Consumer awareness of the link between wholesomeness and balanced nutritional food is increasing, as well as the need for a product with the right taste and quality profile to meet their needs. Flavonoids are valuable feed additives to improve gut health or meat quality in poultry. Rutin, one of the cheapest flavonoids, has a variety of biological functions, such as antioxidant, antidiabetes, and antidyslipidemia activity [
10,
21,
22], and may have regulatory effects on lipid metabolism, fatty acid synthesis, and meat quality of Chinese indigenous, yellow-feathered broiler. In this study, rutin supplementation dramatically increased the breast muscle yield, IMF, HDL,
n-3 PUFAs, and antioxidant capacity but decreased the drip loss, TG levels, TC levels, and the proportion of
n-6/
n-3 PUFAs of Qingyuan partridge chickens. Additionally, these alterations were linked to the upregulated expression of lipid metabolism-related and antioxidant-related genes. These findings highlight the improvement of meat quality, antioxidant capacity, and especially, the FA profile in Qingyuan partridge chickens, suggesting that rutin supplementation contributes to the increased nutritional value and taste of the meat of Qingyuan partridge chickens.
4.2. Dietary Rutin Supplementation Altered the Meat Quality of Qingyuan Partridge Chickens
Drip loss is a vital indicator for judging meat quality, which is tightly correlated with the physical structure, flavor, juiciness, and color of meat. Unfavorably high drip loss would not only induce the loss of soluble flavor substance in meat but would also have a detrimental impact on the sensory quality of the meat end-product and consumer acceptance [
24]. Our results revealed that drip loss in breast muscle was markedly reduced after the dietary inclusion of rutin. Likewise, dietary supplementation with alfalfa flavonoids was reported to have a significant decrease in breast-muscle drip loss of broilers [
6]. Consistent with the results above, dietary supplementation with hesperidin reduced breast-muscle drip loss of broilers [
25]. Lipid peroxidation can change the membrane structure and functions of muscle cells and impair their water-holding capacity [
26]. Moreover, secondary products of lipid oxidation could accelerate the oxidation of proteins and change their stability or peptide structure, resulting in substantial loss of biological function and water-holding capacity [
27,
28]. Thus, the increased antioxidant capacity in broilers by supplementation with flavonoids probably accounts for the decreased drip loss [
6,
25].
The IMF content is also thought to be a critical factor in determining the nutritional value, tenderness, flavor, and juiciness of the meat. Our findings also indicated that dietary rutin supplementation significantly increased IMF content in breast muscle. Similarly, dietary supplementation with flavones of sea buckthorn fruits was reported to significantly increase the IMF level in breast and thigh muscles, presumably as a result of the increased levels of insulin and leptin in serum [
29]. However, dietary supplementation with soybean isoflavone was not found to alter the IMF concentration of breast muscle in broilers [
7], and hesperidin supplementation resulted in no significant change in the IMF concentration of the
Longissimus thoracis muscle of lambs [
30]. These inconsistencies may be relevant to the differences in experimental animals, the kinds of flavonoids used, and the duration of the experiment. The IMF could retain higher water content by altering the microstructure of meat, which facilitates the juiciness and tenderness of meat [
31]. Our results have provided potential evidence that rutin supplementation significantly increased IMF while simultaneously reducing drip loss. In addition, IMF also contributes to the flavor profile of meat due to its high content of UFAs, which act as crucial precursors for the formation of desirable flavor compounds in meat [
32]. Accordingly, the augmented content of IMF, which, in turn, could potentially enhance the meat’s sensory attributes, such as tenderness, juiciness, and flavor, may account for the improved meat quality by dietary rutin supplementation. However, further investigations are needed to elucidate the mechanisms involved in the regulation of the IMF content by rutin supplementation.
To the best of our knowledge, the present study, for the first time, investigates the influence of rutin supplementation on the meat quality of chickens. Our results suggested that the addition of rutin to the diet improved the nutritional value, juiciness, and water retention ability of the meat of Qingyuan partridge chickens.
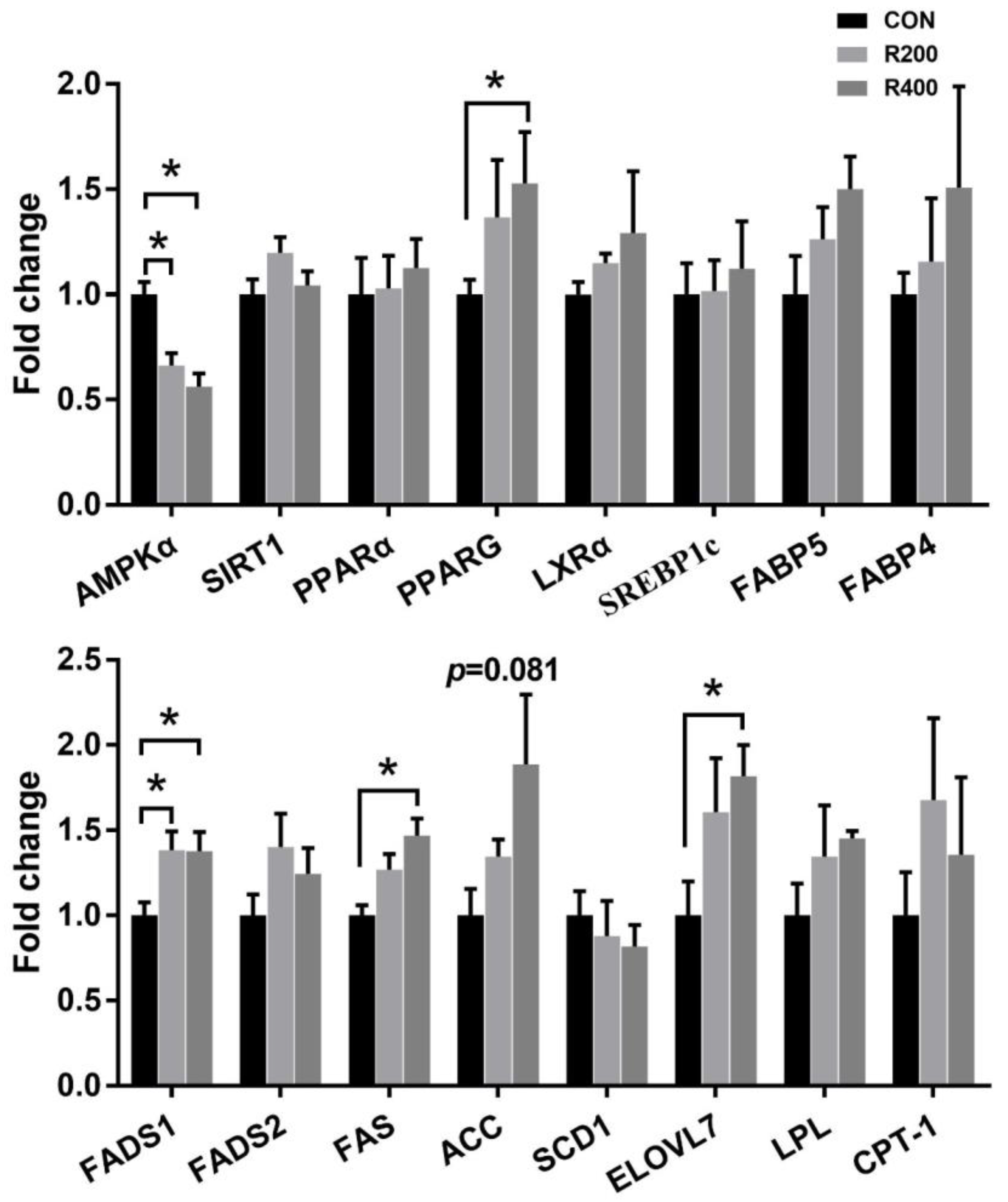
4.3. Dietary Rutin Supplementation Altered the Fatty Acid Profile and Lipid Metabolism of Qingyuan Partridge Chickens
The FA profile of IMF is one of the most critical indexes closely correlated with the flavor and nutritional value of the meat [
33]. Meat with a healthier FA profile is of considerable interest to the meat industry and consumers. PUFAs, particularly
n-3 PUFAs, including α-linolenic acid (ALA, C18:3
n − 3) and DHA, exert a central role in promoting the formation of meat flavor substances and maintaining human health [
34]. However, many organisms, including humans, lack the enzymes to synthesize ALA de novo, which is a crucial precursor for DHA; therefore, these organisms must obtain ALA from the diet [
35]. DHA is mainly obtained through dietary sources or biosynthesis from ALA. The conversion of ALA to DHA involves a series of complex elongation and desaturation processes, in which Δ5 and Δ6 desaturases encoded by FA desaturase 1 and 2 (
FADS1 and
FADS2) genes are the rate-limiting enzymes [
36]. It has been shown that the administration of flavonoid supplements (anthocyanin extracted from corn) increased the concentrations of DHA and
n-3 PUFAs in the plasma of rats [
37]. However, Gallegos et al. [
38] reported that dietary supplementation with anthocyanin extracted from purple corn did not affect hepatic EPA and DHA concentrations in rats. In chicken, the concentrations of DHA and
n-3 PUFAs in the hen liver were considerably increased by genistein supplementation [
39]. Furthermore, a linseed oil (ALA-rich)-enriched diet supplemented with soybean isoflavone significantly increased the EPA and DHA contents in breast muscle in comparison to a linseed oil diet alone, accompanied with higher
FADS2 gene expression [
7]. In accordance with the above two reports, we also found that rutin dietary inclusion increased the content of DHA and
n-3 PUFAs in breast muscle, thereby improving fatty acid composition in chickens. In addition, higher
FADS1 mRNA levels and the product-to-precursor ratio of Δ5 and Δ6 activity were observed in chickens with rutin supplementation. Additionally, flavonoids, including quercetin, daidzein, and genistein, could increase the mRNA levels of Δ5 or Δ6 desaturases in HepG2 cells [
40]. Thus, the increased level of
n − 3 PUFAs in the present study may result from the increased activity and mRNA expression of Δ5 and Δ6 desaturases. Noteworthy, to date, the research on chickens seems more consistent than that on other organisms, including rodents and humans. On the other hand, PUFA-rich chickens contain a greater proportion of double-bonded fatty acids, which are vulnerable to oxidative attack [
41]. The increased antioxidant capacity could provide oxidative-proof protection for PUFAs and possibly contribute to the accumulation of PUFAs. However, further investigations are needed to establish the underlying mechanisms.
The content of lipid metabolism-related products can be used as an indicator to assess the lipid metabolism function of the body and is tightly linked with the growth and development of animals and their health states [
42]. The TG and TC are important fractions of blood fat, and their contents reflect the fat utilization rate. LDL enriches cholesterol, and a high content of LDL can cause atherosclerosis. HDL could carry cholesterol from peripheral tissue to the liver to synthesize bile acid, which contributes to reducing the formation of atherosclerosis [
43]. It has been reported that oral rutin could decrease serum TC and TG contents and increase the serum HDL content in rats [
44]. Furthermore, dietary rutin supplementation was found to decrease the contents of TC, TG, and LDL in the serum of broilers [
8]. These studies are in accordance with our finding that dietary rutin supplementation significantly decreased the serum contents of TC and TG but increased the serum HDL content. Flavonoids could be reciprocal with cholesterol and suppress the intestinal absorption of endogenous and exogenous cholesterol by forming insoluble complexes [
45]. In parallel, rutin reduced TG synthesis through the reduction in lipid synthesis and acceleration of FA catabolism in the liver [
46]. Thus, a possible explanation for the improvement of the homeostasis of blood lipids is that rutin could inhibit the synthesis of lipid and intestinal absorption of cholesterol and accelerate the lipid decomposition of cells in the liver and the reverse transport of cholesterol.
TGs, as the primary constituent of IMF, are crucial for the manipulation of energy metabolism in muscle. The metabolic processes involved in energy utilization and lipid storage in muscular tissue are inseparable from the biosynthesis and deposition of TGs [
47]. As a key molecule in maintaining energy balance, AMPK regulates lipid accumulation and gene transcription [
48]. AMPK increases FA oxidation, reduces FA synthesis, and improves lipid homeostasis through activation of Carnitine palmitoyl transterase-1 (CPTl) and PPARα and suppression of ACC in the liver [
49]. It has been reported that the IMF of the
biceps femoris muscle in dry-lot feeding lambs was increased by downregulating the expression level of AMPK and upregulating the expression level of ACC compared to those in grazing lambs [
50]. Consistent, in part, with the above study, our study showed that rutin supplementation downregulated mRNA expression levels of
AMPK and tended to upregulate mRNA expression levels of
ACC in breast muscle, which is partially responsible for the increased lipid deposition. PPARs are AMPK downstream targets, and AMPK could regulate body fat deposition by decreasing mRNA expression of
PPARG [
51]. PPARG is an essential manipulator that regulates adipogenesis and glucose homeostasis. In mice, rutin was reported to upregulate the expression level of
PPARG in skeletal muscles [
52]. In this study, rutin supplementation upregulated the expression level of
PPARG in breast muscle. Thus, the AMPK/PPARG signaling pathway may regulate the IMF content in breast muscle. FAS represents a critical regulatory component of the biosynthetic pathway of FA, primarily mediating the ultimate step of FA synthesis in muscular tissue [
53]. In contrast to the previous studies [
51,
54], indicating that rutin inhibited the expression and transcription of
FAS in hepatocytes, our results indicated that the mRNA expression of
FAS was upregulated by rutin supplementation. These controversial results might be due to the different species of animals and tissues used for the measurement. Long-chain fatty acids (LCFAs) and very long-chain fatty acids (VLCFAs) are important participants in the synthesis of TGs and lipid metabolism. The elongation of the very long-chain fatty acid (ELOVL) protein family is necessary for the synthesis of LCFAs and VLCFAs [
55]. In particular,
ELOVL7, a newly discovered ELOVL protein family member, evokes lipid accumulation in differentiated adipocytes [
56].
ELOVL7 was found to be one of the overexpressed key genes associated with adipogenesis and lipogenesis in the breast muscle of animals in the group with higher TG content [
47]. Therefore, the increased mRNA expression of
ELOVL7 is potentially responsible for the rutin treatment-induced increased IMF in breast muscle. Overall, these findings indicate that rutin supplementation changed the FA profile and lipid metabolism in Qingyuan partridge chickens by manipulating the lipid metabolism-related gene expression.
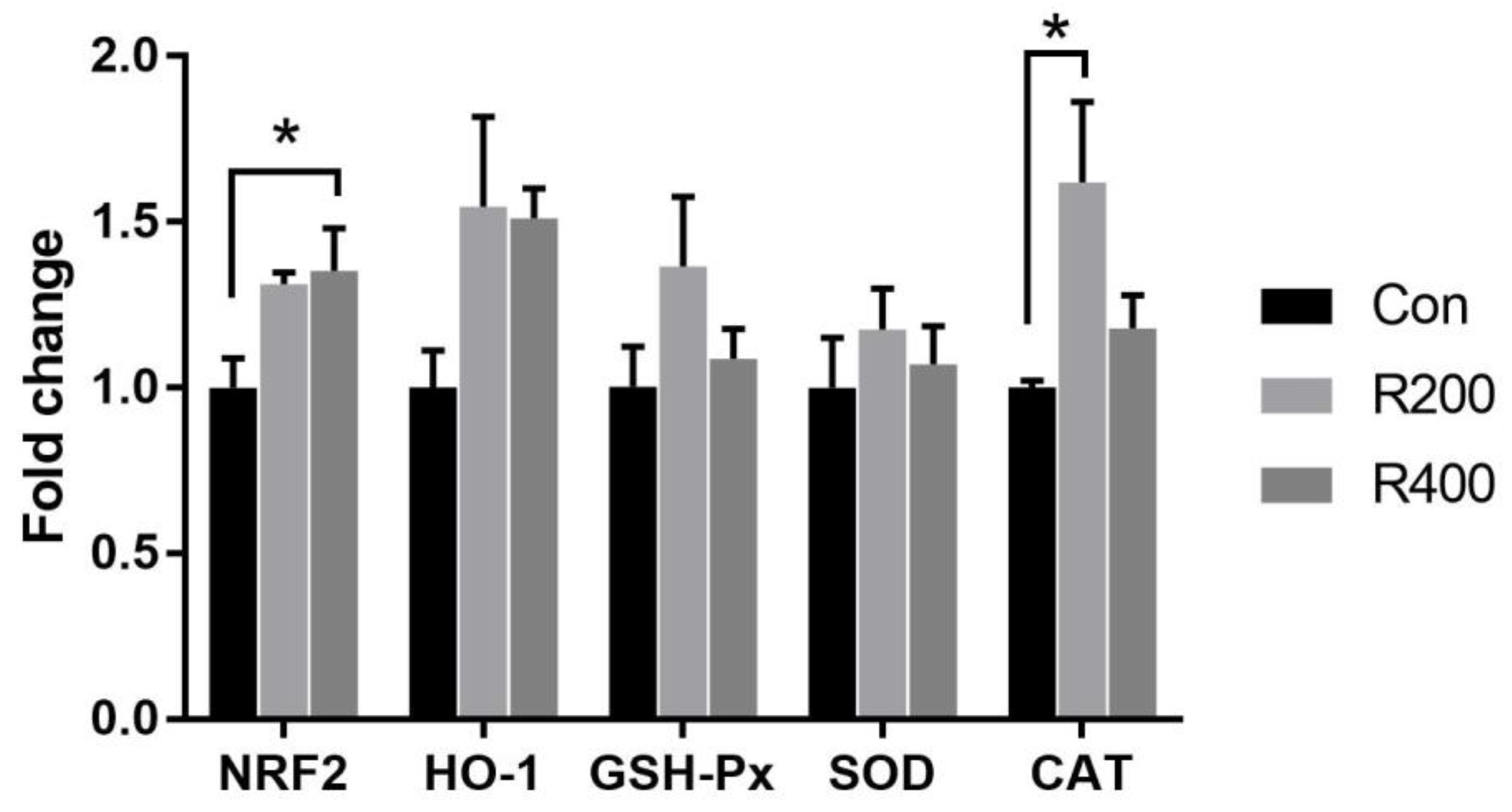
4.4. Dietary Rutin Supplementation Altered the Antioxidant Capacity of Qingyuan Partridge Chickens
The antioxidant capacity is positively linked with the body’s health and meat quality [
57]. The MDA content is considered an index for the degree of lipid peroxidation [
58]. SOD, GSH-Px, and CAT are the main antioxidant enzymes in the organism, which can efficiently scavenge free radicals. Furthermore, T-AOC broadly reflects the antioxidant capacity of living organisms [
59]. Hassan et al. [
8] showed that the activities of anti-oxidase (SOD, GSH-Px, and CAT) and the MDA content in the serum of broilers were increased and decreased by rutin supplementation, respectively. Similarly, dietary rutin supplementation has been found to increase the T-AOC and T-SOD activities in the jejunal mucosa of broilers [
9]. Our findings also indicated that rutin supplementation prominently elevated the T-AOC and CAT activities but decreased the MDA content in serum and breast muscle. The activities of antioxidant enzymes partly depend on their gene expression levels, which have been shown to be regulated by
NRF2 [
60]. In this study, we further revealed that rutin supplementation increased the mRNA levels of
NRF2 and
CAT in breast muscle. Rutin, which is regarded as a powerful antioxidant, effectively inhibited lipid peroxidation and directly scavenged the generated ROS, such as hydroxyl radicals, H
2O
2, and superoxide anions [
61]. Rutin also promoted the production of GSH-Px and SOD by activating the
NRF2 signaling pathway, which led to the improvement of the scavenging of free radicals and maintenance of the body’s redox balance [
62]. Therefore, dietary rutin supplementation can enhance the antioxidant capacity and alleviate oxidative stress, ultimately having a positive effect on the health and meat quality of chickens.

 ,
, 




{kind=link}
{kind=link}
{kind=link}