frohlich-Wyder, M.T. Influence of inoculums level of Lactobacillus parabuchneri in vat milk and of the cheese-making conditions on histamine formation during ripening. Int. Dairy J. 2021, 113, 104883. [Google Scholar] [CrossRef]
- Botello-Morte, L.; Moniente, M.; Gil-Ramírez, Y.; Virto, R.; García-Gonzalo, D.; Pagán, R. Identification by means of molecular tools of the microbiota responsible for the formation of histamine accumulated in commercial cheeses in Spain. Food Control 2022, 133A, 108595. [Google Scholar] [CrossRef]
- Moniente, M.; García-Gonzalo, D.; Llamas-Arriba, M.G.; Virto, R.; Ontañón, I.; Pagán, R.; Botello-Morte, L. Potential of histamine-degrading microorganisms and diamine oxidase (DAO) for the reduction of histamine accumulation along the cheese ripening process. Food Res. Int. 2022, 160, 111735. [Google Scholar] [CrossRef] [PubMed]
- Sarquis, A.; Bajrami, D.; Mizaikoff, B.; Ladero, V.; Alvarez, M.A.; Fernández, M. Characterization of the biofilms formed by histamine-producing Lentilactobacillus parabuchneri strains in the dairy environment. Foods 2023, 12, 1503. [Google Scholar] [CrossRef] [PubMed]
- Coton, M.; Lebreton, M.; Salas, M.L.; Garnier, L.; Navarri, M.; Pawtowski, A.; Le Blay, G.; Valence, F.; Coton, E.; Mounier, J. Biogenic amine and antibiotic resistance profiles determined for lactic acid bacteria and a propionibacterium prior to use as antifungal bioprotective cultures. Int. Dairy J. 2018, 85, 21–26. [Google Scholar] [CrossRef]
- Calles-Enríquez, M.; Eriksen, B.H.; Andersen, P.S.; Rattray, F.P.; Johansen, A.H.; Fernández, M.; Ladero, V.; Álvarez, M.A. Sequencing and transcriptional analysis of the Streptococcus thermophilus histamine biosynthesis gene cluster: Factors that affect differential hdcA expression. Appl. Environ. Microbiol. 2010, 76, 6231–6238. [Google Scholar] [CrossRef] [PubMed]
- Buňková, L.; Buňka, F.; Pollaková, E.; Podešová, T.; Dráb, V. The effect of lactose, NaCl and an aero/anaerobic environment on the tyrosine decarboxylase activity of Lactococcus lactis subsp. cremoris and Lactococcus lactis subsp. lactis. Int. J. Food Microbiol. 2011, 147, 112–119. [Google Scholar] [PubMed]
- Ladero, V.; Rattray, F.P.; Mayo, B.; Martin, M.C.; Fernández, M.; Álvarez, M.A. Sequencing and transcriptional analysis of the biosynthesis gene cluster of putrescine-producing Lactococcus lactis. Appl. Environ. Microbiol. 2011, 77, 6409–6418. [Google Scholar] [CrossRef]
- Leuschner, R.G.; Heidel, M.; Hammes, W.P. Histamine and tyramine degradation by food fermenting microorganisms. Int. J. Food Microbiol. 1998, 39, 1–10. [Google Scholar] [CrossRef]
- Herrero-Fresno, A.; Martínez, N.; Sáncez-Llana, E.; Díaz, M.; Fernández, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. Lactobacillus casei strains isolated from cheese reduce biogenic amine accumulation in an experimental model. Int. J. Food Microbiol. 2012, 157, 297–304. [Google Scholar] [CrossRef]
- Bäumlisberger, M.; Moellecken, U.; König, H.; Claus, H. The potential of the yeast Debaryomyceshansenii H525 to degrade biogenic amines in food. Microorganisms 2015, 3, 839–850. [Google Scholar] [CrossRef]
- Guarcello, R.; De Angelis, M.; Settanni, L.; Formiglio, S.; Gaglio, R.; Minervini, F.; Moschetti, G.; Gobetti, M. Selection of amine-oxidizing dairy lactic acid bacteria and identification of the enzyme and gene involved in the absence of biogenic amines. Appl. Environ. Microbiol. 2016, 82, 6870–6880. [Google Scholar] [CrossRef] [PubMed]
- Tittarelli, F.; Perpetuini, G.; Di Gianvito, P.; Tofalo, R. Biogenic amines producing and degrading bacteria: A snapshot from raw ewes’ cheese. LWT 2019, 101, 1–9. [Google Scholar] [CrossRef]
- Samelis, J.; Lianou, A.; Kakouri, A.; Delbès, C.; Rogelj, I.; Matijašic, B.B.; Montel, M.C. Changes in the microbial composition of raw milk induced by thermization treatments applied prior to traditional Greek hard cheese processing. J. Food Prot. 2009, 72, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N. Microbiological characteristics of Greek traditional cheeses. Small Rumin. Res. 2011, 101, 17–32. [Google Scholar] [CrossRef]
- Bintsis, T.; Athanasoulsas, A. Dairy Starter Cultures. In Dairy Microbiology: A Practical Approach; Papademas, P., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 114–154. [Google Scholar]
- Samelis, J.; Kakouri, A. Major technological differences between an industrial-type and five artisan-type Greek PDO Galotyri market cheeses as revealed by great variations in their lactic acid microbiota. AIMS Agric. Food 2019, 4, 685–710. [Google Scholar] [CrossRef]
- Rhoades, J.; Anastasiou, I.; Michailidou, S.; Koinidis, A.; Doulgerakis, C.; Alexa, E.A.; Alvarez-Ordonez, A.; Argiriou, A.; Likotrafiti, E. Microbiological analysis of Greek Protected Designation of Origin cheeses and characterization of the isolated lactic acid bacteria. Int. Dairy J. 2021, 123, 105183. [Google Scholar] [CrossRef]
- Samelis, J.; Tsanasidou, C.; Bosnea, L.; Ntziadima, C.; Gatzias, I.; Kakouri, A.; Pappas, D. Pilot-scale production of traditional Galotyri PDO cheese from boiled ewes’ milk fermented with the aid of Greek indigenous Lactococcus lactis subsp. cremoris starter and Lactiplantibacillus plantarum adjunct strains. Fermentation 2023, 9, 345. [Google Scholar]
- Pappa, E.C.; Kondyli, E. Descriptive characteristics and cheesemaking technology of Greek cheeses not listed in the EU geographical indications registers. Dairy 2023, 4, 43–67. [Google Scholar] [CrossRef]
- Samelis, J.; Argyri, A.; Doulgeraki, A.; Koukkou, A.I.; Skandamis, P.; Athanasoulas, A.; Pappas, D.; Chorianopoulos, N. Development of novel biofunctional foods and total quality enhancement of traditional dairy products by suitable management of their microbial ecology—BIO TRUST. In Proceedings of the 26th International ICFMH Conference FoodMicro, Freie Universität, Berlin, Germany, 3–6 September 2018; P1.62. p. 152. [Google Scholar]
- Anonymous. Recognition of a Protected Denomination of Origin (P.D.O.) for Galotyri cheese. In Newspaper of the Government of the Republic of Greece; National Publishing Office: Athens, Greece, 1994; pp. 51–62. [Google Scholar]
- Samelis, J.; Kakouri, A. Microbiological characterization of Greek Galotyri cheese PDO products relative to whether they are marketed fresh or ripened. Fermentation 2022, 8, 492. [Google Scholar] [CrossRef]
- Sameli, N.; Skandamis, P.N.; Samelis, J. Application of Enterococcus faecium KE82, an enterocin A-B-P-producing strain, as an adjunct culture enhances inactivation of Listeria monocytogenes during traditional protected designation of origin Galotyri processing. J. Food Prot. 2021, 84, 87–98. [Google Scholar] [CrossRef]
- Tsanasidou, C.; Asimakoula, S.; Sameli, N.; Fanitsios, C.; Vandera, E.; Bosnea, L.; Koukou, A.I.; Samelis, J. Safety evaluation, biogenic amine formation, and enzymatic activity profiles of autochthonous enterocin-producing Greek cheese isolates of the Enterococcus faecium/durans group. Microorganisms 2021, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Asimakoula, S.; Giaka, K.; Fanitsios, C.; Kakouri, A.; Vandera, E.; Samelis, J.; Koukkou, A.-I. Monitoring growth compatibility and bacteriocin gene transcription of adjunct and starter lactic acid bacterial strains in milk. J. Food Prot. 2021, 84, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Kakouri, A. Cell growth density and nisin A activity of the indigenous Lactococcus lactis subsp. cremoris M78 costarter depend strongly on inoculation levels of a commercial Streptococcus thermophilus starter in milk: Practical aspects for traditional Greek cheese processors. J. Food Prot. 2020, 83, 542–551. [Google Scholar] [PubMed]
- Parapouli, M.; Delbés-Paus, C.; Kakouri, A.; Koukkou, A.-I.; Montel, M.C.; Samelis, J. Characterization of a wild, novel nisin A-producing Lactococcus strain with an L. lactis subsp. cremoris genotype and an L. lactis subsp. lactis phenotype isolated from Greek raw milk. Appl. Environ. Microbiol. 2013, 79, 3476–3484. [Google Scholar]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef] [PubMed]
- ISO 5534|IDF 4; Cheese and Processed Cheese, Determination of the Total Solids Content (Reference Method). International organization for standardization (ISO): Geneva, Switzerland, 2004.
- BS 696-2:1989; Gerber Method for the Determination of Fat in Milk and Milk Products. British Standard 696; British Standards Institution: London, UK, 1955.
- ISO 8968-1/IDF 20-1; Milk and Milk Products—Determination of Nitrogen Content—Part 1: Kjeldahl Principle and Crude Protein Calculation. International organization for standardization (ISO): Geneva, Switzerland, 2014.
- Kosikowski, F.V. Cheese and Fermented Milk Foods; Edwards Brothers: Ann Arbor, MI, USA, 1982. [Google Scholar]
- AOAC. Ash of cheese 935.42 method. In Official Methods of Analysis EI, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995; p. 59. [Google Scholar]
- Ling, R. A Text Book of Dairy Chemistry, 3rd ed.; Chapman & Hall: London, UK, 2001. [Google Scholar]
- Sameli, N.; Sioziou, E.; Bosnea, L.; Kakouri, A.; Samelis, J. Assessment of the spoilage microbiota during refrigerated (4 °C) vacuum-packed storage of fresh Greek Anthotyros whey cheese without or with a crude enterocin A-B-P-containing extract. Foods 2021, 10, 2946. [Google Scholar] [CrossRef] [PubMed]
- Eerola, S.; Hinkkanen, R.; Lindfors, E.; Hirvi, T. Liquid chromatographic determination of biogenic amines in dry sausages. J. AOAC Int. 1993, 76, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Granvalet, C.; Cotin, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef]
- Fiechter, G.; Sivec, G.; Mayer, H.K. Application of UHPLC for the simultaneous analysis of free aminoacids and biogenic amines in ripened Acid-Curd cheeses. J. Chrom. B-Anal. Technol. Biomed. Life Sci. 2013, 927, 191–200. [Google Scholar] [CrossRef]
- Mayer, H.K.; Fiechter, G. UHPLC analysis of biogenic amines in different cheese varieties. Food Control 2018, 93, 9–16. [Google Scholar] [CrossRef]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Izquerdo-Pulido, M.; Vidal-Carou, M.C. Distribution of biogenic amines and pyamines in cheese. J. Food Sci. 2003, 68, 750–755. [Google Scholar] [CrossRef]
- Moniente, M.; García-Gonzalo, D.; Llamas-Arriba, M.G.; Garate, J.; Ontañón, I.; Jaureguibeitia, A.; Virto, R.; Pagán, R.; Botello-Morte, L. The significance of cheese sampling in the determination of histamine concentration: Distribution pattern of histamine in ripened cheeses. LWT 2022, 171, 114099. [Google Scholar] [CrossRef]
- Del Rio, B.; Ladero, V.; Redruello, B.; Linares, D.M.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Lactose-mediated carbon catabolite repression of putrescine production in dairy Lactococcus lactis is strain-dependent. Food Microbiol. 2015, 48, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, M.R.; Fernández-Otero, C.; Rodríguez-Alonso, P.; Fernández-No, C.; Carabal, J.I.; Centeno, J.A. Characterization of yeasts isolated from artisanal short-ripened cows’ cheeses produced in Galicia (NW Spain). Food Microbiol. 2016, 53, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Geronikou, A.; Srimahaeak, T.; Rantsiou, K.; Triantafillidis, G.; Larsen, N.; Jespersen, L. Occurrence of yeasts in white-brined cheeses: Methodologies for identification, spoilage potential and good manufacturing practices. Front. Microbiol. 2020, 11, 582778. [Google Scholar] [CrossRef] [PubMed]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Roig-Sagués, A.X.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Evaluation of biogenic amines and microbial counts throughout the ripening of goat cheeses from pasteurized or raw milk. J. Dairy Res. 2004, 71, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Roig-Sagués, A.X.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Influence of starter and nonstarter on the formation of biogenic amine in goat cheese during ripening. J. Dairy Sci. 2002, 85, 2471–2478. [Google Scholar] [CrossRef]
- Valsamaki, K.; Michaelidou, A.; Polychroniadou, A. Biogenic amine production in Feta cheese. Food Chem. 2000, 71, 259–266. [Google Scholar] [CrossRef]
- Öner, Z.; Karahan, A.G.; Aloglu, H. Changes in the microbiological and chemical characteristics of an artisanal Turkish white cheese during ripening. LWT-Food Sci. Technol. 2006, 39, 449–454. [Google Scholar] [CrossRef]
- Bonczar, G.; Filipczak-Fiutak, M.; Pluta-Kubica, A.; Duda, I.; Walczycka, M.; Staruch, L. The range of protein hydrolysis and biogenic amines content in selected acid- and rennet-curd cheeses. Chem. Pap. 2018, 72, 2599–2606. [Google Scholar] [CrossRef]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Biogenic amines and polyamines in milks and cheeses by ion-pair high performance liquid chromatography. J. Agric. Food Chem. 2000, 48, 5117–5123. [Google Scholar] [CrossRef] [PubMed]
- Ercan, S.Ş.; Soysal, Ç.; Bozkurt, H. Biogenic amine contents of fresh and mature Kashar cheeses during refrigerated storage. Food Health 2019, 5, 19–29. [Google Scholar] [CrossRef]
- Del Rio, B.; Linares, D.M.; Ladero, V.; Redruello, B.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Putrescine biosynthesis in Lactococcus lactis is trascriptionally activated at acidic pH and counteracts acidification of the cytosol. Int. J. Food. Microbiol. 2016, 236, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.K.; Raslan, A.A.; Elbadry, S.; El-Ghareeb, W.R.; Mulla, Z.S.; Bin-Jumah, M.; Abdel-Daim, M.M.; Darwish, W.S. Levels of biogenic amines in cheese: Correlation to microbial status, dietary intakes, and their health assessment. Environ. Sci. Pollute Res. 2020, 27, 44452–44459. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, B.; Redruello, B.; Ladero, V.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Putrescine production by Lactococcus lactis subsp. cremoris CECT 8666 is reduced by NaCl via a decrease in bacterial growth and the repression of genes involved in putrescine production. Int. J. Food Microbiol. 2016, 232, 1–6. [Google Scholar]
- Del Rio, B.; Fernández, M.; Redruello, B.; Ladero, V.; Alvarez, M.A. New insights into the toxicological effects of dietary biogenic amines. Food Chem. 2024, 435, 137558. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, P.; Grossgut, R.; Bauer, F.; Rauscher-Gabernig, E. Estimates of maximum tolerable levels of tyramine content in foods in Austria. J Food Nutr. Res. 2012, 51, 52–59. [Google Scholar]
- Fernández, M.; Linares, D.M.; del Rio, B.; Ladero, V.; Alvarez, M.A. HPLC quantification of biogenic amines in cheeses: Correlation with PCR-detection of tyramine-producing microorganisms. J. Dairy Res. 2007, 74, 276–282. [Google Scholar] [CrossRef]
- Til, H.P.; Falke, H.E.; Prinsen, M.K.; Willems, M.I. Acute and subacute toxicity of tyramine, spermidine, spermine, putrescine and cadaverine in rats. Food Chem. Toxicol. 1997, 35, 337–348. [Google Scholar] [CrossRef]
- Del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernández, M.; Martin, M.C.; Alvarez, M.A. The biogenic amines putrescine and cadaverine show in vitro cytotoxicity at concentrations that can be found in foods. Sci. Rep. 2019, 9, 120. [Google Scholar] [CrossRef]
- Rauscher-Gabernig, E.; Gabernig, R.; Brueller, W.; Grossgut, R.; Bauer, F.; Paulsen, P. Dietary exposure assessment of putrescine and cadaverine and derivation of tolerable levels in selected foods consumed in Austria. Eur. Food Res. Technol. 2012, 235, 209–220. [Google Scholar] [CrossRef]
- Linares, D.M.; del Rio, B.; Redruello, B.; Ladero, V.; Martin, M.C.; Fernández, M.; Ruas-Madiedo, P.; Alvarez, M.A. Comparative analysis of the in vitro cytotoxicity of the dietary amines tyramine and histamine. Food Chem. 2016, 197, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Fernández, M.; Cuesta, I.; Alvarez, M.A. Quantitative detection and identification of tyramine-producing enterococci and lactobacilli in cheese by multiplex qPCR. Food Microbiol. 2010, 27, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Spermine and spermidine are cytotoxic towards intestinal cell cultures, but are they a health hazard at concentrations found in foods? Food Chem. 2018, 269, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Vandera, E.; Parapouli, M.; Koukkou, A.-I.; Hatziloukas, E.; Samelis, J. Selection of potential probiotic lactic acid bacteria isolated from traditional dairy products by in vitro tests. In Book of Abstracts of the IAFP’s, Proceedings of the European Symposium on Food Safety, Athens, Greece, 11–13 May 2016; International Association for Food Protection: Des Moines, IA, USA, 2016; P2–24; pp. 115–116. [Google Scholar]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns—An update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Graham, K.; Stack, H.; Rea, R. Safety, beneficial and technological properties of enterococci for use in functional food applications—A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 3836–3861. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.D.; Franco, B.D.G.D. Lactobacillus plantarum: Characterization of the species and application in food production. Food Rev. Int. 2010, 26, 205–229. [Google Scholar] [CrossRef]
- Cogan, T.M.; Beresford, T.P.; Steele, J.; Broadbent, J.; Shah, N.P.; Ustunol, Z. Invited review: Advances in starter cultures and cultured foods. J. Dairy Sci. 2007, 90, 4005–4021. [Google Scholar] [CrossRef]
- Papadopoulou, O.S.; Argyri, A.A.; Varzakis, E.E.; Tassou, C.C.; Chorianopoulos, N.G. Greek functional Feta cheese: Enhancing quality and safety using a Lactobacillus plantarum strain with probiotic potential. Food Microbiol. 2018, 74, 21–33. [Google Scholar] [CrossRef]
- Yilmaz, C.; Gökmen, V. Formation of tyramine in yogurt during fermentation—Interaction between yogurt starter bacteria and Lactobacillus plantarum. Food Res. Int. 2017, 97, 288–295. [Google Scholar] [CrossRef]
- Zotou, A.; Notou, M. Study of the naphthalene-2-3-dicarboxaldehyde pre-column derivatization of biogenic mono- and diamines in mixture and fluorescence-HPLC determination. Anal. Bioanal. Chem. 2012, 403, 1039–1048. [Google Scholar] [CrossRef]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N. Chapter 9: The microfloras of traditional Greek cheeses. In Cheese and Microbes; Donnelly, C.W., Ed.; ASM Press: Washington, DC, USA, 2014. [Google Scholar]



{kind=link}
{kind=link}
| Biogenic Amine | Strain Cultured in BA Broth with 1% Histidine or 1% Tyramine | Strain Culture in BA Broth without Added Amino Acids (Control) | ||||||
|---|---|---|---|---|---|---|---|---|
| Lc. lactis ssp. cremoris M78 | E. faecium KE82 | E. faecium GL31 | L. plantarum H25 | Lc. lactis ssp. cremoris M78 | E. faecium KE82 | E. faecium GL31 | L. plantarum H25 | |
| Cadaverine | 2.05 ± 0.16 | <0.50 | 2.05 ± 0.02 | 2.13 ± 0.18 | 4.10 ± 0.22 | 3.94 ± 0.18 | 7.97 ± 0.13 | 8.16 ± 2.79 |
| Histamine | 16.26 ± 0.88 | <0.50 | 16.00 ± 0.58 | 17.00 ± 1.29 | 13.63 ± 0.00 | 13.29 ± 0.22 | 18.41 ± 0.75 | 18.10 ± 0.40 |
| Putrescine | <0.50 | <0.50 | <0.5 | <0.50 | <0.50 | <0.50 | <0.50 | <0.50 |
| Spermidine | 5.13 ± 0.84 | 1.19 ± 0.00 | 5.88 ± 0.05 | 5.60 ± 0.13 | 10.88 ± 2.65 | 10.16 ± 1.55 | 11.32 ± 0.53 | 21.94 ± 15.83 |
| Tryptamine | 2.83 ± 0.28 | <0.50 | 2.42 ± 0.86 | 2.91 ± 0.22 | 32.26 ± 6.19 | 4.10 ± 0.13 | 10.88 ± 3.27 | 198.38 ± 44.10 |
| Tyramine | 20.22 ± 0.04 | 2706.41 ± 11.51 | 1247.27 ±8.55 | 35.06 ± 21.96 | 31.60 ± 10.39 | 7.28 ± 2.16 | 35.25 ± 22.54 | 187.07 ± 58.78 |
| 2-Phenyl-etlhylamine | 5.58 ± 6.03 | 46.06 ± 0.00 | 21.24 ± 0.77 | 10.35 ± 1.11 | 3.47 ± 0.13 | 76.19 ± 16.79 | 573.32 ± 185.2 | 3.47 ± 0.13 |
| Chemical Parameter | SRM Treatment | |||
|---|---|---|---|---|
| A1 (ST1 + M78) | A2 (ST1 + M78 + KE82) | A3 (ST1 + M78 + GL31) | A4 (ST1 + M78 + H25) | |
| Milk pH 2 | 4.25 ± 0.04 ab | 4.28 ± 0.05 b | 4.29 ± 0.04 b | 4.18 ± 0.11 a |
| Lactose | 2.88 ± 0.13 ab | 3.00 ± 0.12 b | 2.78 ± 0.17 ab | 2.67 ± 0.30 a |
| D-glucose | Traces | 0.02 ± 0.02 a | 0.02 ± 0.01 a | 0.04 ± 0.02 a |
| D-galactose | 0.98 ± 0.11 b | 0.90 ± 0.10 a | 0.84 ± 0.11 a | 0.94 ± 0.09 ab |
| L-lactate | 640.3 ± 167.2 b | 734.2 ± 23.6 b | 747.6 ± 26.3 b | 402.0 ± 147.2 a |
| Acetate | 13.6 ± 3.5 a | 22.8 ± 3.5 b | 33.0 ± 2.2 c | 20.0 ± 0.4 b |
| Parameter | Artisan Galotyri Cheese Treatment | |||||
|---|---|---|---|---|---|---|
| A1 (ST1 + M78) | A2 (ST1 + M78 + KE82) | A4 (ST1 + M78 + H25) | ||||
| Fresh Cheese (Day 0) | Ripened Cheese (Day 30) | Fresh Cheese (Day 0) | Ripened Cheese (Day 30) | Fresh Cheese (Day 0) | Ripened Cheese (Day 30) | |
| pH | 4.52 ± 0.20 a | 4.50 ± 0.17 a | 4.60 ± 0.33 a | 4.50 ± 0.24 a | 4.47 ± 0.14 a | 4.47 ± 0.19 a |
| Moisture (%) | 67.5 ± 6.6 a | 63.5 ± 7.4 a | 68.3 ± 4.5 a | 64.7 ± 7.2 a | 68.3 ± 4.8 a | 65.9 ± 4.4 a |
| Fat (%) | 13.9 ± 2.5 b | 13.8 ± 1.8 b | 13.5 ± 1.8 b | 13.2 ± 1.6 ab | 13.0 ± 1.4 ab | 12.4 ± 0.6 a |
| Protein (%) | 12.9 ± 0.9 c | 11.8 ± 0.4 bc | 10.8 ± 0.3 a | 11.4 ± 0.4 ab | 10.9 ± 0.9 a | 10.6 ± 1.5 a |
| Salt (NaCl) (%) | 1.73 ± 1.13 a | 2.31 ± 1.30 a | 1.56 ± 0.64 a | 2.07 ± 0.64 a | 1.58 ± 0.69 a | 1.87 ± 0.68 a |
| Ash (%) | 2.50 ± 1.11 a | 2.83 ± 1.09 a | 2.26 ± 0.65 a | 2.57 ± 0.43 a | 2.21 ± 0.71 a | 2.50 ± 0.64 a |
| Acidity (%) | 1.07 ± 0.20 a | 1.04 ± 0.03 a | 0.99 ± 0.20 a | 1.14 ± 0.31 a | 0.96 ± 0.07 a | 1.06 ± 0.13 a |
| Gram-Negative Bacterial Group | Cheese Trial | Artisan Galotyri Cheese Treatment | |||||
|---|---|---|---|---|---|---|---|
| A1 (ST1 + M78) | A2 (ST1 + M78 + KE82) | A4 (ST1 + M78 + H25) | |||||
| Fresh Cheese (Day 0) | Ripened Cheese (Day 30) | Fresh Cheese (Day 0) | Ripened Cheese (Day 30) | Fresh Cheese (Day 0) | Ripened Cheese (Day 30) | ||
| Pseudomonad-like bacteria (CFC agar) | GL1 | <3.00 (5.26 ± 0.11) * | <3.00 (5.72 ± 0.34) * | <3.00 (5.31 ± 0.01) * | <3.00 (5.96 ± 0.25) * | <3.00 (6.06 ± 0.07) * | <3.00 (6.12 ± 0.06) * |
| GL2 | 7.05 ± 0.01 | <3.00 (5.54 ± 0.07) * | 5.22 ± 0.02 | <3.00 (5.78 ± 0.19) * | 3.63 ± 0.31 | <3.00 (5.13 ± 0.00) * | |
| GL3 | <2.00 | <2.00 | <2.00 | <2.00 | <2.00 | <2.00 | |
| Coliform bacteria (VRB agar) | GL1 | 6.26 ± 0.09 | <1.00 | 5.40 ± 0.13 | <1.00 | 6.11 ± 0.01 | <1.00 |
| GL2 | 6.25 ± 0.07 | 4.43 ± 0.07 | 4.83 ± 0.18 | 2.13 ± 0.39 | 3.96 ± 0.15 | <1.00 | |
| GL3 | <1.00 | <1.00 | <1.00 | <1.00 | <1.00 | <1.00 | |
| Biogenic Amine | Cheese Trial | Artisan Galotyri Cheese Treatment | |||||
|---|---|---|---|---|---|---|---|
| A1 (ST1 + M78) | A2 (ST1 + M78 + KE82) | A4 (ST1 + M78 + H25) | |||||
| Fresh Cheese | Ripened Cheese | Fresh Cheese | Ripened Cheese | Fresh Cheese | Ripened Cheese | ||
| Cadaverine | GL1 | 18.1 | NT | 4.7 | NT | 21.9 | NT |
| GL2 | 158.6 | 3521.1 | <0.5 | 5664.8 | 815.1 | 3350.4 | |
| GL3 | 242.6 | 5.8 | 4.0 | 4.1 | 4.0 | 4.4 | |
| Histamine | GL1 | 245.9 | NT | <0.5 | NT | <0.5 | NT |
| GL2 | 503.5 | 87.5 | <0.5 | 373.1 | 179.8 | 145.9 | |
| GL3 | 130.2 | 15.3 | 14.9 | 16.6 | 14.0 | 16.4 | |
| Putrescine | GL1 | <0.5 | NT | <0.5 | NT | <0.5 | NT |
| GL2 | <0.5 | 897.3 | <0.5 | 1396.7 | 373.3 | 278.4 | |
| GL3 | 570.9 | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | |
| Spermidine | GL1 | <0.5 | NT | <0.5 | NT | <0.5 | NT |
| GL2 | 74.4 | <0.5 | <0.5 | 34.3 | 21.6 | 13.1 | |
| GL3 | 60.4 | 9.4 | 9.2 | 9.3 | 9.2 | 9.2 | |
| Tryptamine | GL1 | 10.9 | NT | 1.1 | NT | 0.8 | NT |
| GL2 | <0.5 | 401.3 | <0.5 | <0.5 | 99.5 | <0.5 | |
| GL3 | 79.9 | 6.0 | 5.0 | 7.0 | 4.6 | 6.9 | |
| Tyramine | GL1 | 26.3 | NT | 21.8 | NT | 4.4 | NT |
| GL2 | 591.0 | 90.0 | 3.7 | 134.1 | 165.2 | 61.8 | |
| GL3 | 103.3 | 3.9 | 7.0 | 3.9 | 4.6 | 4.0 | |
| 2-Phenylethyl Amine | GL1 | <0.5 | NT | <0.5 | NT | <0.5 | NT |
| GL2 | 251.6 | 140.1 | <0.5 | 125.9 | 1989.5 | 34.1 | |
| GL3 | 65.5 | 1.3 | 1.5 | 2.7 | 1.4 | <0.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsanasidou, C.; Bosnea, L.; Kakouri, A.; Samelis, J. Biogenic Amine Formation in Artisan Galotyri PDO Acid-Curd Cheeses Fermented with Greek Indigenous Starter and Adjunct Lactic Acid Bacteria Strain Combinations: Effects of Cold (4 °C) Ripening and Biotic Factors Compromising Cheese Safety. Appl. Microbiol. 2024, 4, 536-562. https://doi.org/10.3390/applmicrobiol4010038
Tsanasidou C, Bosnea L, Kakouri A, Samelis J. Biogenic Amine Formation in Artisan Galotyri PDO Acid-Curd Cheeses Fermented with Greek Indigenous Starter and Adjunct Lactic Acid Bacteria Strain Combinations: Effects of Cold (4 °C) Ripening and Biotic Factors Compromising Cheese Safety. Applied Microbiology. 2024; 4(1):536-562. https://doi.org/10.3390/applmicrobiol4010038
Chicago/Turabian StyleTsanasidou, Charikleia, Loulouda Bosnea, Athanasia Kakouri, and John Samelis. 2024. "Biogenic Amine Formation in Artisan Galotyri PDO Acid-Curd Cheeses Fermented with Greek Indigenous Starter and Adjunct Lactic Acid Bacteria Strain Combinations: Effects of Cold (4 °C) Ripening and Biotic Factors Compromising Cheese Safety" Applied Microbiology 4, no. 1: 536-562. https://doi.org/10.3390/applmicrobiol4010038